Класс Паукообразные, или арахниды (Arachnida)
Паукообразные, или арахниды (Arachnida)*, - это собрание всех наземных хелицеровых. Их насчитывается около 35 000 видов, и они очень различны по внешнему облику. Различают от 9 до 13 отрядов современных арахнид и несколько ископаемых. Среди них семь отрядов общеприняты: скорпионы (Scorpiones), печений (Palpigradi), сольпуги (Solifugae), ложноскорпионы (Pseudoscorpiones), сенокосцы (Opiliones), рицинулеи (Ricinulei) и пауки (Aranei). Но в понимании нескольких группировок имеются разноречия. Это телифоны (Uropygi), фрины (Amblypygi) и тартариды (Tartarides), объединяемые в группу жгутоногих (Pedipalpi), и клещи (Acarina), на классификации которых мы остановимся в дальнейшем. При большом разнообразии арахнид основные признаки хелицеровых свойственны им всем. Тело состоит из головогруди - просомы и брюшка - опистосомы, соединенных в области седьмого, предполового, сегмента. Усиков нет, глаза простые. Конечности головогруди - хелицеры, педипальпы и 4 пары ног - служат для захвата пищи и передвижения; конечности брюшка видоизменены, выполняют дыхательную и другие специальные функции и в значительной части атрофируются. Отличия арахнид от первичноводных хелицеровых обусловлены приспособлениями к жизни на суше. Главные из них: превращение жаберных ножек в легкие и далее замена их дыхательными трубками - трахеями; дальнейшая концентрация отделов тела; приспособление ног для передвижения по суше, а околоротовых конечностей для питания полужидкой пищей - содержимым жертвы, предварительно растворенным пищеварительными соками; ряд изменений жизненного цикла и общее уменьшение размеров.
* (Латинское название класса, в этой транскрипции теперь более принятое, прежде писалось Arachnoidea.
Арахна - по-гречески "паук". В древнегреческих мифах это имя девушки, которая, по преданию, достигла такого высокого ткацкого искусства, что вызвала на состязание саму богиню Афину. Арахна выткала ткань не хуже Афины, но та в наказание за дерзость состязаться с богами не признала ее достоинств. В отчаянии Арахна хотела повеситься, тогда Афина превратила ее в паука, вечно ткущего свою паутину.)

Таблица 2. Скорпион пестрый (Buthus eupeus) и сольпуга обыкновенная (Galeodes araneoides)
Строение головогруди (просомы) в общем однотипно. Обычно все 6 сегментов просомы слиты и она покрыта цельным головогрудным щитом. Но у сольпуг, кенений и части клещей слиты только четыре передних сегмента, отвечающие сегментам головы трилобитов. Они покрыты головным щитом (пропельтидием), а сегменты третьей и четвертой пар ног расчленены и имеют свои тергиты, - состояние, более примитивное, чем даже у меростомовых. Строение и функции околоротовых конечностей связаны со способом питания. Подавляющее большинство арахнид хищники, питающиеся живой добычей, главным образом насекомыми. При этом покровы жертвы разрываются и внутрь вводятся пищеварительные соки, которые обладают протеолитическим действием (способностью растворять белки). Затем разжиженное содержимое жертвы всасывается. Питание полужидкой пищей привело к тому, что у арахнид околоротовые конечности не приобрели характера челюстей в том виде, как у насекомых. Хелицеры служат для схватывания и разрывания добычи. Они обычно короткие, клешневидные; иногда концевой членик хелицер имеет вид когтя, на конце которого открывается проток ядовитой железы (например, у пауков), или хелицеры колющие,игловидные (у многих клещей). Тазики педипальп имеют отростки - эндиты, но они обычно не служат для пережевывания пищи, а ограничивают предротовую полость, на дне которой расположено ротовое отверстие (рис. 11). Верхняя стенка этой полости образована эпистомом с верхней губой. Изнутри на эндитах педипальп и в глотке имеются волоски, через которые фильтруется полужидкая пища. После питания твердые частички счищаются с волосков и выбрасываются. Щупальца педипальп служат органами осязания, но иногда они участвуют в передвижении (сольпуги, кенении), или они хватательные, с клешнями (скорпионы, ложноскорпионы) или когтевидными выростами (жгутоногие). В строении ног характерно образование членистой лапки с коготками - приспособление к хождению по суше. Жевательная функция ног у арахнид утрачена, но коксэндиты у примитивных форм частично сохраняются. Ноги, особенно передние, богато оснащены осязательными волосками и наряду со щупальцами педипальп имитируют исчезнувшие усики.
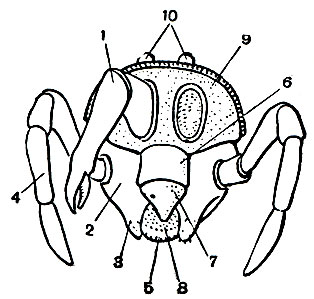
Рис. 11. Околоротовые придатки арахнид (вид спереди): 1 - хелицеры (левая не показана); 2 - тазики педипальп; 3 - их жевательные отростки; 4 - щупальца педипальп; 5 - стернит педипальп; 6 - эпистом; 7 - верхняя губа; 8 - предротовая полость; 9 - головогрудной щит; 10 - медиальные глаза
Наибольшее разнообразие наблюдается в строении брюшка (рис. 12). Число сегментов его заднего отдела - метасомы - постепенно сокращается, и брюшко укорачивается. Только у скорпионов метасома ("хвост") состоит, как у эвриптерид, из 6 сегментов и концевого членика и общее число сегментов тела равно 19. У жгутоногих и пауков в метасоме 5 сегментов, у сольпуг, кенений, сенокосцев и рицинулей - 4, у ложноскорпионов - 3, у примитивных клещей - 2-1, у некоторых паразитических клещей исчезает не только метасома, но и несколько сегментов мезосомы. У некоторых отрядов оставшиеся сегменты метасомы имеют вид маленького стебелька (постабдомена), иногда с членистой хвостовой нитью. Степень развития соединительного предполового сегмента различна. У скорпионов, сенокосцев и рицинулей он не выражен, у ложноскорпионов и части клещей он, напротив, развит, как соседние. В обоих случаях брюшко широким основанием примыкает к головогруди. У других арахнид этот сегмент меньше остальных и иногда представлен узким стебельком, подвижно соединяющим головогрудь и брюшко. На этом основании арахнид иногда явно искусственно делят на сидячебрюхих и стебельчатобрюхих. Степень слияния сегментов брюшка различна. Обычно их границы хорошо различимы, но у большинства пауков и клещей границы сегментов исчезают. У многих клещей и головогрудь с брюшком сливаются, в одно целое.
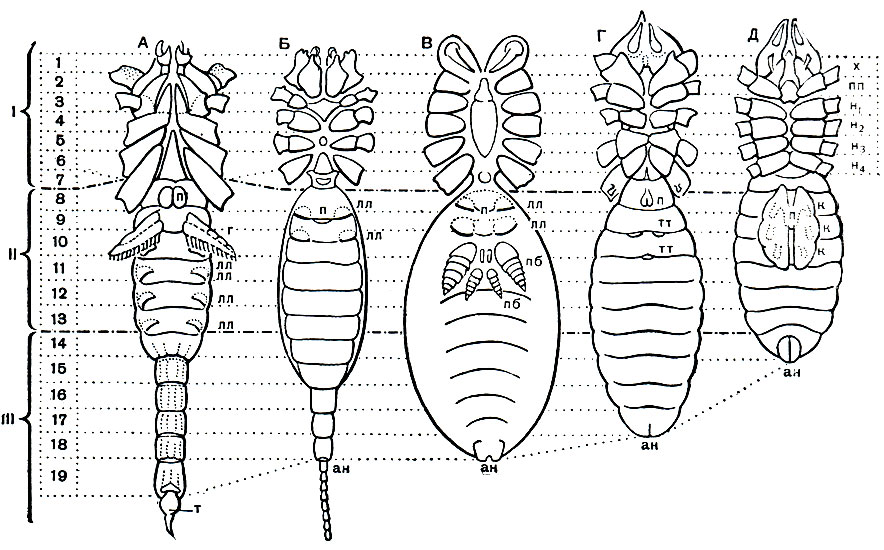
Рис. 12. Строение арахнид (вид с брюшной стороны): А - скорпиона; Б - телифона; В - паука; Г - сольпуги; Д - примитивного акариформного клеща; I - просома; II - мезосома; III - метасома; 1-19 - сегменты тела; m - хвостовой придаток; лл - стигмы легких; mm - стигмы трахей; n - половое отверстие; г - гребневидные органы; nб - паутинные бородавки; к - коксальные органы; ан - анальное отверстие; х - хелицеры; пп - педипальпы; н1 - н4 - ноги
Конечности брюшка у арахнид превращены в легкие и другие специальные образования. Они имеются только на сегментах мезосомы. Наиболее полный набор видоизмененных брюшных конечностей сохранился у скорпионов: половые крышечки на восьмом сегменте, гребневидные органы на девятом, четыре пары легких на десятом - тринадцатом сегментах. У телифонов, фринов и четырехлегочных пауков по паре легких имеется на восьмом и девятом сегментах, у тартарид и двулегочных пауков - пара легких на восьмом сегменте, причем у последних на месте легких на девятом сегменте образуются трахеи. У всех пауков конечности десятого и одиннадцатого сегментов превращены в паутинные бородавки. У других арахнид легкие исчезают. Иногда на их месте открываются трахеи (сольпуги, сенокосцы), в других случаях трахеи не имеют отношения к легким. Рудиментами конечностей брюшка являются еще так называемые коксальные органы, имеющиеся на восьмом - десятом сегментах у кенений и части клещей, у которых на брюшке нет органов дыхания. Они имеют вид маленьких выпячивающихся мешочков, наполняемых гемолимфой, и, по-видимому, служат органами чувств, определяющими влажность (гигрорецепторы). Они приурочены к тазикам ножек и при утрате последних остаются на их месте. У кенений они расположены на брюшке открыто, а у части клещей входят в состав сложного наружного полового аппарата, указывая на участие в его образовании трех пар видоизмененных конечностей восьмого - десятого сегментов. Заметим, что система подобных коксальных органов наиболее полно развита у некоторых многоножек и низших насекомых. Наличие коксальных органов на брюшке кенений и низших клещей указывает на то, что у этих мелких форм легких никогда не было.
Покровы арахнид образованы хитинизированной кутикулой со слоем подстилающих плоских клеток - гиподермой. Щитки и членики конечностей образованы твердой кутикулой, в сочленениях между ними кутикула более тонкая, эластичная. Наиболее тверда кутикула коготков, пальцев хелицер. У некоторых скорпионов в кутикуле имеются отложения углекислого кальция. Строение наружного скелета арахнид различно и в той или иной мере отражает степень слияния сегментов. Головогрудь защищена сверху твердым щитом, между тазиками ног иногда имеется грудная пластинка (стернум). Тергиты и стерниты брюшка обычно посегментно расчленены. Но у части сенокосцев и многих клещей тергиты срастаются друг с другом и с головогрудным щитом, образуя цельный спинной щит, а иногда к ним примыкает и часть стернитов брюшка, так что тело заковано в панцирь. В других случаях, например у пауков, щиты имеются на головогруди, а брюшко покрыто эластичной кутикулой. Среди низших почвенных и эндопаразитических клещей есть формы с очень тонким покровом, лишенные щитков. Установлено, что кутикула арахнид имеет особые оптические свойства, она обладает двойным лучепреломлением, т. е. поляризует проходящий через нее свет. Благодаря этому в поляризационном микроскопе определенные элементы покрова светятся в темном поле. Французский исследователь Ф. Гранжан, впервые обнаруживший это явление у клещей, приписывает его наличию в кутикуле хитина особой структуры, названного им актинохитином или лучистым хитином. Распределение актинохитина в скелете закономерно. У одних отрядов он имеется только в толще щитков, у других только в волосках и щетинках, у третьих и там и тут.

Рис. 13. Свечение актинохитина щетинок у акариформного клеща в поляризованном свете
Будучи хищниками, арахниды принуждены справляться подчас с сильной добычей. Мускулатура хорошо развита, особенно мышцы головогруди, приводящие в движение конечности. Разнообразны железы покровного (гиподермального) происхождения: железы предротовой полости пауков, лобные и анальные железы жгутоногих, пахучие железы сенокосцев и др. К этой же категории относятся ядовитые и паутинные железы. Первые имеются у скорпионов в концевом членике брюшка, у пауков, у которых открываются на крючках хелицер, у ложноскорпионов, некоторых клещей. Ядовитый аппарат скорпионов и пауков служит весьма эффективным средством нападения и защиты. Паутинные железы имеются у ложноскорпионов, некоторых клещей и пауков. У последних они особенно развиты и открываются многочисленными отверстиями на брюшных паутинных бородавках.
Органы чувств образуются путем дифференциации клеток покровного эпителия. Глаза имеются на просоме в разном числе: до 5 пар у скорпионов, обычно 4 пары у пауков ижгутоногих, 2-1 пара у большинства других; кенении, многие клещи, рицинулеи слепы. Глаза построены по типу простых глазков (оцелл). В глазу имеется диоптрический аппарат - хрусталик, образованный прозрачным утолщением кутикулы, и стекловидное тело, а под ним слой чувствительных клеток (ретина), связанных волокнами зрительного нерва с мозгом (рис. 14). Пара срединных (главных) глаз и боковые различаются в деталях строения. Зрительные возможности большинства арахнид ограниченны, они воспринимают вариации освещенности и движение. Лучше других видят сольпуги и бродячие пауки. Среди последних пауки-скакуны обладают предметным зрением, но различают форму на сравнительно близком расстоянии.
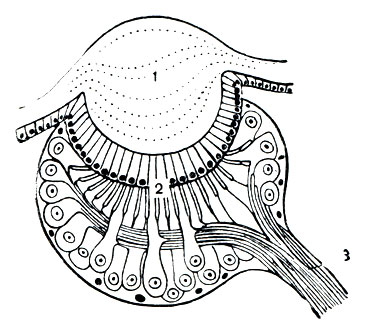
Рис. 14. Строение медиального глаза арахнид: 1 - светопреломляющий аппарат; 2 - сетчатка; 3 - зрительный нерв
Слабое зрение компенсируется осязанием, играющим первостепенную роль в поведении арахнид. На теле и конечностях имеются многочисленные осязательные волоски, к основаниям которых подходят нервные окончания чувствительных клеток. По величине и форме эти волоски у арахнид чрезвычайно разнообразны. Кроме того, имеются специальные волоски, воспринимающие колебания,- трихоботрии. Эти своеобразные органы обычно в определенном количестве имеются на педипальпах и ногах, иногда на туловище (у части клещей). Длинный торчащий волосок, иногда утолщенный на конце, прикрепляется тонкой мембраной на дне воронковидного углубления. Малейшее сотрясение или дуновение воздуха приводит его в колебания, которые воспринимаются группой чувствительных клеток. У арахнид имеются и органы химического чувства, обонятельные и вкусовые. Первыми считают так называемые лировидные органы, многочисленные на туловище и конечностях. Это микроскопические щели в кутикуле, затянутые тонкой мембраной, к которой подходит окончание чувствительной клетки. Лировидным органам, правда, приписываются и другие функции, в частности механорецепторов, воспринимающих степень натяжения кутикулы. Более сложно устроены обонятельные тарзальные органы на лапках передних ног. Чувствительные вкусовые клетки найдены в стенках глотки у пауков.
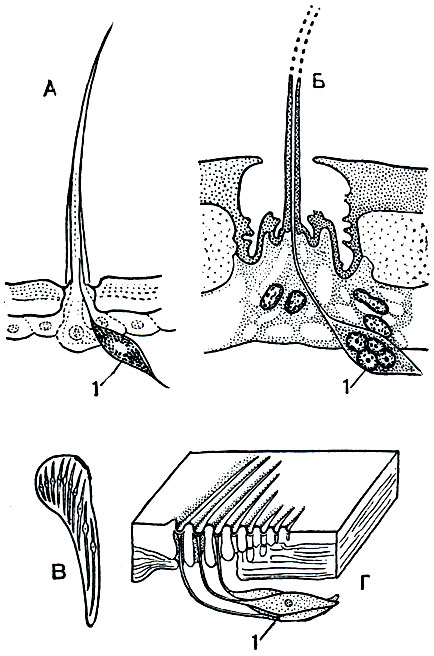
Рис. 15. Покровные органы чувств арахнид: А - осязательный волосок; Б - трихоботрия; В, Г - лировидные органы с поверхности и в разрезе; 1 - чувствительные нервные клетки
Нервная система концентрированна. Отсутствие обособленной головы, усиков и сложных глаз привело к тому, что надглоточный ганглий (головной мозг), иннервирующий у членистоногих эти органы, в той или иной мере объединяется с головогрудной нервной массой. У скорпионов имеется парный надглоточный ганглий, связанный тяжами с подглоточным ганглиозным скоплением, и 7 ганглиев брюшной нервной цепочки. У сольпуг, кроме общей нервной массы, остается один брюшной узел; у большинства арахнид вся нервная цепочка сливается в головогрудную массу.
Кишечник подразделяется на переднюю, среднюю и заднюю кишку. Ротовое отверстие ведет в расширение - снабженную мышцами глотку, которая служит для насасывания полужидкой пищи. Глотка переходит в тонкий пищевод, который у некоторых форм, например пауков, имеет еще расширение - сосательный желудок. Средняя кишка обычно образует несколько пар слепых выростов, увеличивающих ее вместительность и всасывающую поверхность. В брюшке слепые выросты кишечника хорошо развиты и образуют крупный железистый орган-печень. Клетки печени выделяют пищеварительные ферменты, и в них происходит внутриклеточное переваривание пищи. Задний отдел средней кишки образует клоаку, в которой скопляются экскременты и экскрет выделительных мальпигиевых трубок. Отбросы выводятся через короткую заднюю кишку и анальное отверстде. В кишечник арахнид в большинстве случаев попадает только жидкая пища, все крупные частички задерживаются фильтрами предротовой полости и глотки. Будучи прожорливыми хищниками, арахниды способны принимать большое количество пищи и потом долго голодать. Последнее возможно благодаря накоплению питательных веществ в запасной ткани, подобной жировому телу насекомых. Выделительными органами служат коксальные железы и мальпигиевы сосуды. Первые, как говорилось, представляют остатки целомодуктов - посегментно расположенных выделительных органов предков членистоногих - кольчатых червей. Они состоят из экскреторного мешочка, извитого протока (лабиринта) и выводного канала и сохраняются обычно лишь в числе 1-2 пар, открываясь у оснований ног. Мальпигиевы сосуды арахнид - новообразование. Это 1-2 пары слепо замкнутых, иногда ветвящихся трубочек, которые открываются в кишечник близ клоаки. В Клетках их стенок накапливаются экскреты, выводимые затем в клоаку. Выделительную функцию выполняют, кроме того, кишечник, печень, клоака и особые клетки - нефроциты, имеющиеся в полостях между органами. Главный продукт выделения арахнид - гуанин. Это вещество в организме находится в определенных биохимических отношениях с черным пигментом меланином, вместе с ним обусловливая окраску покровов.
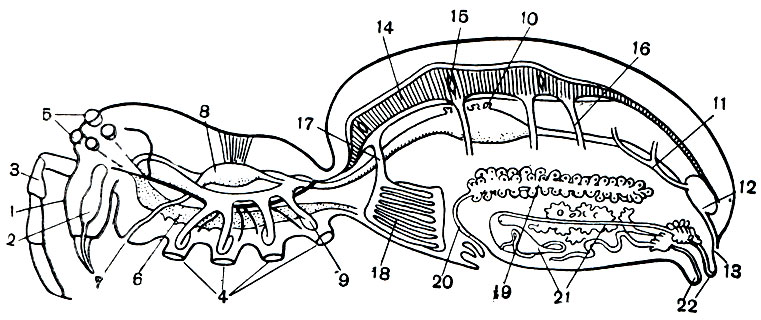
Рис. 16. Схема внутреннего строения паука: 1 - хелицеры; 2 - ядовитая железа; 3 - педипальпы; 4 - ноги; 5 - глаза; 6 - мозг; 7 - ротовое отверстие; 8 - сосательный желудок; 9 - выросты кишечника; 10 - протоки печени (печень не показана); 11 - мальпигиевы сосуды; 12 - клоака; 13 - анальное отверстие; 14 - сердце; 15 - остии; 16 - артерии; 17 - легочная вена; 18 - легкие; 19 - яичник; 20 - яйцевод; 21 - паутинные железы; 22 - паутинные бородавки
Строение дыхательной и кровеносной систем тесно связано. Органы дыхания арахнид двойственны по природе. Это органы локализованного дыхания - легкие, образовавшиеся из брюшных жаберных ножек водных форм, и органы диффузного дыхания - трахеи, возникающие вновь, как более совершенное приспособление для дыхания атмосферным воздухом (рис. 17). Каждый легочный мешок вдается внутрь от щелевидной стигмы. От его внутренней стенки отходят многочисленные листовидные карманы, сложенные подобно страницам книги. В карманах циркулирует кровь, а между ними проникает воздух. Трахеи представляют собой трубки, неветвистые или ветвящиеся, которые доставляют воздух непосредственно к органам и тканям. Их стенки образованы продолжением наружного покрова и выстланы кутикулой, обычно имеющей опорные утолщения: трахеи легко изгибаются, и стенки их не спадаются.
Число пар легких, как говорилось, различно, и в ряде случаев они отсутствуют, заменяясь трахеями, а у некоторых мелких форм нет ни легких, ни трахей, и дыхание кожное (кенении, часть клещей). Число трахейных стволов также различно, причем они могут открываться стигмами в разных местах: на сегментах брюшка, по бокам головогруди, у оснований хелицер, что указывает на независимое их происхождение у разных арахнид. В некоторых случаях трахеи занимают место легких (у сольпуг, двулегочных пауков) и, по-видимому, возникли из них, хотя как органы не гомологичны легким. В целом у арахнид трахейная система гораздо менее развита, чем у насекомых, обычно не наблюдается у них и дыхательных сокращений брюшка, которые так характерны для многих насекомых.
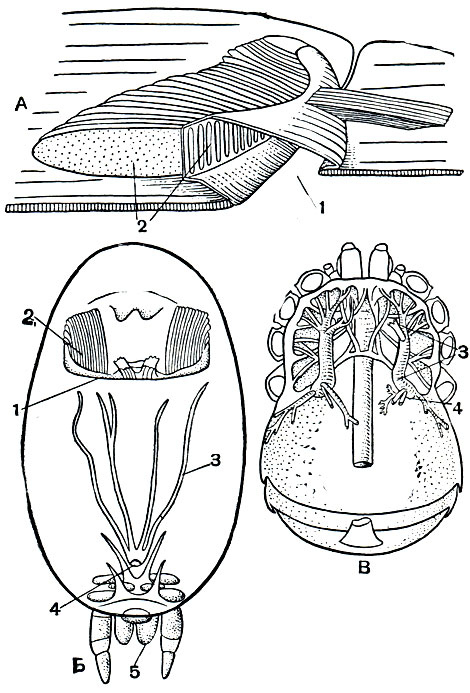
Рис. 17. Легкие и трахеи арахнид: А - схема строения легкого; В - легкие и трахеи двулегочного паука; В - трахеи сенокосца; 1 - легочная стигма; 2 - легочные карманы; 3 - трахеи; 4 - стигмы трахей; 5 - паутинные бородавки
Кровеносная система хорошо развита у крупных форм, дышащих легкими. Имеется пульсирующий спинной сосуд - сердце с несколькими парами боковых отверстий - остий, снабженных клапанами. От сердца отходят передняя и задняя аорты и несколько посегментных пар артерий, которые ветвятся. Кровь (гемолимфа) из сердца через артерии изливается в систему лакун - пространств между органами, собирается в легочные синусы, обогащается кислородом в легочных карманах, по легочным венам возвращается в околосердечное пространство и через остии - в сердце. По мере перехода от легочного дыхания к трахейному кровеносная система становится менее развитой, уменьшается число артерий и остий сердца. Так, у скорпионов и большинства жгутоногих остий 7 пар, у сольпуг - 6, у пауков - от 5 до 2, у сенокосцев - 2 пары, у клещей сердце в виде маленького мешочка с парой остий или оно отсутствует. Кровь обычно бесцветна, в ней имеются кровяные клетки нескольких типов.
Арахниды раздельнополы. Половые железы - яичники и семенники - расположены в брюшке и в исходном состоянии парны. В ряде случаев наблюдается объединение правой и левой гонад. Так, у самцов скорпионов семенники парные, каждый состоит из двух трубок, связанных перемычками; у самок яичник один и состоит из трех трубок, из которых средняя есть результат продольного слияния двух трубок. У многих арахнид парные гонады срастаются концами в кольцо. Парные яйцеводы и семепроводы открываются непарным половым отверстием на восьмом сегменте. Устройство выводной части половой системы и совокупительные приспособления разнообразны. У самок обычно имеется расширение яйцеводов - матка и семеприемники, в которых сохраняется сперма.
Биология размножения разнообразна. Наружное оплодотворение, свойственное водным хелицеровым, сменяется на суше внутренним, сначала свободным сперматофорным, а далее различными способами копуляции. При сперматофорном оплодотворении сперматозоиды заключены в специальный мешочек - сперматофор, выделяемый самцом и защищающий сперму от высыхания. В наиболее примитивных случаях, у многих клещей, живущих во влажной почве, ложноскорпионов, самцы оставляют сперматофоры на субстрате, а самки захватывают их наружными половыми органами. При этом особи совершают характерные взаимные движения - брачные танцы. У многих арахнид самец тем или иным способом переносит сперматофор в половое отверстие самки, что чаще делается с помощью хелицер, имеющих для этого особые приспособления. Наконец, у ряда форм сперматофоров нет, и сперма вводится с помощью специальных копулятивных органов. Последние образуются либо в составе собственно наружного полового аппарата, либо для копуляции служат совсем иные органы, например концевые членики щупалец педипальп у самцов пауков, третьей пары ног у рицинулей. Копуляция сопровождается подчас очень сложным поведением партнеров и проявлением целой цепи инстинктов, особенно у пауков.
У некоторых клещей наблюдается партеногенез, т. е. развитие неоплодотворенных яиц. Иногда самцы появляются периодически, а в остальное время развитие партеногенетическое. Есть также формы, у которых самцы вообще неизвестны.
Арахниды в большинстве откладывают яйца, но есть и живородящие формы (часть скорпионов, некоторые клещи и др.). Плодовитость очень различна. Чаще она составляет несколько сотен яиц, но есть некоторые паразитические клещи, у которых самка откладывает до 30 000 яиц. Обычно яйца откладываются общей кладкой по нескольку десятков или сотен. В течение жизни бывает одна или несколько кладок, что зависит от биологии вида и длительности жизни самки. Среди мелких арахнид есть формы, у которых яйца созревают и откладываются по одному; общая плодовитость при этом обычно невелика. Яйца арахнид богаты желтком. В связи с этим основные процессы формирования организма протекают в эмбриональный период, и из яйца вылупляется особь, похожая на взрослую, только меньшего размера. Послезародышевое развитие прямое, без метаморфоза, сопровождается ростом и линьками. Этот тип развития унаследован арахнидами от водных хелицеровых, у которых, как говорилось, произошла эмбрионизация первичного метаморфоза. Только у клещей в связи с малыми размерами и меньшим запасом желтка в яйце послезародышевое развитие представляет своеобразный метаморфоз.
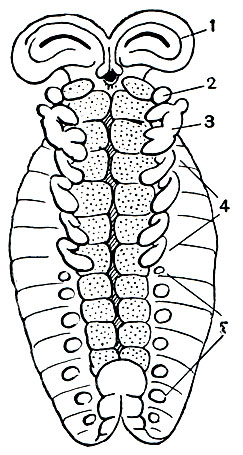
Рис. 18. Зародыш скорпиона: 1 - головные лопасти; 2 - хелицеры; 3 - педипальпы; 4 - ходильные ноги; 5 - зачатки брюшных конечностей
В связи с большим запасом желтка дробление яйца в большинстве случаев поверхностное: ядра, делясь, выходят на поверхность желтка, где образуется слой клеток (бластодерма). Желток при этом обычно не делится. Зародышевые листки арахнид были открыты впервые у скорпионов в 1870 году И. И. Мечниковым и в дальнейшем найдены у других форм. Изучение эмбрионального развития позволяет лучше понять строение взрослых форм. Например, в тех случаях, когда у взрослых исчезает сегментация, она бывает выражена у эмбриона (пауки и др.). В эмбриональном развитии удается проследить, как зачатки конечностей брюшка превращаются в легкие и другие органы и т. д. Большой интерес представляет эмбриональное развитие низших клещей, сохранившее примитивные черты, на чем мы остановимся в дальнейшем.
У многих арахнид наблюдается охрана потомства. Самка откладывает яйца в специально вырытую норку и остается при них. У пауков яйца оплетаются паутинным коконом, который, обычно самка охраняет в гнезде или носит при себе. Вылупившиеся молодые особи обычно в первое время не питаются активно, пищей им служит эмбриональный желток, остающийся в кишечнике. Молодь в этот период держится в гнезде или на теле матери (у скорпионов, телифонов, ряда бродячих пауков и др.) и, только перелиняв, переходит к самостоятельной жизни.
По общему характеру жизненного цикла арахниды весьма различны. В этом отношении можно наметить два типа, между которыми имеются переходы. Один крайний тип представляют крупные долговечные формы, живущие по многу лет и периодически размножающиеся. Таковы, например, некоторые тропические скорпионы, жгутоногие, крупные пауки-птицеяды. Среди последних некоторые живут до 20 лет и всю жизнь не теряют способности линять. При этом типе жизненного цикла индивидуальное развитие длительное и половая зрелость достигается после долгого роста. Особи обычно не образуют массовых скоплений, и вообще численность таких форм в природе относительно невелика. Этот долговечный способ жизни, связанный с крупными размерами или даже гигантизмом и многократным периодическим размножением, очевидно, унаследован арахнидами от водных хелицеровых и вообще не характерен для наземных членистоногих. Среди водных форм меростомовые, а также многие крупные ракообразные по жизненному типу именно таковы. На суше этот тип удержался только у некоторых арахнид, живущих главным образом во влажных тропиках, где условия жизни, так сказать тепличные. Среди трахейнодышащих известную аналогию представляют некоторые гигантские тропические многоножки - кивсяки. Заметим, что среди наземных животных по пути долговечной жизни при крупных размерах особей пошли позвоночные, но у них имелись к тому свои особые биологические предпосылки.
Для большинства арахнид характерен другой, противоположный жизненный тип, который в своих крайних вариантах представлен у многих клещей. Эти мелкие арахниды недолговечны, но зато развиваются очень быстро, причем поколения следуют друг за другом, пока есть подходящие условия. Как только условия становятся неблагоприятными, все активные особи погибают, но остаются покоящиеся яйца или специальные формы (молодые или взрослые), способные переносить неблагоприятные условия (высыхание, низкую температуру, отсутствие пищи и т. п.). При наступлении подходящих условий покоящиеся формы пробуждаются, начинается активная жизнь, размножение, и в короткий срок численность восстанавливается. Этот эфемерный тип жизни, связанный с малыми размерами, высокой скоростью развития и обычно с наличием специальных переживающих стадий, очень характерен для наземных членистоногих вообще, в частности для насекомых. Он, несомненно, есть важнейшее биологическое приспособление к жизни на суше, где условия гораздо более изменчивы, чем в море. Помимо всякого рода случайных изменений среды, при выработке этого жизненного типа сказываются периодические сезонные явления, особенно резкие в умеренном климате. Большинство арахнид, например пауков, подобно многим насекомым, представлено односезонными формами, которые в течение лета успевают проделать одну генерацию, Зимуют обычно яйца или молодь, которая размножается в следующем году. Реже у арахнид бывает 2-3 генерации в год, и только некоторые клещи успевают проделать много генераций.
Не вызывает сомнений, что все арахниды произошли от водных хелицеровых. Как мы видели, переход к жизни на суше сопровождался выработкой многих приспособлений. Жаберное дыхание сменилось легочным, а далее стало дополняться и заменяться трахейным. Число сегментов тела сократилось, брюшко концентрировалось как единый отдел. Произошла дальнейшая специализация конечностей головогруди. Ноги потеряли жевательную функцию, лапки расчленились, и возникло стопохождение. Широкое распространение получило внекишечное разжижение пищи, и околоротовые конечности приспособились к этому своеобразному способу питания. Дифференцировалась сложная система кожных органов чувств, особенно осязательных" Изменения произошли и во .внутреннем строении - концентрация нервной системы:, дополнение и замена выделительных кексальных желез мальпигиевыми сосудами, сокращение кровеносной системы в связи с переходом к трахейному и кожному дыханию, особенно у мелких форм, и др. Изменилась биология размножения. Водный тип наружного оплодотворения сменился внутренним, сначала свободным сперматофорным, а далее разнообразными способами копуляции. В ряде случаев возникло живорождение, охрана потомства.
Выработался эфемерный тип жизни, свойственный наземным членистоногим: способность заканчивать развитие в ограниченный срок, недолговечность и сравнительно малые размеры взрослой формы, наличие переживающих стадий. Так была решена задача перехода к сухопутной жизни.
Однако, как говорилось выше, предки арахнид были достаточно специализированными водными хелицеровыми, и при выходе на сушу новые приспособления могли развиваться лишь на основе уже сложившейся, весьма своеобразной организации водных форм, что создало ряд ограничений. И если взглянуть на арахнид не с обычной точки зрения - восхищения совершенством приспособлений к окружающей среде, а с противоположной - с точки зрения ограничений и трудностей, которые создались благодаря былой специализации и которые пришлось преодолевать или обходить, то многое в их эволюции станет более понятным. Очень показательно также сравнение с насекомыми - трахейнодышащими животными, наземными по природе. Так, дыхание с помощью легких, образовавшихся из жаберных ножек, у членистоногих с их незамкнутой кровеносной системой представляет гораздо менее совершенный способ газообмена, чем трахейное дыхание. Защита от пересыхания - главной опасности на суше- при локализованном легочном дыхании несовершенна, и действительно, большинство арахнид для дыхания нуждается в сильно увлажненном воздухе. Поскольку арахниды встали на путь легочного дыхания, трахейная система не развивалась в должной степени. Несмотря на многочисленные попытки в этом направлении, она не достигла такого совершенства, как у насекомых. Только сольпуги и сенокосцы по степени развития трахей несколько напоминают последних. Характерно, что мелкие тонкокожие арахниды (многие клещи, кенении), живущие во влажном почвенном воздухе, вообще избавлены от противоречивого по природе легочно-трахейного аппарата и дышат через покровы. Многие ограничения в жизни на суше создались благодаря отсутствию обособленной подвижной головы с усиками и челюстями и в особенности атрофии сложных глаз. Арахниды были вынуждены пойти по пути совершенствования в основном осязания, имитируя усики конечностями, и ориентации в окружающем мире "на ощупь", что, помимо прочих неудобств, ограничивает эффективность охоты бродячего хищника. Вместо питания с помощью набора специальных ротовых конечностей - челюстей, приспособляемых для приема разнообразной пищи, что свойственно насекомым, у арахнид выработался очень однотипный способ питания разжиженным содержимым жертвы, т. е. почти поголовное хищничество. Только часть клещей сумела выбраться из этого однообразия. Прямое укороченное послезародышевое развитие, связанное с обилием желтка в яйце и поздним вылуплением, при всех преимуществах имело ту отрицательную сторону, что на его основе не могли возникнуть сложные формы метаморфоза, какие свойственны насекомым и открыли перед ними широчайшие возможности приспособления к различным условиям жизни. Только клещи с их своеобразным метаморфозом в этом отношении стали соперничать с насекомыми.
В том, как и в какой степени преодолевались или обходились эти исторически сложившиеся ограничения, отряды арахнид различны. Эволюционные возможности арахнид наглядно обнаруживаются при сопоставлении видового разнообразия и распространения отрядов. Из общего числа 35 000 видов львиная доля приходится только на пауков (20 000) и клещей (10 000). Из остальных 5000 видов 2500 составляют сенокосцы, 1100 ложноскорпионы, а прочие насчитывают по нескольку сотен или даже десятков видов. Такие соотношения не случайны. Маловидовые отряды - это как раз арахниды, в образе жизни и распространении которых отчетливо проявляются те ограничения, о которых только что говорилось. Все они теснейшим образом связаны с почвой и различными укрытиями, где воздух достаточно влажен. Это бродячие хищники, в большинстве ночные, которые ловят добычу "на ощупь" и прячутся днем в трещинах почвы, под камнями, в норах или постоянно живут под пологом растительности, в лесной подстилке, древесной трухе и т. п. В своем распространении эти отряды ограничены теплыми странами, многие формы не выходят за пределы тропиков. Только часть видов сенокосцев и ложноскорпионов встречается в умеренных широтах.
Иную картину представляют пауки и клещи. Среди арахнид по существу только они сумели вполне преодолеть или, вернее сказать, обойти историческую ограниченность своего класса. Немногочисленные примитивные представители этих групп - низшие норовые и бродячие пауки и примитивные клещи - по экологическому облику еще стоят в одном ряду с другими арахнидами, но дальнейшая судьба пауков и клещей совсем иная.
Решающее значение в эволюции пауков имела паутина, которая первоначально использовалась для устройства яйцевых коконов и выстилания убежищ, а далее стала применяться и для постройки ловчих сетей. В жизни высших тенетных пауков паутина - это все. Это убежище и ловушка. В убежище создается благоприятный микроклимат, особенно важный для дыхания, здесь паук подстерегает добычу, укрывается от врагов и непогоды. Добыча попадает в ловчую сеть, схватывается "на ощупь" при минимальном участии зрения и убивается с помощью хелицер, которыми впрыскивается яд. На паутине происходит спаривание, из нее сплетается яйцевой кокон, в ней укрывается неокрепшая молодь, на паутинках молодые паучки разносятся ветром и т. д. Обеспечив себя всем необходимым, пауки со своими паутинными приспособлениями проникли во всевозможные места обитания, широко заселили сушу и достигли небывалого расцвета. При довольно стереотипном общем облике высшие тенетные пауки исключительно разнообразны по местам обитания, форме и окраске, конструкциям ловчих сетей и повадкам. По сложности поведения и совершенству инстинктов пауки напоминают насекомых. Совсем иным путем обошли затруднения клещи. Решающее значение в их эволюции имели микроскопически малые размеры. Прежде всего клещи заселили почву и гниющую растительную подстилку. Другие, более крупные арахниды либо роют норы, либо отыскивают достаточно объемистые временные укрытия, которые приходится покидать при выходе на охоту. Клещам этого не нужно. Они так малы, что для них пространства между частичками почвы - это готовая система пещер и галерей. А условия жизни здесь весьма подходящие: воздух влажен, пища - мелкие беспозвоночные и микрофлора - всегда в изобилии. В почве, лесной подстилке, скоплениях всевозможных гниющих остатков развился целый мир разнообразнейших клещей, который служил и продолжает служить источником новых форм в их дальнейшей эволюции. Выход из почвы происходил по нескольким направлениям. Одни устремились на растения, другие на животных в качестве паразитов, третьи заселили пресные воды и море. Паразитизм получил очень широкое распространение, и по своеобразию его форм клещи превзошли даже насекомых. Малые размеры и тут сыграли решающую роль. Наряду с наружными паразитами, сосущими кровь животных или сок растений, возникли такие своеобразные микроскопические формы, как чесоточные зудни, живущие в коже, железницы, живущие в волосяных сумках млекопитающих, паразиты трахей насекомых, галлообразующие клещи, обитатели вызываемых ими патологических разрастаний тканей растений - галлов, и многие другие.
Существенное значение в эволюции клещей имело разнообразие питания, возникшее уже на почвенной стадии их жизни. В почве клещам представился неограниченный выбор пищи, разнообразнейшая микрофлора, бактерии, дрожжи, низшие грибы, водоросли, всевозможные мелкие беспозвоночные. Для более крупных арахнид вся эта мелюзга не представляла бы ценности, но клещам пришлась по вкусу. Одни из них стали питаться растительной пищей, грибами, гниющими остатками с обильной микрофлорой, другие - беспозвоночными, мелкими червями, членистоногими, в том числе другими видами клещей. Разнообразие способов питания привело к специализации ротовых органов, возникли грызущие и колюще-сосущие формы. У большинства клещей хелицеры и педипальпы объединились в особый отдел - головку, или гнатосому, чем была обеспечена подвижность ротового аппарата как целого и в какой-то мере компенсировалось отсутствие подвижной головы, столь характерное для хелицеровых. Разнообразие питания отразилось и при переходе к паразитизму. Грызущие потребители органических остатков дали начало многочисленным паразитам птиц и млекопитающих, питающимся кожей, пером, волосом, кожными выделениями. Хищники, высасывающие мелких членистоногих, перешли к сосанию крови позвоночных. Сосущие растительноядные формы заселили высшие растения.
Как мы говорили, благодаря малым размерам яиц клещи развиваются с метаморфозом. По мере приспособления к новым условиям изменялась не только взрослая форма, но и способы метаморфоза, и это значительно расширило эволюционные возможности. В частности, возникли чрезвычайно быстро размножающиеся формы, способные в кратчайший срок достигать колоссальной численности, выработались специальные переживающие и расселительные стадии и т. п. По разнообразию и обилию в природе клещи превзошли пауков, хотя и уступают им по числу известных видов.
Таким образом, большинство отрядов арахнид оказалось ограниченным в освоении суши, и только пауки и клещи пошли значительно дальше и из бедных поселенцев превратились в завоевателей суши. Пауки и клещи распространены очень широко, от тропиков до полярных стран и высокогорий. Их можно встретить там, где жизнь скудна и почти нет даже насекомых. По численности в природе они не уступают последним. Не следует, однако, думать, что остальные, меньшие по числу видов отряды более похожи друг на друга. Напротив, каждый из них имеет свои неповторимые особенности и свои варианты приспособлений, вполне обеспечивающие жизнь в присущих ему условиях. Только эти приспособления более частного характера и не ведут к таким грандиозным эволюционным последствиям, как у пауков и клещей. Сравнивая отряды арахнид, можно как-то обрисовать лицо каждого.
Так, скорпионы - это древнейшие арахниды, по существу вышедшие на сушу эвриптериды. Минимум приспособлений к сухопутной жизни (легочное дыхание, стопохождение, арахнидный тип хищничества) сочетается у них с очень своеобразными чертами (ядовитый аппарат на конце метасомы, переход к живорождению, вынашивание молоди на себе и др.). По образу жизни и примитивности несколько напоминают скорпионов телифоны и фрины, но эти очень бедные видами отряды более узко приурочены к влажным теплым местам обитания, в основном К тропическим лесам, и отличаются по строению (иное число и положение легких, отсутствие ядовитого аппарата на метасоме и др.). Фрины в то же время имеют настолько много общего с пауками, что их считают беспаутинными родичами последних и называют иначе жгутоногими пауками.
Два отряда - сольпуги и сенокосцы - настолько выделяются по степени развития трахейной системы, что их можно назвать трахейнодышащими арахнидами. Главные трахейные стволы открываются стигмами на брюшке там, где у арахнид бывают легкие, и весьма вероятно, что трахеи здесь возникли из легких, в чем, может быть, и кроется причина такого мощного их развития. В остальном сольпуги и сенокосцы очень различны и далеки друг от друга. У сольпуг мощная трахейная система сочетается с примитивностью организации (полнота сегментации туловища, расчлененная просома, педипальпы, подобные ногам, и т. д.). Подобно большинству арахнид, сольпуги - ночные хищники, прячущиеся днем в укрытиях. Но распространены они в основном в сухих и жарких районах, чрезвычайно подвижны, и даже есть несколько видов, бегающих по песку под палящим солнцем в пустынях. Все это указывает на совершенство регуляции дыхания и водного обмена. Однако сама по себе трахейная система при прочих примитивных арахнидных свойствах, видимо, недостаточна для перехода к более совершенным формам открытой наземной жизни, и видовое разнообразие сольпуг невелико.
Сенокосцы по своему жизненному облику - это самые, если можно так выразиться, насекомоподобные арахниды. Наряду с развитым трахейным дыханием в этом отряде преобладает та панцирная жизненная форма, которая характерна для некоторых нелетающих или мало пользующихся крыльями насекомых, например жуков.
Компактное тельце защищено кожистым или очень твердым панцирным покровом. Сегменты брюшка сомкнуты, а у многих форм их тергиты срастаются с головогрудным щитом в общий спинной щит. В то же время тело сенокосцев как бы подвешено на длинных ногах, которые при невысокой частоте движений обеспечивают большую скорость перемещения: шаг сенокосцев очень велик. Наряду с ночными хищниками среди сенокосцев много видов, активных днем, свободно разгуливающих на ярком солнце, даже и в сухих местностях. Не имея тех преимуществ, которые свойственны богатым видами отрядам, сенокосцы тем не менее широко распространились и достигли значительного разнообразия (2500 видов).
Несколько отрядов мелких арахнид - кенении, ложноскорпионы, рицинулеи - приспособились к скрытой жизни в естественных полостях и трещинах почвы, в лесной подстилке, древесных остатках и т. ц. В этом отношении они напоминают клещей. Однако все они крупнее и не перешагнули ту ступень измельчания, за которой возникла микроскопическая жизненная форма клещей с ее эволюционными возможностями. Кенении и рицинулеи представлены немногими редкими, в основном тропическими видами, ложноскорпионов известно 1100 видов и распространены они шире. Кенении - это типичные обитатели скважин почвы, одни из примитивнейших арахнид, напоминающие, с одной стороны, сольпуг в миниатюре, с другой - некоторых низших клещей. Ложноскорпионы также весьма примитивны, но имеют некоторые очень своеобразные черты: хватательные педипальпы с клешнями, как у скорпионов, чрезвычайно своеобразный способ живорождения и др. Они живут скрыто в лесной подстилке, древесной трухе, под отставшей корой, под камнями и могут расселяться, прикрепившись к насекомым. По-видимому, такой образ жизни способствовал довольно широкому распространению ложноскорпионов, хотя и они явно преобладают в тропиках. Об образе жизни рицинулеи известно мало. Эти медлительные формы с очень твердым покровом замечательны тем, что в их развитии, как у клещей, имеется шестиногая личинка.
Смену мест обитания в эволюции арахнид можно иллюстрировать схемой (рис. 19). Выходя на сушу, арахниды были вынуждены ограничиться влажными местами обитания, в которых и до сего времени живут многие из них. Важнейшим условием выхода на сушу была наземная растительность. Многие нашли приют под ее пологом, другие, в особенности мелкие, заселили продукты разложения растений, органическую подстилку и почву. Выработавшаяся у арахнид способность устраивать логова и норки для себя и своего потомства в сочетании с ночной активностью заметно расширила возможности освоения суши и позволила выйти из-под покрова влажной растительности. Тесная связь арахнид с почвой на этой ступени их эволюции хорошо согласуется с представлениями М. С. Гилярова о переходной роли этой среды при смене водного образа жизни наземным, изложенными в его известной книге "Особенности почвы как среды обитания и ее значение в эволюции насекомых" (изд. АН СССР, 1949).
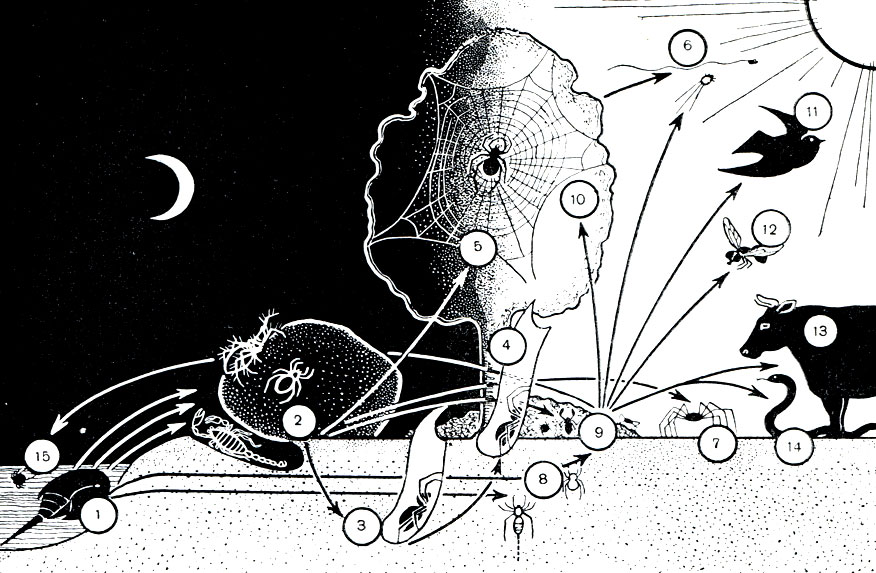
Рис. 19. Выход хелицеровых на сушу (схема): 1 - первичноводные хелицеровые; 2 - ночные бродячие хищники (скорпионы, жгутоногие, сольпуги, низшие пауки и др.); 3 - устройство норок (норковые пауки и др.); 4 - выход пауков из норки с паутинной выстилкой-трубкой; 5 - заселение растительности высшими тенетными пауками; 6 - полет пауков на паутине и подъем клещей токами воздуха; 7 - прямое освоение дневной поверхности суши (некоторые сенокосцы, пауки и др.); 8 - переход из воды к жизни в скважинах почвы (кенении, низшие клещи); 9 - заселение гниющих органических субстратов (многие клещи, ложноскорпионы и др.); 10-14 - выход клещей из почвы и гниющих остатков в качестве паразитов животных и растений, пассивное расселение на хозяевах, по воздуху и иными путями; 15 - водяные клещи
Прямое освоение открытой дневной поверхности суши, как мы видели, у арахнид явление редкое и связано с выработкой надежной защиты от высыхания, трахейным дыханием, уплотнением покровов (часть сенокосцев и немногие другие). Иными, обходными, способами порывают связь с почвой высшие пауки и клещи. Первые уходят от почвы со своими паутинными устройствами, причем наиболее широко заселяют растительность. Вторые благодаря микроскопическим размерам и разнообразию, достигнутому уже в почве, заселяют буквально все мыслимые места обитания, в том числе растения и животных в качестве их паразитов, а также вторично проникают в водоемы. Не имея крыльев, пауки и клещи проникают и в воздушную среду: молодь пауков на паутине разносится ветром, мелкие клещи поднимаются токами воздуха на огромные высоты, будучи паразитами птиц, совершают с ними дальние перелеты.
Чтобы перейти к более подробному обзору отрядов арахнид, необходимо остановиться на некоторых вопросах классификации. Как говорилось, класс Arachnida есть собрание хелицеровых, перешедших к сухопутной жизни. Отряды арахнид очень различны. При глубокой общности их всех как представителей подтипа Chelicerata почти каждый отряд по сочетанию признаков уникален, и не только невозможно вывести его из какого-либо соседнего, но в ряде случаев затруднительно точно сказать, к каким из остальных отрядов он ближе. Такая уникальность отрядов объясняется, с одной стороны, различными вариантами приспособлений к сухопутной жизни, о которых говорилось выше. Но с другой стороны, признаки отрядов таковы, что их невозможно свести к одним только этим приспособлениям, они ведут куда-то глубже и заставляют думать, что арахниды более или менее независимо произошли от различных водных хелицеровых. Непосредственные предки большинства отрядов пока не обнаружены. Но в отношении одного отряда, а именно скорпионов, они теперь известны. Ряд ископаемых переходных форм, независимо от других арахнид, связывает скорпионов с некоторыми силурийскими эвриптеридами. Иначе говоря, класс Arachnida в его традиционном составе приходится считать искусственным. В связи с этим в последнее время не раз делались попытки сгруппировать отряды по их возможному происхождению и разделить арахнид на несколько классов. Но мнения зоологов расходятся, и работу по упорядочению классификации нельзя считать завершенной.
Как говорилось, большинство отрядов арахнид в качестве четких систематических группировок не вызывает сомнений. Разноречия имеются в отношении жгутоногих (Pedipalpi) и клещей (Acarina). С первыми обстоит несколько проще. Под жгутоногими понимают три достаточно четко разграниченные, хотя и близкие в некоторых отношениях группы: телифонов, фринов и тартарид. Большинство авторов вполне основательно считает телифонов и фринов самостоятельными отрядами, Тартарид одни оставляют в составе телифонов, другие, в том числе и мы, считают отдельным отрядом.
Гораздо сложнее обстоит дело с клещами. Клещи представляют огромное собрание мелких арахнид, очень различных по строению и образу жизни и в большинстве сильно изменившихся по сравнению с другими. До недавнего времени все это многообразие объединялось в один отряд Acarina со многими подотрядами и специальными более дробными подразделениями, (когортами, фалангами, сериями и т. п.), систематический состав которых различен у разных авторов. И, пожалуй, нет другой такой группы членистоногих, которая по запутанности и противоречивости классификации была бы подобна клещам. Клещи рассматривались как совсем особые арахниды, которые деградировали и настолько уклонились от исходного состояния, что их даже трудно сравнивать с остальными. Считалось, да и теперь пишется, особенно в учебниках зоологии, что всем клещам свойственны три основных признака, отличающие их от остальных арахнид. Во-первых, сегменты тела у клещей слились и границы между ними исчезли, а если и наблюдается разделение тела на отделы, то последние не соответствуют отделам тела других арахнид. Во-вторых, у клещей имеется особый подвижный передний отдел - головка, или гнатосома, в составе которой объединяются хелицеры и педипальпы. В-третьих, у клещей из яйца вылупляется шестиногая личинка, которая затем превращается в восьминогую форму.
Пока главное внимание исследователей было направлено на изучение отдельных наиболее распространенных групп клещей, особенно паразитических, действительно сильно измененных по сравнению с другими арахнидами, казалось, что по перечисленным признакам клещи сходны между собой и вполне отличимы от других паукообразных. Но со временем стали накапливаться данные о том, что далеко не все клещи так сильно видоизменены. В 30-х годах нашего века французским исследователем Ф. Гранжаном были детально описаны примитивные сегментированные клещи, вполне сравнимые с другими арахнидами. Эти работы привлекли внимание советского зоолога А. А. Захваткина, который в 1941 году в специальном исследовании показал, что отделы тела клещей вполне соответствуют таковым других арахнид, так что первый из упомянутых выше признаков потерял для клещей отличительное значение. Несостоятельным оказался и второй признак: головка, или гнатосома, свойственна не всем клещам, у примитивных форм строение околоротовых конечностей такое же, как у большинства арахнид. Шестиногая личинка была открыта в отряде рицинулей, так что и этот признак потерял для клещей уникальность. Автором этих строк были изучены низшие представители так называемых паразитиформных клещей, и выяснилось, что по строению и типу развития они резко отличны от остальных. В результате создалось довольно странное положение: клещи - один из крупнейших отрядов арахнид - оказались лишенными общих признаков, по которым всех их представителей можно было бы отличить от других отрядов. Это наводило на мысль, что клещи - отряд сборный, что в нем искусственно объединены различные по происхождению арахниды. Анализируя данные о клещах, А. А. Захваткин в 1947 году пришел к выводу, что в действительности существует не один, а три самостоятельных отряда клещей, каждый из которых имеет определенные признаки, четко отличающие его от любого другого отряда. Это акариформные клещи (отряд Acariformes), пapaзитиформные клещи (отряд Parasitiformes) и клещи-сенокосцы (отряд Opilioacarina). Первый отряд сближается с такими примитивными арахнидами, как кенении и сольпуги, а два других более сходны с сенокосцами, рицинулеями и некоторыми ископаемыми отрядами. Основная масса клещей очень естественно и закономерно распределилась между первыми двумя отрядами. Третий отряд представлен лишь несколькими примитивными, в основном тропическими видами.
Дальнейшие исследования подтвердили правильность такого деления. Так, нами было показано, что строение головки (гнатосомы, или, как она теперь называется, гнатемы) различно и что это образование возникло в эволюции каждого отряда клещей независимо, своим особым способом. Исследование эмбрионального развития примитивных клещей показало, что в разных отрядах принципиально различны и способы образования шестиногих личинок, так что этот признак внешнего сходства при более внимательном изучении еще углубил различия. Каждый отряд клещей самобытен не только по своим исходным формам, но и по общему строю эволюции. Из двух многовидовых отрядов акариформные клещи замечательны, с одной стороны, большой примитивностью и древностью низших форм, которые были почти такими же, как сейчас, уже в девоне, а с другой - исключительным многообразием эволюционных ветвей, ведущих к различным паразитам животных и растений, водяным клещам и многим другим. Паразитиформные клещи несколько однотипнее, судя по низшим представителям отряда, они возникли из какого-то иного источника и, вероятно, позже, а главное направление их эволюции - переход от почвенных хищников к кровососам наземных позвоночных и частью сожителям и паразитам насекомых.
Несмотря на фактическую неопровержимость самостоятельности отрядов клещей, новая классификация вызывает различное отношение специалистов. Одни относятся к ней положительно, например, такой выдающийся зоолог и сравнительный анатом нашего времени, как В. Н. Беклемишев, приводит ее в своих "Основах сравнительной анатомии беспозвоночных" (издания 1962, 1964 гг.). Отношение других неопределенно, а некоторых отрицательно. Причины разноречий различны и, как ни странно, мало связаны с фактами. Сказывается главным образом сила традиции. Некоторые авторы пытаются найти выход в том, что, признавая три отряда клещей, объединяют их все в особый подкласс или даже класс. Так поступает, например, наш известный специалист по клещам В. Б. Дубинин в очерке хелицеровых, помещенном в капитальном академическом издании "Основы палеонтологии" (1962). Но такая операция по существу не меняет дела: возвышение ранга не придает естественности объединению клещей. С другой стороны, преобладает чисто формальное отношение к данному вопросу, что обусловлено самым характером изучения клещей. Дело в том, что благодаря многообразию клещей и сложности их исследования подавляющее большинство специалистов занимается отдельными систематическими группами. И для систематика, изучающего, например, только чесоточных или только галловых клещей, не так уж важно, относят ли их к отряду Acariformes или к отряду Acarina. А думать о клещах как о чем-то целом привычнее. Важно и то, что благодаря медицинскому и хозяйственному значению клещей возникла целая самостоятельная отрасль знания, наука о клещах - акарология, параллельная науке о насекомых - энтомологии,- отрасль знания со своими методами, своим кругом научно-практических проблем, сложнейшей терминологией, своими симпозиумами и конгрессами, своими традициями. Но если энтомология имеет объектом естественную группу членистоногих - класс насекомых, то акарология, при новом подходе к клещам, оказывается наукой всего лишь о нескольких разнородных отрядах мелких арахнид. Такое "упразднение" единого объекта целой отрасли знания подчас вызывает и чисто психологический протест.
Совсем иначе предстает разделение клещей на отряды, как только мы от частной и прикладной акарологии обратимся к акарологии общей, задача которой состоит в упорядочении всего огромного материала по клещам, по их строению, развитию, образу жизни, распространению и т. д., ив конечном счете в выяснении происхождения и эволюции клещей. Здесь пути и результаты анализа фактов всецело зависят от того, признаем ли мы клещей единой группой или тремя независимыми отрядами, не более родственными друг другу, чем арахниды вообще. В первом случае мы вынуждены изучать клещей как таковых, отвлекаясь на первых порах от других арахнид, и главные наши усилия направить на то, чтобы представить себе и по возможности найти исходную прототипическую форму для клещей в целом, проследить, как из этого прототипа возникло все многообразие клещей, а затем установить, каковы родственные отношения этого прототипа с другими отрядами. Во втором случае поиски единого прототипа клещей становятся бессмысленными. Мы должны изучать отряды клещей порознь и в каждом случае выяснить исходное состояние, пути эволюции каждого отряда и место каждого в общей эволюции арахнид. И весь фактический материал по клещам с полной убедительностью показывает, что единого прототипа клещей, если можно так выразиться, "проклеща", в природе нет и никогда не было. Традиционный подход к клещам как единой группе ничего хорошего не приносит. Достаточно раскрыть общие монографии по клещам, например наиболее известную объемистую сводку немецкого акаролога Г. Фицтума 1943 года, как мы наталкиваемся на груду фактов, бесконечное перечисление несвязуемых вариантов строения, развития, образа жизни и т. д. Попытки же свести эти данные к чему-то единому неизменно приводят к противоречиям, а подчас к таким фантастическим гипотезам, рассматривать которые здесь вряд ли уместно.
Иные результаты дает изучение клещей как трех независимых отрядов. При таком подходе выясняется, что многообразие клещей не хаотично. В пределах отрядов вырисовываются многочисленные естественные ряды форм, представляющие нам пути минувшего эволюционного процесса. Полнота этих рядов, сохранность в современной фауне исходных, переходных и конечных форм, особенно в отряде акариформных клещей, поистине поразительны, и среди членистоногих вряд ли найдутся другие подобные группы, где современное многообразие так наглядно отражает историю. Сравнительное изучение рядов форм подтверждает и глубокую самобытность отрядов клещей. Ведь если группа организмов едина по происхождению, то по мере того, как мы спускаемся от измененных ее представителей к примитивным, сходство неизменно должно возрастать и все ряды форм сходиться или по крайней мере сближаться. Внутри отрядов именно это и наблюдается: ряды форм сходятся к примитивам. Но при сравнении отрядов клещей друг с другом картина прямо противоположная: чем примитивнее, тем разительнее несходство. Иными словами, клещи представляют яркий пример конвергенции, т. е. внешнего сближения в эволюции нескольких самостоятельных отрядов арахнид. Степень конвергенции клещей очень велика и объясняется общностью их исторической судьбы, в которой, как мы говорили, решающими были малые размеры. Последние повлекли за собой внешне сходные изменения строения и развития (потеря сегментации, образование головки, своеобразный метаморфоз и др.), а также образа жизни (заселение почвы и других пористых субстратов, выход из почвы в качестве паразитов и т. д.).
Говоря о конвергенции клещей, не следует забывать и другую сторону этого явления. До сих пор мы говорили о разнородности клещей как трех отрядов. Но ведь все они хелицеровые и в этом смысле глубоко родственны, как и другие арахниды, так что явления конвергентного сближения отрядов клещей разыгрывались в эволюции на общей им всем арахнидной основе, и в этом также причина глубины конвергенции. Об этом приходится говорить еще потому, что некоторые ученые, отчаявшись понять своеобразие клещей, вообще отделяют их от паукообразных, что представляет другую крайность в вопросах классификации и абсолютно неприемлемо. Как нельзя соединить клещей в одну группу, так нельзя и выкинуть их из паукообразных. Клещи, или, точнее, клещеобразные арахниды, это три самостоятельных отряда, в такой же мере уникальные, как пауки, сенокосцы, сольпуги и другие, и в такой же - мере относящиеся к собранию наземных хелицеровых, именуемому паукообразными. Словом, клещи явились порядочной загадкой, разрешение которой только теперь, после разделения на отряды, встало на твердую почву. В этом отношении клещи служат прекрасным примером того, как классификация организмов есть не только средство их опознавания, или, как некоторые думают, условное "раскладывание по полочкам", но имеет гораздо более глубокое значение. Будучи сама выводом из некоторой, на первых порах ограниченной, группы фактов, естественная классификация придает правильное направление дальнейшему исследованию, избавляя науку от заблуждений и бесплодной траты времени.
Практическая значимость арахнид очень неодинакова. Действительно большое медицинское и хозяйственное значение имеют только клещи. Среди них имеются многочисленные паразиты, передающие человеку и животным возбудителей различных опасных инфекционных заболеваний - энцефалитов, клещевых тифов, чумы, туляремии и др. Многие виды повреждают культурные растения, запасы зерна, муки и других пищевых продуктов. Некоторые почвенные клещи служат промежуточными хозяевами ленточных глистов, поражающих сельскохозяйственных и ценных промысловых животных. Многообразное практическое значение клещей,, как говорилось, вызвало к жизни самостоятельную отрасль знания - акарологию, которая по своим задачам параллельна науке о насекомых - энтомологии.
Наряду с общими разделами эта наука включает и прикладные - медицинскую, ветеринарную и сельскохозяйственную акарологию. В последние десятилетия акарология развивается особенно интенсивно. Многое в этой области сделано советскими учеными. Во всех странах мира число акарологов, занимающихся разработкой только систематики и экологии отдельных групп клещей, по данным каталога, издаваемого в США, превышает тысячу. Значение остальных арахнид более ограниченно и сводится в основном к ядовитым укусам некоторых скорпионов и пауков. Среди последних обращают на себя внимание некоторые тропические виды, а в нашей стране - паук каракурт, распространенный в южных районах. Кроме того, пауки, будучи массовыми хищниками, уничтожающими в основном насекомых, несомненно играют определенную роль в жизни природных сообществ организмов - биоценозов, но они в этом отношении мало изучены.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'