Отряд Усоногие ракообразные (Cirripedia)
Чарлз Дарвин в своей автобиографии писал: "В октябре 1846 г. я начал работать над "Усоногими ракообразными"... я занялся изучением и анатомированием ряда обычных форм, и это постепенно привело меня к исследованию всей группы. В течение восьми лет я непрерывно работал над этим предметом и в конце концов издал два толстых тома, содержащих описание всех известных современных видов, и два тонких in quarto о вымерших видах. Не сомневаюсь, что сэр Э. Литтон-Булвер, выведя в одном из своих романов профессора Лонга, который написал два увесистых тома о ракушках, имел в виду меня".
Усоногие ракообразные, которым отдал восемь лет своей жизни великий натуралист, во многих отношениях замечательны, и интерес к ним Ч. Дарвина вполне понятен. Внешне они даже непохожи на ракообразных. Во взрослом состоянии они ведут сидячий образ жизни, прикрепляясь к всевозможным подводным предметам - скалам, камням, сваям, днищам судов, а также к самым разнообразным морским животным. Тело большинства этих рачков заключено в твердую известковую раковину, состоящую из различного количества отдельных пластинок. Часть этих пластинок подвижно сочленена между собой. Рачок может их раздвигать, и в образующуюся щель просовываются время от времени грудные ножки, совершающие характерные взмахи. При этом вода с находящимися в ней организмами и их остатками загоняется внутрь раковинки; так осуществляется питание и дыхание.
Твердая известковая раковина, постоянное прикрепление к субстрату и отсутствие внешних признаков ракообразных долго мешали выяснению истинной природы усоногих. Даже такой зоолог, как Ж. Кювье, по примеру предыдущих исследователей относил усоногих к моллюскам. Ж. Б. Ламарк рассматривал усоногих как переходную группу от червей к моллюскам. Только в 1830 году, после того как стали известны личинки усоногих, сходные с личинками других ракообразных, была доказана принадлежность усоногих к этому классу.
В настоящее время в этом никто не сомневается. Больше того, нетрудно подметить многие признаки, сближающие усоногих с внешне совершенно несходными с ними веслоногими. Однако усоногие эволюционировали в ином направлении, чем подавляющее большинство остальных ракообразных. Среди непаразитических представителей этого класса только они, вместо того чтобы совершенствовать свою двигательную систему, перешли к прикрепленному существованию и развили мощное средство пассивной защиты - толстую известковую раковину, несколько напоминающую раковину моллюсков. В связи с прикрепленным образом жизни организация усоногих очень сильно изменилась: исчезли наружная сегментация и хитиновый панцирь, бесполезный при наличии защитной известковой раковины, перестроилась голова, которой рачки прикрепляются к субстрату, укоротился и почти исчез брюшной отдел. Изменилась и функция конечностей. Они перестали служить для передвижения и приспособились к тому, чтобы загонять воду в мантийную полость.
Часть усоногих ракообразных перешла от сидяче-прикрепленного образа жизни на поверхности тела различных морских животных к паразитическому существованию, сменив питание планктоном на использование соков своих хозяев. Можно наметить полную серию переходов от свободного существования этих ракообразных к паразитизму. Как и в других подобных случаях, паразитизм вызвал ряд изменений организации усоногих и эти изменения отразились на приспособлениях к прикрепленному образу жизни. Среди паразитических усоногих известны предельно упрощенные формы, утерявшие все признаки не только отряда, но даже класса ракообразных и типа членистоногих. Только изучение строения личинок позволяет понять их истинную природу.
Все усоногие - морские обитатели, причем некоторые из них могут выносить значительное опреснение. Тело их окружено двойной мантийной складкой, соответствующей карапаксу других ракообразных. Мантия непаразитических форм выделяет различное количество известковых пластинок, образующих защитную раковину. Мягкое тело подразделено на головной и грудной отделы. Передние антенны служат для прикрепления к субстрату, снабжены цементными железами и очень сильно укорочены. Задние антенны отсутствуют, ротовые придатки упрощенного строения. На грудном отделе расположено 6 пар двуветвистых ножек. У сверлящих усоногих число грудных ножек сокращено до 4 пар и они одноветвисты. У подотряда корнеголовых ракообразных, все представители которого - паразиты, ротовых придатков и грудных ножек нет совсем.
Из яиц усоногих выходит типичный науплиус, отличающийся от науплиусов других ракообразных присутствием боковых выростов на переднем конце тела, так называемых "рожек". Перелиняв несколько раз, науплиус переходит в стадию метанауплиуса, для которой характерно появление челюстей и зачатков других конечностей и начало развития фасеточных глаз по бокам от науплиального. "Рожки" у метанауплиуса обычно сохраняются, задняя часть тела часто снабжена выростами. За метанауплиальной стадией следует свойственная только усоногим стадия циприсовидной личинки, иногда называемая также "куколкой". Она обладает двустворчатой хитиновой раковиной, покрывающей все ее тело, 4-члениковыми передними антеннами с присасывательными дисками на предпоследнем членике и с цементными железами при основании, упрощенными жвалами и челюстями и 6 парами грудных двуветвистых плавательных ножек. На переднем конце личинки расположены науплиальный и пара фасеточных глаз.
Присутствие двустворчатой раковины придает этой личинке внешнее сходство с представителями отряда ракушковых ракообразных (см. ниже), в частности с относящимся к нему родом Cypris, почему личинка и получила название циприсовидной. Циприсовидная личинка при помощи своих грудных ног свободно плавает в воде, а затем прикрепляется передними антеннами к подходящему субстрату. Дальнейшее ее превращение во взрослого рачка у разных подотрядов происходит по-разному.
В отряд усоногих включают три подотряда, резко различающихся как по строению, так и по образу жизни.
Подотряд Торациковые ракообразные (Thoracica)
Вновь построенный корабль покидает гавань и начинает плавание. Скорость его хода известна, и сначала он идет точно по графику. Однако с каждым днем скорость его уменьшается. На преодоление одного и того же маршрута тратится все больше времени и топлива, и рейсы корабля обходятся все дороже и дороже. В чем тут дело?
Днище корабля обросло различными морскими животными, трение его о воду увеличилось, и скорость хода соответственно уменьшилась. Корабль приходится ставить в док и удалять обрастания его днища.
Подсчитано, что из-за обрастаний судов и необходимости их периодического докования владеющие значительным флотом страны ежегодно терпят миллионные убытки.
Основу обрастаний судов составляют усоногие ракообразные - морские желуди. Они нередко покрывают днище корабля сплошным слоем. С китобойной матки длиной 140 м, плававшей в течение 7 месяцев к югу от экватора, сняли 40 т морских желудей. На наших дальневосточных судах за год плавания нарастает 10-12 кг морских желудей на на 1 м2 поверхности днища.
Морские желуди и относящиеся к тому же подотряду морские уточки (табл. 32), отличающиеся присутствием стебелька, на котором расположено защищенное известковыми пластинками тело рачка, поселяются не только на кораблях. Прибрежные скалы и камни усыпаны ими, они прикрепляются к раковинам моллюсков, к панцирям ракообразных, к раковинам себе подобных, иногда к рыбам. Некоторые виды приспособились к существованию на коже китов и акул. Есть морские желуди, сидящие на скалах значительно выше уровня моря и довольствующиеся брызгами, которые долетают до них только при сильных штормах. С другой стороны, известны усоногие, обитающие на глубине до 6860 м. Всего описано около 700 видов этих ракообразных.

Таблица 32. Усоногие ракообразные: 1 - Lepas anatifera; 2 - L. Fascicularis; 3 - Veruca stromla; 4 - Mitella mitella; 5 - Conchoderma auritum на Coronula diadema; 6 - Scalpellum japonicum; 7 - Balanus improvisus; 8 - B. Evermanni; 9 - B. Balanus; 10 - Chthamalus stellatus; 11 - Xenobalanus globicipitis; 12 - Cruptolepas rachianecti

Таблица 32. Усоногие ракообразные: 1 - Lepas anatifera; 2 - L. Fascicularis; 3 - Veruca stromla; 4 - Mitella mitella; 5 - Conchoderma auritum на Coronula diadema; 6 - Scalpellum japonicum; 7 - Balanus improvisus; 8 - B. Evermanni; 9 - B. Balanus; 10 - Chthamalus stellatus; 11 - Xenobalanus globicipitis; 12 - Cruptolepas rachianecti
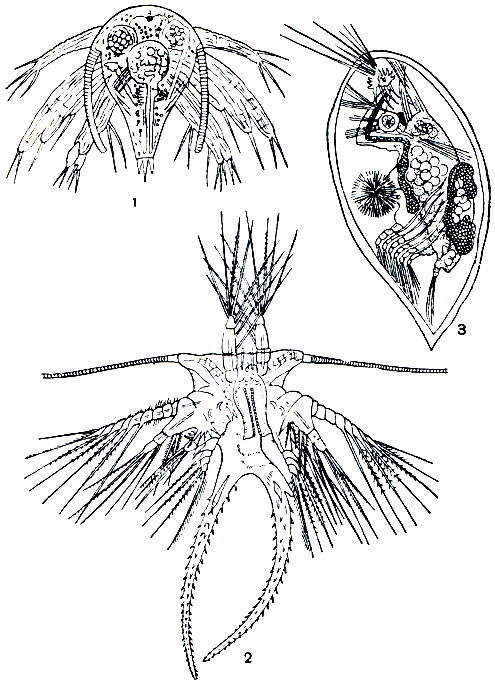
Рис. 222. Личиночные стадии морской уточки Lepas anatifera: 1, 2 - первая и вторая стадии науплиуса; 3 - циприсовидная личинка
Тело морских желудей (семейство Balanidae) и веррук (семейство Verrucidae) целиком находится внутри раковинки, состоящей из нескольких известковых пластинок. Часть этих пластинок неподвижно соединена друг с другом и составляет стенки домика. Другие образуют его крышку. Они могут смыкаться и размыкаться благодаря действию специальных мышц. На дне домика спинной стороной вниз лежит рачок. Передняя часть его сильно видоизмененной головы подогнута под тело так, что остатки передних антенн оказываются приблизительно в центре подошвы. У морских уточек (надсемейство Lepadomorpha) передняя часть головы вытянута в стебелек, голый или снабженный известковыми чешуйками, и только остальная часть тела покрыта известковыми пластинками. Этим стебельком, на конце которого помещаются остатки передних антенн, рачки прикрепляются к субстрату. Задняя часть головы всех Thoracica вздутая. На ней расположен обращенный кверху рот, окруженный верхней губой, жвалами, передними и слившимися воедино задними челюстями. Строение всех ротовых придатков упрощенно, так как они не служат для фильтрации. Глаза очень слабо развиты и представлены раздвоенным науплиальным глазом.
Грудной отдел несет 6 пар двуветвистых ножек. Их размеры увеличиваются по направлению спереди назад. Многочисленные членики ветвей ножек снабжены длинными щетинками на направленной ко рту стороне. Эти ножки, высовываясь из домика, расправляются, как веер, а затем складываются. Они совершают, таким образом, характерные взмахи, загоняя воду и взвешенные в ней частицы в полость домика.
Частота взмахов ножек зависит от внешних условий, в первую очередь от скорости течения воды и от температуры. Обычный морской желудь Balanus balanoides производит 6,2 биения в 10 секунд при 17° С и скорости течения более 0,1 см/сек. При температуре ниже 1,8°С и выше 31,5° С, а также при скорости течения больше 10 см/сек рачок смыкает створки раковины и прекращает движения ножек. Концентрация кислорода и углекислоты в воде не влияет на ритм работы рачков. Щетинки на передних ножках сидят более, а на задних менее часто, благодаря чему разные ножки отфильтровывают частицы разного размера. Поэтому пища усоногих рачков довольно разнообразна: это водоросли, даже бактерии и целый ряд планктонных животных, в первую очередь веслоногих. Захватывают они и собственных личинок, но, по имеющимся наблюдениям, циприсовидные личинки в кишечнике усоногих не перевариваются. Морской желудь Chthamalus, живущий в условиях сильного течения, держит свою ловчую сеть расправленной до тех пор, пока в ней не окажется достаточное количество пищевых частиц.
Весьма своеобразны способ питания и функции грудных ножек морской уточки Lepasfascicularis (табл. 32,2), широко распространенной в тропической и субтропической частях всех океанов. Эти рачки прикрепляются к каким-нибудь плавающим предметам (кусочки дерева, пемза, водоросли) или к выделяемому ими пенистому поплавку и целыми группами плавают на самой поверхности моря. Они могут грести своими ножками и активно подплывать к сифонофорам (Velella, Porpita), которых обгрызают жвалами.
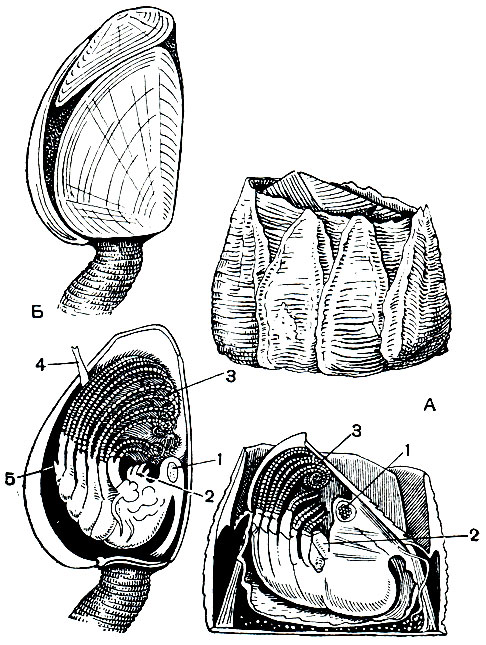
Рис. 223. Строение раковины и тела торациковых: А - морского желудя (Balanus); Б - морской уточки (Lepas); 1 - мускул крышки раковины; 2 - ротовой конус; 3 - грудные ножки; 4 - пенис; 5 - редуцированное брюшко
Есть морские уточки, добывающие себе пищу не прибегая к помощи грудных ножек, которые в таких случаях укорачиваются и упрощаются. Malacolepas conchicola живет в мантийной полости двустворчатых моллюсков и использует пилгевые частицы, поступающие через вводной сифон хозяина. Pagurolepas conchicola селится в раковинах брюхоногих моллюсков, занятых раками-отшельниками, и питается экскрементами последних. Некоторые морские желуди прикрепляются к коже китов и в той или иной степени питаются за счет своих хозяев. Н. И. Тарасов и Г. Б. Зев и на пишут: "Род Coronula не полностью перешел к питанию за счет тканей и крови китов, причем молодые особи, по-видимому, в большей степени питаются биосестоном [взвешенными в воде частицами], нежели взрослые, сросшиеся с кожей хозяина, проникшей в полости стенки их домика. Tubicinella глубоко погружена в кожу кита, а взрослых Cryptolepas невозможно механически отделить от плотно и полностью облекающей их кожи единственного хозяина этого вида - серого кита Eschrichtius gibbosus". У таких специализированных паразитов грудные ножки укорочены, а известковые пластинки домика уменьшены и частично исчезли совсем (табл. 32, 12).
Взмахи грудных ножек служат не только для добывания пищи, но и для обновления воды в полости домика. Это необходимо для дыхания и для удаления из полости домика продуктов выделения. Дыхание совершается через тонкие покровы тела; у некоторых видов имеются нитевидные придатки, которым приписываются функции жабр Сердца ,у усоногих нет и кровь передвигается по полости тела благодаря сокращению мышц.
Весьма своеобразны половая система и особенности размножения Thoracica. В отличие от подавляющего большинства остальных членистоногих эти рачки, за немногими исключениями, гермафродиты. Каждая особь обладает и мужскими и женскими половыми железами. Яйцеводы открываются при основании передней пары грудных ножек. Выходящие из них яйца откладываются в мантийную полость. Семепроводы впадают в длинный трубкообразный мужской совокупительный орган - пенис, представляющий собой вырост редуцированного брюшного отдела и помещающийся непосредственно позади задней пары грудных ножек. При спаривании пенис распрямляется, высовывается из раковины наружу, проникает в мантийную полость соседней особи и выделяет туда сперму, оплодотворяющую яйца. Поскольку обычно морские желуди и морские уточки сидят в непосредственной близости друг к другу, происходящий, таким образом, процесс перекрестного оплодотворения осуществляется без всяких затруднений. В некоторых случаях может происходить самооплодотворение, как это следует из опытов по изоляции отдельных особей, которые и в одиночестве продолжали размножаться.
Только у морских уточек из "семейства Scalpellidae, обитающих на больших океанических глубинах и часто сидящих поодиночке, наблюдаются более сложные явления, на которые впервые обратил внимание Ч. Дарвин. Среди них известны раздельнополые виды; их самки развиты нормально, а самцы в той или иной степени редуцированы (они маленькие, иногда мешковидные, у некоторых сохраняются остатки ножек и пластинок раковины, у других - нет) и сидят по одному или попарно, прикрепившись к стенке мантийной полости самки. Есть и виды-гермафродиты, в мантийной полости которых находятся один или несколько деградировавших "дополнительных" самцов. Наконец, есть виды, нормальные особи которых представлены как самками, так и гермафродитами, причем и в тех и в других живут карликовые самцы.
Некоторые исследователи предполагают, что некогда все усоногие были раздельнополыми. Те виды, которые имеют мало редуцированных самцов, должны считаться наиболее древними, а те, которые совсем их лишены и представлены только гермафродитными особями,- наиболее молодыми.
Недавно дополнительные самцы были неожиданно открыты и у одного из представителей морских желудей - Balanus masignotus с западного побережья Центральной и Южной Америки. По своему строению они напоминают самцов некоторых видов морских уточек, но располагаются снаружи от мантии, в специальной камере, образованной углублением внутренней стенки передней непарной пластинки раковины, а не близ мускула крышки раковины, как у стебельчатых усоногих. Поскольку все морские желуди гермафродиты, присутствие и у них дополнительных самцов осложняет понимание биологической сущности и происхождения этого явления.
Весьма интересен, хотя и до настоящего времени совершенно неясен, вопрос об условиях, определяющих развитие личинки в самца, в самку или в гермафродитную особь. По некоторым наблюдениям, в случае попадания личинки в мантийную полость гермафродитной или женской особи из нее развивается самец, а в случае ее прикрепления к неорганическому субстрату - гермафродитная или женская особь. Эта гипотеза нуждается в дополнительной проверке.
Кучки яиц после оплодотворения в мантийной полости родительской особи объединяются тонкими хитиновыми оболочками в так называемые яйценосные пластинки и подвергаются дроблению. У большинства видов из яиц выходят науплиусы, у некоторых метанауплиусы.
У холодолюбивых морских желудей (Balanus balanoides, B. hameri, B. balanus) яйца образуются летом, оплодотворение их происходит зимой, а личинки выходят на волю весной; теплолюбивые виды (B. improvisus, Elminius modestus, Chthamalus stellatus и др.) успевают отложить яйца несколько раз в течение лета. Один морской желудь (B. balanoides) в наших северных морях за период размножения выпускает 3-4 тысячи науплиусов. Температура и освещение оказывают очень сильное влияние на разные этапы процесса размножения. Так, например, круглосуточное освещение подавляет созревание яиц.
Выход личинок из мантийной полости родительской особи в воду зависит от других причин. У Balanus balanoides он определяется не календарными сроками и не температурой воды, а массовым развитием микроскопических водорослей, которые служат пищей науплиусам морских желудей. Когда эти водоросли размножатся, науплиусы наших северных морских желудей в огромном количестве появляются в планктоне. Они интенсивно питаются и трижды линяют, после чего переходят в стадию метанауплиуса. Метанауплиусы ведут столь же активный образ жизни и тоже трижды линяют, превращаясь в конце концов в стадию циприсовидной личинки. Она не питается, некоторое время плавает, а затем, попав в подходящие условия, прикрепляется к субстрату при помощи своих передних антенн. У тропических видов все личиночное развитие протекает в течение 3-5 суток, у холодноводных видов оно занимает 2 недели - месяц, а то и больше. У многих преимущественно глубоководных Scalpellidae из мантийной полости родителя выходит сразу циприсовидная личинка.
Изучение хода оседания циприсовидных личинок помогает понять особенности распределения взрослых морских желудей и морских уточек как на камнях, скалах и других естественных субстратах, так и на днищах судов, сваях, буях и т. д. По-видимому, разные виды оседают при разных скоростях течения. При скорости течения более 50 см/сек оседание личинок невозможно. Обычно личинки активно движутся против течения, а оседание происходит в момент относительного покоя, т. е. тогда, когда скорость движения течения сравняется со скоростью движения личинки. Личинки Elminius modestus садятся при большей, a Balanus balanoides - при меньшей скорости. Личинки В. crenatus лишены передних антенн и прикрепляются при помощи специальных липких тяжей; поэтому они оседают при незначительной скорости течения. Различные завихрения течений, образуемые ими водовороты вызывают оседание личинок плотными кучками. Кроме того, доказано, что особенно охотно личинки садятся на домики особей того же вида или непосредственно рядом с ними. Только соприкосновение личинки с домиком морского желудя стимулирует ее оседание. Выделяемые взрослым рачком в воду вещества не имеют здесь значения. Благодаря этому возникают плотные одновидовые поселения морских желудей и морских уточек, что для них очень выгодно, так как обеспечивается возможность перекрестного оплодотворения.
Однако существуют наблюдения, показывающие, что личинки морских желудей оседают таким образом, чтобы распределиться равномерно по заселяемой поверхности и благодаря этому избежать ожесточенной внутривидовой конкуренции. Найт-Джонс и Мойз так описывают оседание личинок Elminius на шиферные плиты: "Самые первые особи, заселяющие эти плиты, образуют скопления, руководствуясь, без сомнения, соответствующим инстинктом.
Затем промежутки между скоплениями заполняются и в конечном счете распределение по поверхности становится равномерным. Все же плотность заселения плиты составляет лишь 35-55 особей на 1 см2, тогда как циприсовидные личинки эльминиусов, длина которых не превышает 0,5 мм, а ширина еще меньше, способны заселять ровные поверхности с плотностью до 230 особей на 1 см2 даже в естественных условиях. При тех плотностях, при которых вступает в действие механизм, регулирующий распределение по поверхности, еще остается много места, так что запоздалым пришельцам есть где прикрепиться и передвигаться. Приходится сделать вывод, что рачки продолжают передвигаться, пока не найдут свободное место". Выяснилось также, что личинки B. balanoides распределяются равномерно по отношению к молоди и взрослым особям только своего вида и близких видов, но садятся в непосредственной близости или даже прямо на домик представителя другого семейства - Chthamahis stellatus. Это приводит к ожесточенной конкуренции между Balanus и Chthamalus.
Личинки некоторых видов оседают только на других животных. Есть виды, живущие исключительно в губках (Acasta spongicola, Balanus spongicola и др.) или главным образом на раковинах моллюсков и на панцирях ракообразных, в том числе камчатского краба (B. hesperius в Тихом океане).
В процессе дальнейшего роста молодых морских желудей степень густоты их поселения очень важна, так как определяет форму их домика. В густых поселениях часто он приобретает "лилиевидную" форму, т. е. его диаметр в верхней части больше, чем в нижней, в разреженных поселениях он конический. Возможно, что на форму домика действуют и другие факторы.
Морские желуди растут довольно быстро. В тропиках некоторые виды достигают половозрелости через 8-16 дней после оседания. Обычному в Балтийском, Черном и Азовском морях В. improvisus, а также дальневосточному В. cariosus для этого требуется 3 месяца. Продолжительность жизни разных видов колеблется от 1-2 до 5-7 и более лет.
Как уже указывалось, морские желуди и морские уточки населяют самые различные глубины. Некоторые виды хорошо приспособлены к существованию в приливо-отливной зоне. Они активны только во время прилива, а когда вода отступает, они плотно закрывают крышки домика и погружаются в состояние, сходное с анабиозом. При переносе в аквариум рачки в течение нескольких дней сохраняли тот же ритм, прекращая биение ножек в часы отлива. Однако, даже находясь вне воды, животные продолжают дышать, хотя и менее интенсивно, чем под водой. Только при наступлении резко неблагоприятных условий они впадают в состояние "скрытой жизни" и в течение нескольких месяцев сохраняют жизнеспособность в сухом состоянии, без вреда для себя подвергаются длительному действию пресной воды, а в условиях опыта переносят заливание вазелиновым маслом, содержание в азоте и водороде, т. е. продолжают существовать при полном отсутствии кислорода.
Этой способностью к "скрытой жизни" объясняется, между прочим, появление морских желудей (B. improvisus и B. eburneus) в 1955 году в Каспийском море, где их раньше не было. Они, несомненно, переселились туда из Черного и Азовского морей на днищах судов, прошедших по Волго-Донскому каналу, и сумели пережить длительное воздействие пресной воды. Вероятно, несколько легче было преодолеть еще более длительное расстояние австралийскому морскому желудю Elminius modestus, появившемуся на побережье Англии только в 1943-1944 годах. Он также проделал этот путь на днищах судов, но оставаясь все время в морской воде, а затем стал быстро расселяться вдоль берегов Европы, вытесняя местные виды Balanus balanoides и В. improvisus. Однако при пониженной солености В. improvisus преобладает над новым вселенцем, а при низкой температуре преимущество получает B. balanoides.
Выше остальных морских желудей поселяются виды рода Chthamalus. В Черном море Ch. stellatus (табл. 32, 10) часто и в большом количестве (у Карадага в среднем 28 экземпляров на 1 м2) встречается на прибрежных скалах, выше уреза воды, довольствуясь брызгами, которые иногда до него долетают. Другие морские желуди, в частности обычные виды рода Balanus, образуют массовые скопления в пределах приливо-отливной полосы и во время прилива оказываются под водой. Целый ряд видов морских желудей, как например крупный (до 20 см высотой) дальневосточный В. evermanni (табл. 32, 8), и морских уточек приурочен к области материкового склона. На больших океанических глубинах преобладают морские уточки из рода Scalpellum, к которому относится и наиболее глубоководный из известных видов S. japonicum, обнаруженный в Курило-Камчатской и Японской океанических впадинах до глубины 6860 м.
Некоторые морские уточки прикрепляются не к дну, а к плавающим в воде предметам - кускам дерева, судам, а также к различным животным. Таковы виды родов Lepas и Conchoderma. Обычный широко распространенный вид C. auritum встречается и на кусках дерева, и на днищах судов, и на китах, морских черепахах и медленно плавающих рыбах. При этом C. auritum всегда садится только на твердую поверхность. На китах эта морская уточка находится только на прикрепившихся к их коже морских желудях Coronula (табл. 32, 5) и на китовом усе, а у кашалотов - на зубах, образуя целые гроздья. Уховидные придатки рачка, служащие, вероятно, для отвода профильтрованной воды, сильнее развиты у особей, сидящих на быстро движущихся животных.
Практическое значение Thoracica определяется прежде всего тем, что морские желуди составляют основу обрастаний судов. Как уже указывалось, обросшее морскими желудями (и другими животными) судно теряет исходную скорость. Соответственно увеличиваются время рейса, затраты на топливо и все остальные эксплуатационные расходы. Корабли периодически приходится очищать от обрастаний, а это обходится очень дорого. Подсчитано, что торговый флот США ежегодно терпит убытки от обрастаний, превосходящие 100 миллионов долларов.
Во многих странах усиленно работают над проблемой защиты судов от обрастаний, в частности над изобретением таких красок для днищ кораблей, которые были бы ядовиты для оседающих на суда личинок морских желудей. Некоторые успехи в этом направлении достигнуты, но "необрастающие" краски очень дороги.
Морские желуди вредят не только судам. Проникнув в Каспийское море, они стали оседать наставных сетях, неводах и других рыболовных орудиях и принесли немало неприятностей местным рыбакам, которые раньше с ними не сталкивались.
С другой стороны, несомненно и положительное значение морских желудей и морских уточек. Их личинки нередко составляют существенную часть прибрежного морского планктона и в большом количестве поедаются некоторыми планктоноядными рыбами. Так, например, у берегов Норвегии весной сайда питается преимущественно личинками усоногих, поедает их и беломорская сельдь, а также некоторые другие рыбы. Взрослые усоногие в малой степени используются рыбами, которые часто не могут оторвать их от субстрата и раскусить. Тем не менее морские желуди отмечены в качестве объектов питания трески, пикши, зубатки, осетра и др.
Человек также не брезгает усоногими. Из крупного морского желудя Balanus psittaceus в Чили делают консервы. В Испании и Португалии добывают морскую уточку Mitella pollicipes, которая используется и в пищу, и для приготовления тука. Многие другие усоногие также, несомненно, съедобны. Тур Хейердал, описывая свое замечательное путешествие на "Кон-Тики", рассказывает, между прочим, о том, как он и его спутники варили суп из морских желудей, которыми быстро обрастал их плот, и высоко оценивает вкусовые качества этих рачков. Весьма возможно, что в будущем человек станет использовать усоногих в большей степени, чем ныне.
Подотряд Акроторациковые ракообразные (Acrothoracica)
Занимаясь усоногими ракообразными, Ч. Дарвин открыл один сильно отличающийся по своему строению и образу жизни от морских желудей и морских уточек вид, сверлящий раковины моллюсков, который он считал необходимым выделить в особый подотряд. В настоящее время таких видов известно более десятка. Они точат ходы в раковинах брюхоногих и панцирных моллюсков, а также в стволах кораллов (рис. 224).
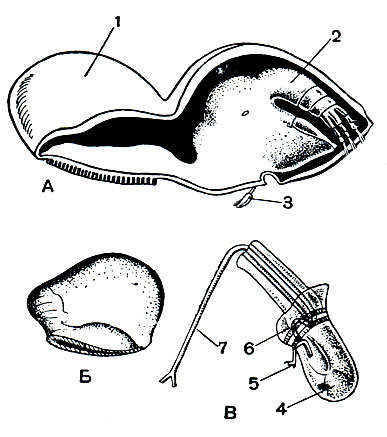
Рис. 224. Строение Tripetosa lampos: А - взрослая самка с карликовым самцом; Б - молодая самка, передняя часть тела которой еще не превращена в диск; В - самец при большом увеличении; 1 - передний конец тела; 2 - грудной отдел с ножками; 3 - самец; 4 - семенник; 5 - передняя антенна; 6 - лопасти мантии; 7 - пенис
Будучи хорошо защищены известковой толщей раковины моллюска или ствола коралла, они, естественно, лишены собственных известковых пластинок, свойственных их только что рассмотренным родичам. Только на передней части головы самки есть хитиновая пластинка, принимающая, вероятно, участие в процессе сверления. Эта передняя часть головы перетяжкой отделена от задней части и от грудного отдела. Вокруг ротового отверстия расположены пара жвал и две пары челюстей, а позади рта пара передних грудных ножек. На значительном расстоянии от них находятся собранные в одну группу остальные три пары одноветвистых грудных ножек. Грудные ножки совершают около 60 взмахов в минуту, но не высовываются при этом за края мантии, как это бывает у морских желудей и морских уточек. Тем не менее благодаря взмахам грудных ножек вода внутри мантийной полости рачка все время сменяется и приносит достаточное количество мелких пищевых частиц. Они поступают в кишечник, лишенный заднего отдела и анального отверстия, но зато снабженный многочисленными ветвящимися слепыми выростами, заходящими в голову, в грудные ножки и в другие части тела.
Все Acrothoracica, за исключением живущего в кораллах рода Lithoglyptes, раздельнополы. Карликовые (0,4-2 мм) самцы живут поодиночке, прицепившись к мантии самки, длина которой достигает 2 см. Они обычно сохраняют облик циприсовидной личинки или имеют очень упрощенное строение. Из оплодотворенного яйца выходит науплиус, такой же, как у Thoracica. Перелиняв три раза, он превращается в метанауплиуса, который дает начало типичной циприсовидной личинке. Циприсовидная личинка садится на подходящий субстрат, сбрасывает свою раковину и превращается в сформировавшегося рачка. Рачок приступает к сверлению известняка, пользуясь хитиновой пластинкой и выделяя вещество, растворяющее соли кальция.
Обыкновенно Acrothoracica селятся в пустых раковинах уже погибших брюхоногих моллюсков (Buccinum, Fusus и др.) или в раковинах, занятых раками-отшельниками .
Подотряд Корнеголовые ракообразные (Rhizocephala)
Еще больше отличаются от обычных усоногих ракообразных паразитические корнеголовые, организация которых предельно упрощена под воздействием паразитизма. Взрослые представители этого подотряда не сохраняют совершенно никаких признаков членистоногих животных. Они живут в десятиногих ракообразных - в крабах, раках-отшельниках и некоторых креветках.
Все тело краба, зараженного корнеголовым рачком Sacculina, пронизано ветвящимися, как корни растений, отростками паразита. Они оплетают все внутренние органы хозяина - кишечник, половые железы, нервную систему, внедряются в мускулатуру. Этими отростками рачки высасывают соки краба. Отростки впадают в обширный мешок, расположенный под брюхом хозяина на поверхности его тела. В мешке находятся сильно развитые яичники и маленькие семенники. Все остальные органы - кишечник, органы чувств и органы выделения - полностью отсутствуют.
Систематическое положение этого существа, так же как и в других подобных случаях (например, описанного выше дендрогастера), выясняется при изучении его развития (рис. 225, Б). Из яиц, откладываемых в полость мешка, выходит типичный для усоногих науплиус, выбирающийся наружу и плавающий в воде. Через 8 дней он превращается в метанауплиуса, который после 4-5 линек переходит в столь же типичную стадию циприсовидной личинки. Все эти личиночные стадии корнеголовых отличаются от соответствующих им стадий других усоногих главным образом отсутствием кишечника. Они питаются за счет содержащихся в их теле запасов желтка. Проплавав 3-4 дня, циприсовидная личинка прикрепляется передними антеннами к основанию щетинок конечностей или брюшной стороны тела молодых крабов. После этого она испытывает удивительные превращения. При очередной линьке она теряет большую часть своего тела со всеми конечностями, но сохраняет мантию, раковину и глаз, а затем линяет еще раз и отбрасывает и их. От личинки остается маленькое овальное мешковидное тельце, внутри которого располагаются недифференцированные клетки, имевшиеся еще у науплиуса. Эта стадия получила название стадии кентрогона. Покровы и передние антенны кентрогона образуют пронизанный каналом вырост, пробуравливающий покровы краба. Через этот канал кучка недифференцированных клеток проникает в тело краба и, вероятно, кровью хозяина переносится на поверхность его средней кишки.
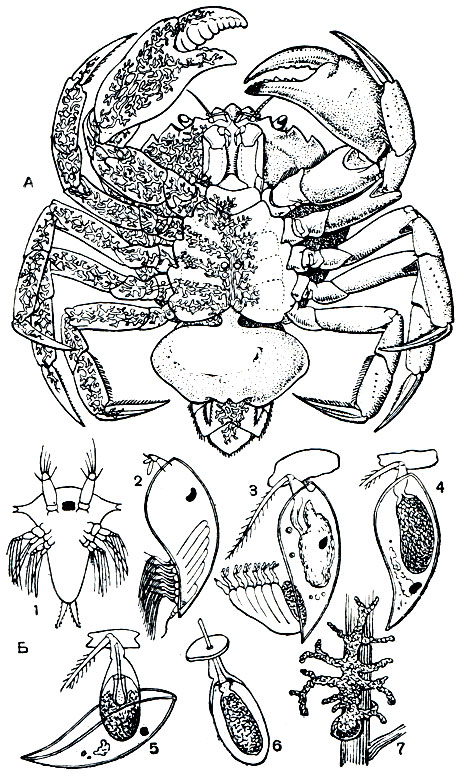
Рис. 225. Sacculina carcini: A - взрослый рачок, паразитирующий в крабе Careinus maenas (левая половина тела краба изображена прозрачной); Б - развитие личинки: 1 - науплиус, 2 - циприсовидная личинка, 3 - ее прикрепление и первая линька, 4 - строение личинки после линьки, 5 - вторая линька и формирование кентрогона, 6 - миграция клеток кентрогона в тело краба, 7 - молодой паразит на поверхности средней кишки краба
Здесь начинается рост паразита. Отростки его тела удлиняются, разветвляются и начинают оплетать внутренние органы краба. Они не разрушают ни кишечника, ни половых желез хозяина, всасывая питательные вещества из омывающей их крови и выделяя в организм хозяина отбросы своей жизнедеятельности. Через 7-8 месяцев паразит образует мешковидный вырост. Когда он достигнет 2 мм в поперечнике, он прорывает покровы хозяина и выпячивается наружу, а затем продолжает расти в течение 6 недель, увеличиваясь до 12 мм в диаметре. К этому времени в мешковидном выросте заканчивается созревание половых желез, которые начинают функционировать. Кроме того, здесь же формируется очень слабо развитая нервная система. Всего для развития взрослой саккулины требуется, таким образом, 9 месяцев с момента прикрепления к хозяину циприсовидной личинки. Паразит живет около 3 лет, и хозяин обычно переживает его и продолжает существовать после смерти саккулины.
Присутствие саккулины оказывает мощное воздействие на организм краба. Заражение саккулиной молодого самца подавляет развитие его половых желез. С каждой последующей линькой он становится все более похожим на самку: брюшной отдел его расширяется, исчезают превращенные в совокупительный орган передние брюшные конечности, меняется характер щетинок брюшка. Японские зоологи Ишикава и Янагимачи описали зараженного саккулиной самца краба Eriocheir japonicus, семенники которого под влиянием паразита переродились в яичники, продуцировавшие нормальные яйца. Если саккулина заразила взрослого крупного самца или самку, это не приводит к изменению вторичнополовых признаков хозяина, но нарушает деятельность его половых желез - происходит паразитарная кастрация.
Нет сомнения в том, что причиной таких изменений в организме краба служит воздействие саккулины на его органы внутренней секреции. Непосредственно или выделяя специфические вещества, паразит подавляет выделение гормонов, действующих на формирование вторично-половых признаков самца и контролирующих деятельность половых органов самца и самки.
Паразиты раков-отшельников Peltogaster и Peltogastrella также вызывают паразитарную кастрацию своих хозяев, но заражение ими самцов не приводит к развитию у них вторичнополовых признаков самок. В отличие от саккулины у этих паразитов есть дополнительные самцы, сохраняющие облик циприсовидных личинок. Семенники гермафродитных особей очень малы и не функционируют. Самцы прикрепляются передними антеннами к мешковидной части гермафродитных особей, и масса заполняющих их недифференцированных клеток переселяется в недоразвитый семенник гермафродита. Часть этих клеток преобразуется в сперматозоиды.
При линьке раков-отшельников наружные мешковидные части их паразитов отваливаются вместе с покровами хозяина. Однако это не приводит к гибели паразита: из корневидной, находящейся внутри рака-отшельника части паразита вновь отрастает его мешковидная часть. У Chologaster наблюдалось четырехкратное отпадение мешков и замена их новыми.
Способность к почкованию особенно ярко проявляется у Thompsonia - паразита креветок из семейства раков-щелкунов. На теле зараженной креветки всегда находится большое количество мешковидных паразитов, объединенных общей корневой системой. Она возникает из одной личинки, а затем образует много мешковидных особей.
Корнеголовые ракообразные широко распространены во всех морях и океанах, за исключением холодных южных и северных морей. Есть они и в Черном море, и в наших дальневосточных морях. В Северной Атлантике, Средиземном и Черном морях около 6% представителей массовых видов крабов заражено саккулиной. В тропиках некоторые корнеголовые вместе со своими хозяевами обитают и в пресных водах. Всего известно около 120 видов этих удивительных ракообразных.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'