Отряд Ластоногие (Pinnipedia) (С. П. Наумов)
Ластоногие, как и китообразные, водные (преимущественно морские и океанические) млекопитающие. Однако связанность с водной средой у них не столь велика, как у китов. Они не могут спариваться и рождать в воде и в период размножения выходят на лед или на сушу. Смена мехового покрова у ластоногих проходит вне воды. Наконец, ряд видов ластоногих выходит из воды и для периодического отдыха. Надо указать также, что пищу они добывают только в воде, чем подчеркивается полнота связи с водой как с обязательной средой обитания.
Ластоногие - немногочисленная (около 30 видов) и очень четко выраженная группа. Сравнительно малые масштабы различий между составляющими ее видами, очевидно, связаны с относительным однообразием жизненной обстановки, в которой возникли и обитают ластоногие, и, быть может, с молодостью отряда, возникшего, видимо, в середине третичного периода. Филогенетически наиболее близкие к ластоногим наземные хищники (отряд Carnivora) имеют значительно более длинную историю, они обитают в крайне разнообразных природных условиях и поражают огромным разнообразием форм.
В большинстве ластоногие - это крупные или средней величины звери, длина тела которых 1,2-6,6 м, а общая масса тела от 40 кг до 3,5 т. Тело удлиненное, веретенообразной формы, округлое в сечении и суживающееся к головному и хвостовому отделам. Характерна форма шеи, которая не отграничена резко от туловища и головы. Шея очень толстая, особенно в задней своей части. По направлению к голове она постепенно суживается, но остается шире последней. У большинства видов подвижность шеи невелика, исключение составляют ушатые тюлени (например, морской лев).
В результате длительного приспособления к жизни в водной среде весьма существенны особенности строения конечностей и их соотношение с туловищем. Значительная часть конечностей скрыта в кожном туловищном мешке, и наружу выступают у передней пары конечностей не более чем половина предплечья и кисть, у задней пары их - только задняя треть голени и стопа, а у некоторых - только стопа. С указанным .сочетается резкое укорочение плеча, предплечья, бедра и голени. Наоборот, кисть и стопа заметно удлинены. Особо характерно наличие толстой кожистой перепонки, связывающей пальцы конечностей, в результате чего они преобразованы в ласты. При этом пальцы кисти снаружи не расчленены, хотя скелет их развит полностью. Пальцы задних конечностей (стопы) снаружи, наоборот, явственно видны. Когти развиты у разных видов неодинаково: у ушатых тюленей и моржей они малы, иногда рудиментарны; у настоящих тюленей когти развиты хорошо, особенно на пальцах передних ластов.
В отличие от своих ближайших родственников - наземных хищников - ластоногие используют конечности только для передвижения и не могут удерживать ими кормовые объекты. Наиболее полно для гребли приспособлены конечности настоящих тюленей. Только передними ластами они могут опираться о твердый субстрат, а задние у них вытянуты назад параллельно продольной оси тела. Наоборот, моржи и ушатые тюлени способны опираться о субстрат и задними ластами (рис. 209). У всех видов основным органом движения в воде служат задние ласты. Вытянутые при плавании назад, они то сближены между собой, то несколько раздвинуты и действуют как более или менее единое целое. Совершая в основном колебательные движения вправо и влево и одновременно несколько изгибаясь сверху вниз, задние ласты своей работой отчасти напоминают гребной винт. Именно они сообщают телу поступательное движение. При работе задних ластов их собственная мускулатура играет сравнительно небольшую роль и основную нагрузку несут мышцы задней части туловища, производящие энергичное колебательное движение позвоночника.
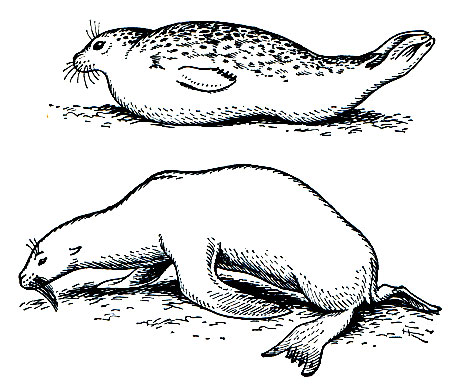
Рис. 209. Характер движения настоящего тюленя (вверху) и ушатого тюленя (внизу) но твердому субстрату
Передние ласты при плавании служат преимущественно для уравновешивания тела; они уточняют работу как горизонтальные и вертикальные рули, дифференцируют движение вперед, возникшее в результате действия задних ластов.
Кожа ластоногих по сравнению с наземными хищниками значительно толще, но покрыта низкой, жесткой шерстью, состоящей (у взрослых) почти исключительно из остевых волос. Особенно сильно редуцирован меховой покров у моржей. Наоборот, у котиков он относительно густой, с хорошо выраженным (особенно у молодых) мягким, густым подшерстком. В той или иной мере выраженная редукция мехового покрова - несомненное приспособление к длительному пребыванию в воде. Ведь в этих условиях мех не может выполнять свою основную роль - теплоизоляцию тела животного. Функцию защиты от охлаждения берет на себя слой подкожного жира, особенно сильно развитый у видов, населяющих холодные страны и выходящих для размножения и линьки на лед. Развитость подкожного жирового слоя (как и развитость мехового покрова у наземных хищников) варьирует по сезонам. Наиболее толстым он бывает зимой, наименее толстым - летом. Как пример приведем сезонные изменения средней массы сала (с кожей - так называемой хоровины) у каспийского тюленя (в процентах к наибольшей массе, наблюдаемой в феврале):

В жизни ластоногих, как и китообразных, подкожный жировой слой играет не только теплоизоляционную, но и гидростатическую роль. Плотность их тела в среднем близка к 1. Зимой в связи с увеличением ожиренности он становится менее единицы, и убитые на воде тюлени, как правило, не тонут. Раненные на воде тюлени нередко в конвульсиях захлебываются, и вода, попавшая к ним в легкие, увеличивает плотность тела, в связи с чем животное может утонуть.
Подобно наземным хищникам, у ластоногих бывает сезонная смена волосяного покрова, но существенной разницы между зимним и летним мехом у них нет.
В строении скелета, помимо уже описанных особенностей костяка конечностей, обращает внимание отсутствие трубчатости костей и ключицы; последнее находится в связи с однообразными движениями конечностей, совершаемыми преимущественно в одной плоскости. Зубная система относительно упрощенная, так как у ластоногих зубы предназначены для схватывания и удерживания кормовых объектов, а не для пережевывания пищи. Особенно это заметно на строении коренных, которые имеют более или менее острые вершины, расположенные часто в одной продольной плоскости. Хищнический зуб не выражен. Клыки хорошо развиты, резцы, наоборот, у большинства в той или иной мере редуцированы.
Мозговая коробка относительно крупная и несколько уплощенная. Лицевой отдел черепа укорочен и сужен. Особенно сужено пространство между глазницами, в связи с чем глаза направлены более вперед, чем в стороны. Головной мозг велик, и полушария переднего мозга имеют многочисленные борозды.
Подавляющее большинство ластоногих без наружных ушных раковин. Исключение составляют виды семейства ушатых тюленей, у которых они хотя и есть, но столь рудиментарны, что не имеют функционального значения. Однако слышат все ластоногие хорошо как в воздушной, так и в водной среде. При погружении в воду наружное слуховое отверстие рефлекторно замыкается специальными кольцевыми мышцами. Доказана (для некоторых видов) способность ластоногих к эхолокации, которая осуществляется при плавании подо льдом и при добывании пищи. Но сравнительно с дельфинами ластоногие издают слабые подводные звуки (щелчки). Вероятно, они могут использоваться животными для эхолокации на небольших расстояниях на последних критических стадиях поимки добычи. У морского льва длительность каждого импульса составляла 3-5 м/с. Основная частота заполнения импульса изменялась от 3 до 13 кгц, будучи в среднем равной 7,5 кгц.
Обоняние вполне удовлетворительное. При промысле тюленей часто наблюдают, что при подъезде на лодке к зверям, лежащим на льдах, по ветру, при почти абсолютной тишине, они уходят в воду с расстояния 150-200 м. Наоборот, подъезжая к ним с такой же степенью осторожности, но против ветра, обычно удавалось приблизиться на 10-20 м, а иногда и ближе. Это было возможно при условии хорошей маскировки: лодка и гребки были выкрашены в белый цвет, а зверобои были одеты в белые халаты.
Зрение, в отличие от обоняния, развито слабо: роговица плоская, хрусталик круглый, способность к аккомодации незначительная. Все это говорит о близорукости, столь типичной для водных позвоночных. У всех видов ластоногих имеются вибриссы - длинные щетинообразные волосы, расположенные на лицевой части головы и служащие главным образом органом осязания.
При добыче корма ластоногие вынуждены более или менее длительное время быть в толще воды. Продолжительность пребывания в погруженном состоянии обычно равна десяткам секунд или нескольким минутам, но в отдельных случаях она может быть заметно большей. Так, максимальная длительность подводного нахождения настоящих тюленей равна 15, а моржей - 16 минутам. Приспособления к нахождению под водой многообразны, но механизм их действия известен только частично.
Установлено, что размеры легких у ластоногих несколько больше, чем у наземных хищников. Так, например, относительная масса легких (в процентах от массы тела) ладожского тюленя равна 2,8, песца - 1,8, енотовидной собаки - 1,5, лисицы - 1,1. Плевра утолщенная, легочная ткань очень эластичная, сильно развита мускулатура, которая имеется даже в стенках альвеол. Легкие в этой связи хорошо противостоят сдавливанию; обеспечивается весьма полное выдыхание использованного воздуха и энергичный прием обильной порции свежего воздуха.
Приспособительные к пребыванию под водой особенности обнаружены и в кровеносной системе. Количество крови у ластоногих заметно большее, чем у наземных зверей, что обеспечивает возможность связывать больше кислорода. Масса крови (в процентах к массе тела) составляет у ластоногих 10-15, у собаки - 6-8, а у свиньи всего 3-5. Повышенная способность связывать кислород обусловливается также обилием мышечного гемоглобина (миоглобина). У тюленей он составляет 50% или несколько больше всего гемоглобина в теле. Во время пребывания под водой мышцы у ластоногих дышат почти исключительно за счет использования кислорода, связанного миоглобином. В это время резко замедляется быстрота тока крови, что обусловлено уменьшением частоты сокращений сердечной мышцы. Наблюдения над серым и обыкновенным тюленями показали, что у зверей, не погрузившихся под воду, число сжатий сердца в минуту равно примерно 180; через 11 секунд после погружения пульс сокращается до 60, через 27 секунд - до 35 и далее в течение всего периода нахождения под водой держался на уровне 30. Все это обеспечивает очень экономное и полное использование кислорода, связанного гемоглобином.
При всей высокой приспособленности ластоногих к жизни в водоемах размножение их возможно только вне воды. Субстрат, на котором происходят деторождение и спаривание, у разных видов различен. Одни из них выходят на сушу, на пляжи островов и материков; таковы ушатые тюлени (котики, сивучи, морские львы) и некоторые из настоящих тюленей (например, обыкновенный тюлень, тюлень-монах). Характерно, что у видов этой группы (обозначаемой как геофилы) подкожный слой жира развит относительно сдабо. Другие (их называют пагофилами) для размножения используют льды. Пагофилы используют плавающие льды открытого моря (гренландский и полосатый тюлени, хохлач) и льды прибрежных мелководий (морж, морской заяц, нерпа).
Ластоногие размножаются медленно, не чаще одного раза в год, и рождают по одному детенышу. Половозрелости достигают, видимо, не ранее чем в 3 года. Новорожденные у многих видов рождаются покрытые длинным густым эмбриональным мехом, резко отличным от меха взрослых не только структурой, но и цветом. Такой мех сохраняется в течение нескольких недель. Новорожденные в этот период не сходят в воду, и подкожный слой жира у них не развит. Молодые растут очень быстро, что обусловлено большой питательностью молока, в котором очень много жиров и отчасти белков.
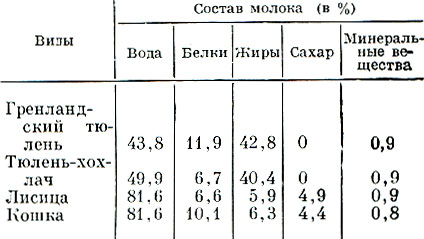
Сравнительный состав молока некоторых ластоногих и наземных зверей
Семейный строй у ластоногих выражен слабо, и мать покидает детеныша после окончания молочного кормления.
Подавляющее большинство видов стадны, хотя бы в течение части года. Наибольшие скопления образуются в период деторождения и линьки, что связано с особой требовательностью зверей к специфической природной обстановке в это время. Степень стадности различна у разных видов. Особо значительные скопления образуют гренландский и каспийский тюлени, хохлач, котики, сивучи. В меньшей мере стадность свойственна нерпе и морскому зайцу.
Филогенетически ластоногие очень близки к наземным хищникам, от которых они отделились в середине третичного периода. Вполне достоверные ископаемые остатки ластоногих известны из отложе-ний нижнего миоцена.
Ластоногие издавна являются объектами промысловой добычи. Их добывает местное население, используя мясо в пищу, а шкуры для изготовления одежды и обуви. Некоторые народности изготовляют из шкур ластоногих лодки. Промысел ластоногих ведут и специализированные организации, используя при этом разнообразные технические средства: самолеты для разведки скоплений зверей, специальные ледокольные или деревянные суда, современное огнестрельное оружие. Основной товарной продукцией являются жир и шкуры, используемые в качестве сырья для кожевенной промышленности. При добыче молодых зверей некоторых видов (например, гренландского и каспийского тюленей), одетых эмбриональным волосяным покровом, их шкура используется как меховое сырье. Особенно дорого ценится котиковый мех.
Отряд ластоногих распадается на семейства ушатых тюленей, моржей, настоящих тюленей.
Семейство Ушатые тюлени (Otariidae)
По ряду признаков ушатые тюлени в меньшей мере, чем другие ластоногие, отклонились от своей предковой группы - наземных хищников. У них есть хотя и рудиментарные, но все же явственно заметные ушные раковины. Задние ласты могут сгибаться в пяточном сочленении, и, будучи направленными вперед, они, как и передние ласты, служат для опоры туловища и участвуют при передвижении по твердому субстрату (рис. 209). Эти ласты в значительной мере оголены от шерсти и оканчиваются фестончатой кожно-хрящевой оторочкой. В отличие от настоящих тюленей пальцы задних ластов не могут широко раздвигаться. Крайние пальцы не имеют когтей, хорошо развитых на средних пальцах.
Передние ласты очень велики: длина их не менее 1/4 длины тела, а у ряда видов еще больше. Как и задние ласты, они на значительном протяжении от конца оголены. Когтей на пальцах передних ластов нет, или они в зачаточном состоянии.
Зубы довольно хорошо дифференцированы. Молочные зубы сменяются через несколько месяцев после рождения.
Зубная формула:
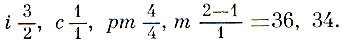
Размеры животных преимущественно крупные: длина тела до 3,8 м. Характерна резкая разница в размерах самцов и самок. Все виды полигамны.
Распространены в умеренных поясах обоих полушарий: в северном полушарии - только в Тихом океане; в южном - во всех океанах (карта 10).
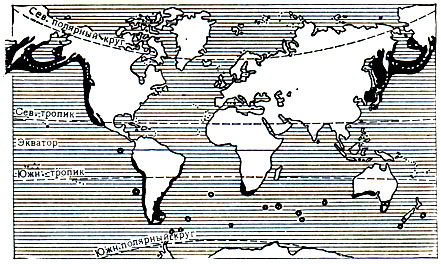
Карта 10. Распространение ушатых тюленей
Ушатые тюлени-типичные геофилы, образующие залежки в сезон размножения только на берегах.
Ископаемые ушатые тюлени известны уже с раннего миоцена из отложений западного побережья Северной Америки, которую считают центром возникновения этого семейства.
В современной фауне 5 общепризнанных родов: северный морской котик (Callorhinus), южный морской котик (Arctocephalus), калифорнийский морской лев (Zalophus), южный морской лев (Otaria), сивуч (Eumetopias).
Северный морской котик (Callorhinus nrsinus) - самый мелкий представитель своего семейства. Длина тела взрослых самцов 135-200 см, масса - 100-188 кг. Самки, как и у других видов этого се-мейства, заметно мельче и более нежного телосложения. Взрослые самки имеют длину тела обычно 110-135 см, изредка до 150 см. Масса их чаще 25-40 кг, изредка наблюдали самок до 61 кг. Общий тип телосложения у котиков массивный, хотя они и не лишены своего рода изящества (табл. 37). Шея у них довольно подвижная и более длинная, чем у других ушатых тюленей. Морда короткая, но заметно заостренная. Глаза маленькие и широко расставленные. Ласты очень длинные, уплощенные. Когда задние ласты отогнуты вперед, их пальцы могут достигать поджатых к телу передних ластов. Когти на передних ластах почти не развиты, на задних когти есть, но только на трех средних пальцах.

Таблица 37 а. Северный морской котик (Callorhinus ursinus). Самец (секач)

Таблица 37 б. Северный морской котик (Callorhinus ursinus). Самка
Волосяной покров весьма своеобразен. Он состоит из двух хорошо выраженных слоев: грубой, жесткой ости и низкой, густой, мягкой подпуши. На 1 см2 шкуры насчитывают около 30-50 тысяч волосков подпуши. Наиболее длинные волосы находятся на задней части шеи. На туловище шерсть короче всего в области крестца. Большая часть передних и задних ластов лишена шерстного покрова. Окраска меха у старого самца (секача) темно-бурая со слабо выраженной рябью, возникающей потому, что многие волосы, особенно на переднем отделе туловища, шее и на затылке, имеют палевые вершины. Подпушь ржаво-бурая.
Мех взрослой самки серо-бурый,более темный у старых и с преобладанием серых тонов у молодых. Неполовозрелые имеют коричневато-серую окраску, более светлую на нижней стороне тела. Новорожденные одеты блестящим буровато-черным мехом, который сохраняется в течение первых трех месяцев, после чего они линяют, и новый мех у них бывает серого цвета.
Описываемый вид котика распространен только в северной части Тихого океана, где он держится преимущественно в районах, не очень далеко отстоящих от берегов. Если учесть очертания этой части Тихого океана, то станет очевидным, что ареал котика рисуется в виде дуги, обращенной вершиной на север. Левая, азиатская часть этой дуги простирается от районов, примыкающих к восточному побережью Камчатки, на юг вдоль Курильской гряды к Японии и берегов Корейского полуострова. В ареал котика входит значительная часть Японского моря, южная часть Охотского моря и омывающие Японские и Курильские острова части открытого Тихого океана. Правая, североамериканская часть ареала в виде широкой полосы простирается от Берингова моря на юг вдоль западного побережья Северной Америки до Калифорнии.
Котики - широко мигрирующие животные, в связи с чем сезонные особенности распространения у них довольно своеобразны. Летом в период размножения большая часть поголовья сосредоточивается в немногих и ограниченных районах: на островах Прибылова, Командорских, острове Тюленьем (у восточного побережья Сахалина), на некоторых Курильских островах и в водах, непосредственно омывающих перечисленные острова. Осенью котики покидают береговые лежбища на островах и откочевывают на значительные расстояния. Во время миграций и зимовки на берег не выходят. Наиболее далеко на юг отходят самки и неполовозрелые звери. Котики командорского стада зимуют у берегов Японии, котики с острова Тюленьего уходят в южные районы Японского моря до берегов Кореи, наконец, звери, размножающиеся на островах Прибылова, мигрируют вдоль побережья Северной Америки и зимуют близ Калифорнии. Самцы- производители (секачи) не предпринимают столь же длительных странствий и по мере наступления зимы лишь несколько отходят к югу. Командорские секачи зимуют несколько южнее Алеутской гряды.
В период размножения большинство половозрелых зверей держится не в море, а на определенных местах побережий островов, которые издавна обозначаются как лежбища (табл. 37). Обычно это участки песчаного или галечного пляжа либо занятые обломками камней, иногда довольно крупными. Лежбища располагаются как на выступающих в море мысах, так и на узкой прибрежной полосе, расположенной иногда под высокими отвесными скалами. Последний тип лежбищ особенно характерен для острова Медного из группы Командорских островов. Обычно на каждом лежбище можно видеть два довольно хорошо выраженных участка: семейные, на которых располагаются секачи, плодующие самки и новорожденные, и холостяцкие, где лежат холостяки, т. е. самцы, не участвующие в размножении. Бывают и такие лежбища, где располагаются только холостяки и не имеющие гаремов секачи.

Таблица 37. Северный морской котик (Callorhinus ursinus). Лежбище котиков
Первыми в районе лежбищ появляются секачи. Это бывает обычно в первой половине мая; наиболее интенсивный подход секачей наблюдается в середине или даже во второй половине мая. Вскоре после появления секачей у участков лежбищ они выходят на берег и располагаются на некотором расстоянии друг от друга. После зимы, в течение которой звери интенсивно кормятся, секачи подходят к лежбищам полные сил и энергии. Они ревниво охраняют занятые ими участки - места будущих гаремов, особенно когда к берегу подходят самки. К этому времени секачи отгоняют с семейных лежбищ холостяков. Эти последние подходят к лежбищам несколько позже секачей: с середины мая до середины июня.
Матки появляются у лежбищ в конце мая - начале июня, а массовый подход бывает в конце июня. В первые же дни появления у островов они выходят на берег, где их поджидают секачи, старающиеся захватить и отогнать в свой гарем возможно большее число самок. Котики - стадные животные, и матки без видимого сопротивления идут в гарем. Бывают случаи, когда два секача, намереваясь захватить одних и тех же самок, вступают в драку. Случается и так, что во время их поединка третий секач, пользуясь этим, отгоняет спорных маток к своему гарему. Гаремные самки ревниво охраняются самцами-владельцами, хотя бывают случаи умыкания маток из гарема. На этой почве также возникают драки между секачами.
Через 1-4 дня после выхода на лежбище самка рождает своего единственного детеныша. Двойни бывают как исключение. Вскоре после родов у самки начинается течка, а через 2-6 дней происходит спаривание. Таким образом, беременность длится около 360 дней. Столь большая ее длительность обусловлена тем, что эмбрион развивается в начале беременности, затем его развитие приостанавливается или, во всяком случае, резко замедляется.
Отношение секачей к маткам в период, их нахождения в гареме различно в зависимости от полового состояния. Особенно ревниво относятся они к беременным и к родившим, но еще не оплодотворенным маткам. Секачи стараются не отпускать их из гарема и упорно удерживают около себя. Наоборот, оплодотворенные матки могут свободно уходить кормиться в море, а между отлучками держаться на суше не более 2-3 дней. В этой связи число новорожденных на лежбище всегда бывает заметно большим (иногда в два раза), чем число находящихся на берегу самок. В начале гаремной жизни секачи находятся на берегу, но уже в середине ее регулярно уходят в море для кормежки или на холостяцкие лежбища для отдыха. Размер гарема по этой причине определить трудно. Во всяком случае, бывают гаремы в 100 самок и даже несколько больше, а наряду с этим наблюдаются секачи, довольствующиеся одной-двумя самками. На Командорах недавно в гареме насчитывались в среднем немногие десятки маток, на островах Прибылова - 30-40 самок.
Самцы достигают половой зрелости на 3-4-м году жизни, и к этому времени семенники у них опускаются в мошонку. Однако семьями секачи обзаводятся значительно позже, в возрасте 8-9 лет. Самки становятся способными быть оплодотворенными уже в возрасте двух лет, но большинство их участвует в размножении лишь с четырехлетнего возраста. Воспроизводительная способность у самок сохраняется примерно до двадцатилетнего возраста.
Котики рождаются, имея массу около 2 кг и длину тела около 50 см. Новорожденный зрячий покрыт короткой черной шерсткой, их и называют черными или черненькими. Первое время они совершенно беспомощны, держатся около матерей и плавать не умеют. Но вскоре они отползают в сторону от гаремов в безопасные места, где скапливаются группами. В таких "детских садах" они защищены от пагубного действия морского прибоя и от ажиотажа, царящего в гаремах среди самцов. Матки регулярно подходят к "детским садам" для молочного кормления новорожденных.
Замечено, что каждая самка кормит только своего детеныша, которого узнает, видимо, по тонким оттенкам его запахов и, быть может, по неуловимым для человеческого взгляда особенностям внешнего вида.
В месячном возрасте черные котики вовсе отделяются от обитателей гаремов и собираются группами по нескольку сотен голов. В это время они имеют длину тела около 60-75 см и массу около 7 кг. Несколько позже они приобретают способность плавать и держатся в бухточках между камнями. В августе (в возрасте около 2,5 месяца) бывает первая линька и взамен черного меха котики покрываются серой шерсткой. Линька заканчивается в середине сентября. К этому времени длина тела увеличивается до 80-100 см. Серые котики больше держатся на воде, а не на берегу. Молочное кормление продолжается до поздней осени.
Максимальная продолжительность жизни превышает 20 лет. На островах Прибылова наблюдали меченых секачей в возрасте 20 лет, а самок - в возрасте 22 лет (беременных). Однако большинство особей доживает до меньшего возраста, примерно до 15-18 лет.
Взрослые котики кормятся рыбой (треска, минтай, бычки, камбала, зубатка, редко лососевые и др.), головоногими моллюсками, реже - ракообразными. Наибольшая масса пищи, обнаруженная у одного из секачей, добытого в море, достигала 17 кг (или 7% от массы тела), а у самки - 4 кг (9% от массы тела). Средняя масса пищи около 5 кг, т. е. 6-7% от массы животного. В желудках котиков часто обнаруживается галька, которая, видимо, служит для механического перетирания пищи.
Линька взрослых бывает с конца июля до ноября; более интенсивно она протекает в сентябре-октябре. Процесс линьки идет медленно, и даже в разгар ее тело котиков покрыто густым мехом, непроницаемым для воды.
Лежбищный период длится до осени. В начале сентября на Командорах наблюдаются большие стада зверей (по 200-300), которые держатся в прибрежных районах моря. Уже в октябре начинается массовый отход на юг. Лишь отдельные секачи остаются и позже. С острова Тюленьего миграция на зимовку бывает в конце октября - начале ноября. Все время до следующего лежбищного периода котики проводят в море, усиленно питаясь.
До недавнего времени считалось, что котики, имеющие лежбища на островах Командорских, Прибылова и Тюленьем, образуют изолированные, несмешивающиеся стада. Массовое мечение не подтвердило полную правильность этого представления. Известен ряд случаев перехода котиков из одного стада в другое.
Котики заражаются гельминтами из трематод (например, Phocitrema furiformis), цестод (Adencephalus septentrionalis, Diphognoporus sp.), нематод (Uncinaria lucasi, Porrocoecum decipiens), клещами (Helarchnae), паразитирующими в трахее и бронхах. Некоторый урон приносят косатки и акулы. Известно много сведений об истреблении котиков особенно молодыми косатками.
Промысел котиков производится только в лежбищный период их жизни, и добыча в море запрещена на основе международных соглашений.
Южный морской котик (Arctocephalus australis) в морях южного полушария ранее был распространен весьма широко. Он водился у побережья Южной Америки, от Чили и Ла-Платы до южной оконечности материка, у юго-западного побережья Африки и берегов Австралии, Тасмании, Новой Зеландии, близ островов Южная Георгия, Южные Шетландские, Кергелен и ряда других, исключая Антарктику. Полагают, что лучше всего котики сохранились на Фолклендских островах, на острове Новая Георгия и у побережья Патагонии. Нет единого взгляда и на систематику южных котиков. Многие исследователи считают, что есть несколько видов южных котиков, например новозеландский (Ar. forsteri), южноавстралийский (Ar. doriferus), тасманийский (Ar. tasmanicus) и др.
По образу жизни южные котики во многом похожи на своих северных собратьев. У них также есть два биологических периода: зимний - морской и летний - береговой, в течение которого котики образуют лежбища, где рождают детенышей, спариваются и линяют. На лежбищах формируются гаремы. Это бывает весной, начиная с ноября. Гаремы существуют около двух месяцев.
Красочное описание поведения патагонских котиков на лежбище дает известный английский зоолог и писатель Дж. Даррелл:
"Колония, насчитывавшая до семисот животных, растянулась полосой вдоль пляжа. Ширина полосы была в десять - двенадцать животных. Тесно сбившись, они ворочались, передвигались, золотисто блестя на солнце, и все вместе напоминали беспокойный рой пчел".
"Первыми обращали на себя внимание взрослые самцы, потому что они были очень массивными. Я никогда не видел животных, у которых был бы такой гордый, такой необычный вид. Они сидели, задрав морды вверх, запрокинув косматые шеи, так что жир на загривке собирался в могучие складки".
"Когда самцы прохаживались, их громадные лягушачьи лапы торчали в разные стороны, а жирные тела колебались в ритме румбы. Это было невероятно трогательно и смешно. Цвета они были от шоколадного до темно-желтого, переходящего в красновато-коричневый на косматых плечах и шее. Громадные и неуклюжие самцы были похожи на бочки, и от этого еще прекраснее и соблазнительнее выглядели рядом с ними их жены, одетые в серебристые и золотистые шубки. Грациозные, изящные, прелестные, кокетливые, с точеными острыми мордочками и большими нежными глазами, они были воплощением женственности".
"Котики имели в своем распоряжении пляж длиной миль в шесть, однако предпочитали лежать, сбившись в тесную кучу, занимая не более четверти мили пляжа. По-моему, если бы они хоть немного рассредоточились, то все неурядицы в колонии сократились бы по крайней мере наполовину, потому что в этой теснотище самцы постоянно нервничали из-за своих жен, и в колонии то и дело возникали драки. Надо сказать, что чаще всего повинны в этом бывали самки. Как только им начинало казаться, что муж не наблюдает за ними, они, грациозно извиваясь, отползали к соседней группе и усаживались там, томно поглядывая на чужого самца".
"И начинался бой. Чаще всего эти драки были чисто символическими, и после нескольких наскоков, сопровождавшихся разеванием пасти и ревом, самолюбие самцов удовлетворялось. Но иногда оба самца приходили в бешенство, и тогда происходило нечто невероятное и страшное - два массивных и на вид отечных существа превращались в быстрых, ловких и беспощадных бойцов. Галька разлеталась во все стороны, два громадных зверя рвали друг другу могучие шеи, и кровь хлестала струей на восхищенных жен и детей".
Большинство стад южного котика не совершает столь длинных сезонных кочевок, как северные котики. После окончания лежбищного периода они уходят в море, но держатся не очень далеко от побережий. Так, например, ведут себя новозеландский и капский котики.
Южного котика добывали издавна. В настоящее время добыча южного котика запрещена, и надо надеяться, что стада этого ценного животного будут в той или иной мере восстановлены.
Калифорнийский морской лев (Zalophus californianus) общим обликом несколько похож на котика, но незначительно крупнее (рис. 210). Длина тела взрослых самцов 215-230 см, изредка до 240 см, у половозрелых самок длина тела обычно 150-175 см, редко до 185 см. Окраска меха однотонная коричневато-бурая, иногда с желтоватым оттенком. Когда звери находятся в воде или вскоре после выхода из нее, мокрая шкура кажется почти черной. Шерстный покров существенно иной, чем у котиков, из низких жестких остевых волос, подшерсток же, столь характерный для котиков, у морских львов отсутствует. Ласты опушены снаружи примерно наполовину, а по внутренней стороне опушения нет.

Рис. 210. Калифорнийские морские львы (Zalophus californianus)
Калифорнийский морской лев распространен только в Тихом океане, в его экваториальных широтах и несколько севернее их (на север примерно до 50° с. ш.). Область распространения этого вида представлена тремя далеко отстоящими друг от друга участками. Первый район обитания - это воды, омывающие Галапагосские острова, второй - побережье Северной Америки от Калифорнийского залива на юге и на север примерно до 40° с. ш., третий район обитания - юго-восточная часть Японского моря и прибрежные части открытого Тихого океана к востоку от Японских островов.
Размножаются, подобно котикам, на суше, но выбирают чаще не отлогие участки побережья (пляжи), а скалистые острова, на камнях которых и образуют залежки. Как и другие ушатые тюлени, морские львы полигамы, и на лежбищах формируются гаремы: около одного самца концентрируется примерно 15-20 самок. Неполовозрелые животные лежат отдельно. Гаремный период длится примерно 3-4 месяца.
На юге Калифорнийского побережья он приходится на май - август, а в Японском море - на июнь - октябрь. Роды соответственно бывают в мае - июне.
Самки рождают одного детеныша вскоре после выхода на лежбище и вслед за этим вновь оплодотворяются. Таким образом, и у этого вида беременность равна примерно году, но значительную часть этого времени оплодотворенное яйцо не развивается (латентная фаза беременности). Новорожденный имеет золотисто-бурую окраску.
После окончания лежбищного периода, в течение которого происходит и линька зверей, они покидают берега и всю остальную часть года проводят в море. В отличие от котиков дальних миграций не совершают и зимой держатся в 5-20 км от берегов. Кормятся преимущественно головоногими моллюсками и рыбой. Считают, что местами они вредят рыболовству не только тем, что уничтожают рыбу (например, сельдь), но и порчей сетных орудий лова.
В настоящее время численность морских львов повсеместно невелика, и в силу этого они не имеют большого промыслового значения.
Южный морской лев (Otaria byronia) систематически близок к только что рассмотренному виду. Кроме особенностей строения скелета, внешне он характеризуется крупными размерами, длина тела самцов-производителей достигает почти 3 м. У взрослых самцов на шее и в области передней части груди волосы удлинены и образуют подобие гривы. Волосяной покров на других частях тела редкий, жесткий, без пуховых волос. Подкожный слой жира значителен, он и выполняет функцию термоизоляции. Окраска в общем буро-серая, более темная на брюхе. Самки темнее самцов, серовато-черные, почти вдвое меньше самцов. Новорожденные имеют более густой коричневато-бурый мех.
Южный морской лев распространен в прибрежных водах Южной Америки, у Огненной Земли, Фолклендских островов, у острова Южная Георгия.
Биологически весьма близок к калифорнийскому морскому льву и вообще к другим ушатым тюленям. Весной (в октябре) у островов или побережья материка появляются взрослые самцы, которые вскоре выходят на берег и занимают определенные участки - места будущих гаремов. Самки подходят к лежбищам в ноябре, и к декабрю большинство гаремов оказываются уже сформированными. Величина гарема невелика и обычно не превышает десятка самок на одного самца.
Рождение молодых бывает примерно в декабре - январе. Первые дни детеныши держатся около самок, которые не сходят в воду. Но после покрытия самки регулярно бывают в море, где кормятся, а новорожденные обычно покидают места залегания родителей и концентрируются на отстоящих недалеко обособленных участках. Замечено, что каждая самка кормит только своего детеныша, которого узнает, видимо, по запаху и голосу. До месячного возраста детеныши плавать не умеют, хотя и делают попытки барахтаться в прибрежных лужах. Позже они сходят в море, где самки учат их плавать. Молочное кормление у южного морского льва длительное, как полагают, не менее 5 месяцев.
В начале февраля гаремы распадаются, и звери переходят к морскому образу жизни. Больших миграций у этого вида не бывает. Характерно, что в апреле лежбища заполняются вновь. Звери выходят на них для линьки.
Промысловое значение невелико, что связано с малоценностью продукции, могущей быть полученной при добыче морских львов.
Сивуч (Eumetopias jubatus) - наиболее крупный вид семейства ушатых тюленей. Длина тела половозрелых самцов 300-325 см, максимальная до 340 см (а при измерении по изгибам спины до 365 см). Самки, как и у всех видов этого семейства, значительно, по крайней мере на 1 м, меньше, чем самцы. Масса взрослого самца около 1000-1120 кг, самок - около 300-350 кг (табл. 38).

Таблица 38. Ластоногие. Сивуч (Eumetopias jubatus)
В отличие от котиков у сивучей ласты оголены не полностью и лишены волос лишь в концевой части. Мех редкий, короткий и жесткий, без подпуши. Окраска меха у взрослых на спине золотисто-соломенная, на брюхе - рыжевато-бурая. Самки несколько темнее самцов. Новорожденные темно-бурого цвета, постепенно светлеющего с возрастом.
Сивучи распространены в северной части Тихого океана, от Берингова пролива на севере на юг до Корейского полуострова по западному побережью и до Мексики по восточному. В деталях распространение носит весьма прерывистый характер и приурочено к прибрежным водам. Сивучи встречаются у юго-западного побережья Чукотского полуострова, у восточного побережья Камчатки, у Командорских островов, вдоль островов Курильской гряды на юг до острова Хоккайдо и северного побережья острова Хонсю. Бывают местами в Охот-ском море, за исключением его западной части.
Распространение сивучей меняется по сезонам. Летом они бывают на лежбищах севернее, чем зимой в морях. Наиболее крупные скопления этих зверей в водах СССР наблюдаются на острове Ионы (Охотское море), на Курильских островах, на островах Ямского архипелага, по восточному побережью Камчатки.
Как и для других ушатых тюленей, для сивучей характерны береговые лежбища, которые в северных ледовитых районах обитания образуются в период размножения, а в южных наблюдаются в различные сезоны года. Сивучи определенно избегают льдов, и лишь изредка приходится наблюдать одиночных зверей на весенних льдах. Полагают, что это те звери, которые при миграциях к местам размножения встретили льды и залегли на них для отдыха.
Лежбища располагаются на скалистых островах, на отдельных камнях рифов, реже - на песчаных и галечных отмелях. Бывают случаи залегания сивучей на скалах, возвышающихся над водой на высоту 20-25 м. С такой большой высоты сивуч бросается в воду без видимого напряжения. Необходимо, чтобы глубина у берега была большой, позволяющей прыгнувшему зверю уйти достаточно глубоко в воду. Для образования щенных залежек важно наличие на камнях хотя бы небольшой площадки, на которой могут расположиться самки с новорожденными.
Щенные залежки образуются летом: на Курильских островах - в середине мая, на островах Охотского моря - в конце мая - начале июня, на Камчатке в середине - конце июня. Сивучи полигамы, и на лежбищах формируются гаремы из 5-20 самок. Характер размножения в основе таков же, как и у других видов семейства. Самки рождают одного детеныша в первые дни выхода на лежбище; спаривание бывает через несколько дней после родов; беременность длится, таким образом, почти год, но есть латентная фаза развития зародыша (см. выше). Новорожденные крупных размеров и составляют 8-9% массы матери, а по длине примерно половину матери (100-120 см). На лежбищах всегда царит оживление и много шума. Самцы ревут низким басом, крик самок несколько похож на мычание коровы, крик детеныша - на блеяние овцы.
Летние береговые лежбища существуют до первых морозов, после чего сивучи уходят в море. Однако в районах, где зимой нет постоянных льдов, залежки образуются и в это время. Таковы, например, зимние залежки взрослых самцов на Командорских островах; на Алеутских островах зимой залегают звери разного возраста и пола. Далеких миграций сивучи не совершают, особенно обитающие в южных частях ареала.
Питаются разнообразным кормом: рыбой (треска, минтай, навага, сайка, камбала, корюшка), головоногими моллюсками (кальмары, осьминоги, каракатицы), редко молодью тюленей. В желудках сивучей обнаруживается галька (до 15 кг), которая, видимо, заглатывается частью случайно, частью преднамеренно: она улучшает механическое измельчение пищи в желудке.
В настоящее время сивучей добывают очень ограниченно. Товарного промысла этих зверей в нашей стране нет.
Семейство Моржи (Odobenidae)
Моржи - самые крупные ластоногие, обитающие в северном полушарии. Как и у ушатых тюленей, у них задние ласты сгибаются в пяточном сочленении и служат для опоры при передвижении по твердому субстрату (земля, лед). Однако, в отличие от ушатых тюленей, наружных ушных раковин у моржей нет. Особенно характерны у моржей клыки верхней челюсти. При закрытом рте они выступают из расщепа рта, далеко спускаясь вниз. Передние ласты обрамлены фестончато вырезанной кожной оторочкой, лишенной волос; нет волос и на нижней части внутренней стороны кисти. На задних ластах вся ступня голая. На обеих парах ластов есть когти, но развиты они слабо.
Зубная формула индивидуально несколько варьирует, но чаще такая:
В настоящее время существует один вид.
Морж (Odobenus rosmarus). Длина тела взрослых самцов (по поверхности) 300-410 см (в среднем 345 см), масса около 1,5 т (в редких случаях до 1,8 т). Размеры самок заметно меньше: длина тела у них от 265 до 335 см (в среднем около 290 см), масса обычно около 700-800 кг (очень редко до 1,1 т). Кроме размеров, самцы-производители отличаются от самок более крупными и толстыми клыками и тем, что шея, грудь и плечи у них в сплошных шишкообразных, бородавчатых утолщениях, что, вероятно, можно рассматривать как вторичный половой признак. У взрослых верхние клыки выступают над десной на 40-50 см (до 60 см). Клыками моржи выкапывают из грунта пищу (моллюсков), защищаются от врагов, а также пользуются при вылезании на льды и сушу. Голова у моржей относительно маленькая с высоко расположенными и широко расставленными глазами. Морда короткая и очень широкая. На верхней губе расположены 13-14 рядов толстых, жестких вибрисс; длина их равна 10-12 см, а толщина - 1,5-2 мм. Вибриссами моржи, видимо, нащупывают кормовые объекты (табл. 38).

Таблица 38. Ластоногие. Моржи (Odobenus rosmarus) на льду
Кожа очень толстая: 3-4 см, а местами (например, на груди) и того более. Волосяной покров у взрослых очень редкий; на значительных участках кожи его нет совсем. Цвет волос рыжеватый или соломенно-сероватый. Только у очень молодых зверей волосяной покров относительно густой, темно-коричневого цвета. С возрастом окраска светлеет. Изменение размеров (длина тела) с возрастом идет так: годовики - 175-220 см, двухлетки - 210-260 см, трехлетки - 260-280 см.
Подкожный жировой слой развит относительно слабо. Толщина его 5-10 см. С одного зверя получают 150-250 кг сала.
Для моржей характерно наличие большого мешковидного выпячивания глотки, расположенного под кожей. Оно может быть заполнено воздухом и этим служить для уменьшения плотности тела, что важно при отдыхе, особенно при сне на воде.
Моржи распространены кругополярно в мелководных окраинных морях Северного Ледовитого океана и соседних участках Атлантики и Берингова моря. В глубоководных районах Центральной Арктики эти звери встречаются лишь изредка.
Отдельные особи проникают на север, во всяком случае, до 79° с. ш. К настоящему времени численность и общая широта распространения заметно сократились и моржи встречаются в отдельных, часто изолированных районах, далеко отстоящих друг от друга. В Западной Арктике малочисленны. Небольшое число моржей встречается в районе архипелага Земля Франца-Иосифа, в области канинско-колгуевского мелководья, местами у Новой Земли, в юго-западной части Карского моря (у западного побережья полуострова Ямал). Также редки моржи у берегов Западного Таймыра и островов Северной Земли. Далее к востоку моржи лучше сохранились у восточного побережья Таймыра, близ устья Лены (например, на острове Куба). В Восточно-Сибирском море они обычны в районе острова Врангеля, более многочисленны в Чукотском море, в рахшне Берингова пролива и северных частях одноименного моря.
Распространение несколько меняется по сезонам, хотя моржам не свойственны столь далекие миграции, как некоторым другим ластоногим. Зимой моржи живут несколько южнее, чем летом, и дальше от берегов. Весной по мере разрушения прибрежных льдов они приближаются к берегам и держатся на плавучих льдах. Так бывает и летом. Численность залегающих на льдах зверей весьма изменчива. Взрослые самцы обычно лежат группами в 10-20 зверей и больше. Самки часто образуют более крупные залежки - до 100-200 особей, но бывают и мелкие скопления - по 10-15 зверей. Для залегания моржи обычно выбирают крупные, многолетние льды. Плотность залегания их на льдах велика, так как звери стремятся к концентрации, и бывает, что они перегружают один конец льдины, в силу этого опускающийся частично в воду, хотя другая часть льдины совершенно свободна.
Стадность у моржей проявляется и в иных формах. Потревоженные чем- либо, они уходят в воду почти одновременно и держатся там очень скученно.
В конце лета часть половозрелых самцов выходит на берег, где на галечных отмелях образует залежки. В настоящее время береговых лежбищ осталось мало, они сохранились преимущественно у Чукотского полуострова, на острове Врангеля, в дельте реки Лены. Численность залегающих зверей также сократилась. На лежбищах бывают десятки, сотни и лишь на немногих лежбищах тысячи зверей.
Соблюдая осторожность и сообразуясь с направлением ветра, можно подойти к лежбищу на 1-2 м. Моржи видят плохо, но обоняние их остро. Вот как описывают В. М. Белькович и А. В. Яблоков (1960) одно из крупнейших в мире лежбищ, расположенное на косе Руддера, на севере Анадырского залива:
"Приближаясь к лежбищу, уже за несколько километров (в хорошую погоду) можно увидеть шевелящуюся массу розовато-коричневых туш... Далеко вокруг разносится неумолчный шум, а от постоянного перемещения зверей в воздухе стоит мелкая пыль. Днем и ночью на лежбище держатся тысячи моржей. По самым приблизительным подсчетам, здесь их не меньше 3000-4000. На расстоянии 50, а то и 100 м от воды весь берег занят лежащими вплотную животными... В море около лежбища на расстоянии нескольких сотен метров также постоянно держится много моржей. Одни из них вылезают, другие, наоборот, уходят в море, третьи сидят неподвижно в прибойной полосе несколько часов подряд, и каждая набегающая волна окатывает их с головой пеной и брызгами.
Вот быстрыми и как-то неожиданно легкими и проворными движениями из воды вылезает большой морж. Не находя свободного места, он взбирается на спины лежащих перед ним животных, расталкивая их, нанося направо и налево удары клыками, пытается пробиться к краю лежки, подальше отводы, где звери лежат не так плотно. Потревоженные им звери обычно отвечают незлобными ударами, но иногда возникают и ожесточенные драки.
Спят моржи в самых разнообразных позах: на спине, на боку или на брюхе, погрузив клыки почти до основания в гальку или песок".
Моржи питаются на мелководьях (до глубины 40-50 м), добывая преимущественно донных двустворчатых и брюхоногих моллюсков, а также червей и ракообразных. Очень редко поедают они рыбу. В желудках моржей лишь иногда находили остатки птиц и лоскуты тюленьей шкуры с салом.
Размножаются очень медленно. Половозрелыми становятся не ранее чем в пятилетнем возрасте. Большинство самок рождают не ежегодно. Считают, что у тихоокеанского моржа ежегодно плодится около 5% самок. Примерно такой же процент самок рождает один раз в два года. Большинство же плодоносит один раз в три или даже четыре года.
Двойни у моржей очень редки. Молочное кормление длится не менее года, так как питание донными организмами возможно лишь при наличии достаточно длинных клыков, которые развиваются только на втором году жизни.
Спаривание бывает в апреле - мае. Гаремов, как у ушатых тюленей, не образуют, но драки между самцами - явление обычное. Беременность длится 330-370 дней, и рождение детенышей происходит примерно в те же сроки, что и спаривание. Детеныши держатся с матерями около двух лет. Хорошо известна большая привязанность самки к детенышу. Указывают, что при любой опасности она не уходит со льдины, если детеныш не может сойти в воду. При приближении людей она помогает ему приблизиться к краю льдины, осторожно подталкивая к воде. Детеныш, спасаясь от опасности или устав плавать, влезает на спину матери.
Хозяйственная ценность моржа весьма значительна. В СССР добыча моржа разрешена только коренному населению Чукотского национального округа и северных районов Якутской АССР. Мясо и жир используются в качестве пищевых продуктов, а шкура идет на различные поделки, в частности для изготовления обшивки вельботов.
Семейство Настоящие тюлени (Phocidae)
Виды, принадлежащие к этому семейству, имеют весьма разнообразные размеры тела: от 1,2 до 6,0 м. В отличие от видов двух предшествующих семейств у настоящих тюленей задние ласты не сгибаются в пяточном сочленении и не могут служить опорой при передвижении по суше или льду; они всегда вытянуты назад и являются основным органом движения при плавании. Обе пары ластов покрыты волосами на всем их протяжении и не имеют кожно-хрящевой оторочки. Когти хорошо развиты и расположены у края ласта. Наружные ушные раковины у всех видов отсутствуют. Шея короткая, малоподвижная. Голова с заметно суженной мордой. На верхней губе 6-10 рядов вибрисс, значительно менее жестких, чем у моржей. Волосяной покров взрослых без явственно выраженной подпуши. У новорожденных ряда видов мех существенно отличен от такового взрослых: он длинный, густой и относительно мягкий. Продолжительность ношения такого младенческого наряда не более трех недель, у некоторых и того менее. Окраска меха разнообразная, "часто пятнистая. Зубная формула:
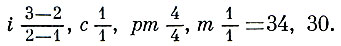
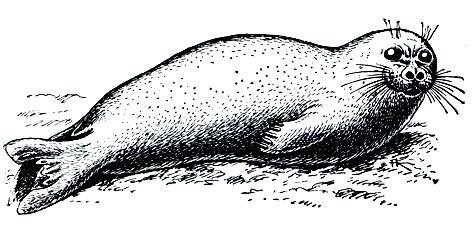
Рис. 211. Морской заяц, или лахтак (Erignathus barbatus)
Большинство видов распространено в холодных и умеренных морях обоих полушарий. Есть в некоторых внутренних водоемах, например в озерах Байкал и Ладожском. В СССР встречаются во всех морях, кроме Аральского и Азовского. В Черном море крайне редки. Размножаются и линяют чаще на льдах, а не на берегах, как ушатые тюлени.
В мировой фауне около 20 видов.
Морской заяц, или лахтак (Erignathus barbatus),- один из наболее крупных видов семейства и самый крупный вид в фауне СССР (рис. 211). Длина тела по прямой от 200 до 225 см, изредка до 240 см. Общая масса взрослых изменчива по сезонам в зависимости от упитанности: в летне-осеннее время обычно до 265 кг; зимой она доходит до 300 кг, а иногда и более того. Размеры самцов и самок практически одинаковы. Окраска волосяного покрова, в общем, однотонная буро-серая, на спине более темная, чем на брюхе. На последнем иногда обнаруживаются слабо выраженные мелкие пятнышки. Волосяной покров сравнительно негустой и грубый. Вибриссы длинные, толстые и гладкие (а не волнистые, как у других тюленей). Самый длинный палец на передних ластах - третий. Зубы сравнительно мелкие, быстро снашивающиеся, и у вполне взрослых зверей они лишь незначительно выступают из десен. Сосков две пары.
Морской заяц распространен кругополярно, преимущественно в окраинных морях Северного Ледовитого океана и северных частей Атлантического и Тихого океанов. В Атлантике к югу встречается до Гудзонова залива и прибрежных вод Лабрадора включительно. В бассейне Тихого океана к югу известен до северной части Татарского пролива. Изредка бывает в центральных частях Северного Ледовитого океана.
Предпочитает мелководные прибрежные районы, особенно такие, где берега изрезаны заливами и бухтами, где есть группы островов. Открытых глубоководных частей моря определенно избегает и в районах, где глубина превышает 50-70 м, обычно не встречается. Такая дислокация вида связана с тем, что лахтак питается преимущественно донными и придонными животными: пластинчатожаберными и брюхоногими моллюсками, креветками, крабами. Местами поедает и сайку (полярную треску).
Сравнительно оседлый вид, у которого бывают лишь местные передвижения. Так, по мере образования прочного сплошного берегового припая большинство лахтаков уходят дальше в море, в зону дрейфующего льда. В морях Тихого океана во второй половине лета и осенью явно тяготеют к прибрежным районам, особенно таким, где есть галечные косы, острова и обнажающиеся во время отлива отмели. В таких местах образуются лежбища, на которых залегают десятки, а местами и сотни тюленей. Лежбища лахтаков принципиально отличны от лежбищ ушатых тюленей тем, что имеют четко выраженный суточный характер. Они образуются каждый день во время отлива и существуют до середины очередного прилива. Береговые лежбища наблюдаются примерно до конца октября - начала ноября, когда по мере появления льдов лахтаки переходят на них и держатся одиночно или группами по 2-3 зверя. Позднее встречаются на льдах чаще, и одновременно можно видеть по нескольку десятков голов. Однако и в это время лахтаки не образуют столь массовых скоплений, как многие другие виды ластоногих.
Некоторые особи и зимой держатся в прибрежной полосе, устраивают во льду лазки, через которые выходят из воды. Иногда лазку заносит толстым слоем снега, и звери сооружают в нем нору.
Весенние скопления на льдах не бывают массовыми, концентрированными, звери залетают распыленно. Залегание на льдах в это время связано с щенкой, линькой и спариванием. Лахтаки выбирают низкие ровные льды и ложатся на краю льдины или около проталины. Лахтак - медлительный, грузный зверь и не может быстро передвигаться по льду.
Щенка бывает в марте - мае. В Охотском море она заканчивается в апреле, в Беринговом море - в мае, в Канадском архипелаге разгар щенки бывает в начале мая. Новорожденный покрыт густыми мягкими, но не длинными волосами темного буро-оливкового цвета, который сохраняется около трех недель. Это вторичный волосяной покров, так как первичный (эмбриональный) коричневато-серого цвета бывает во время утробного развития. Длина тела ново-рожденного около 120 см. Молочное кормление продолжается около 4 недель.
Спаривание бывает на льдах после окончания лактации; таким образом, и у этого вида беременность почти годовая. В начале беременности бывает задержка в развитии и имплантация яйца (латентная фаза) длительностью 2-2,5 месяца. У некоторых самок овуляция бывает после того, как самцы становятся неактивными в половом отношении, и плодоношение бывает не ежегодно. Половой зрелости самки достигают в возрасте 4-6 лет, а самцы - в 5-7 лет.
Промысловое значение морского зайца существенно. Его добывают местное население и специальные зверобойные суда. При промысле используют подкожный жир (40-100 кг с зверя) и шкуру как кожевенное сырье. Местами используют и мясо (преимущественно для кормления пушных зверей).
Обыкновенный тюлень, на Дальнем Востоке - ларга (Phoca vitulina), имеет средний размер. Длина тела географически сильно варьирует: от 140 до 190 см, редко до 210 см. Масса меняется в зависимости от сезона года в пределах 50-150 кг. Самцы лишь несколько крупнее самок. В области Тихого океана живет особенно крупная раса. Окраска также значительно варьирует, чаще ярко-пятнистая: по светлому кремово-серому фону расположены мелкие (2-3 см2) темные пятна неправильной формы. Бывают и значительно более темно окрашенные звери, у которых темные пятна значительно крупнее и чаще расположены (рис. 212).
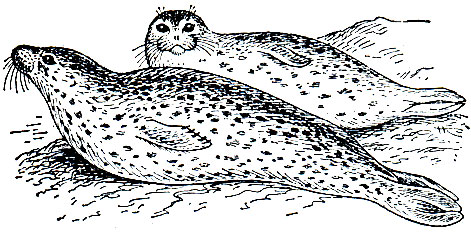
Рис. 212. Обыкновенные тюлени (Phoca vitulina)
В отличие от предыдущего вида у обыкновенного тюленя самыми длинными пальцами на передних ластах является первый и второй. Сосков только одна пара. Вибриссы с волнистыми краями. Зубы крупные, клыки хорошо развиты.
Область распространения состоит из двух разобщенных и далеко отстоящих участков атлантического и тихоокеанского. В первом этот тюлень встречается у южных берегов Гренландии, восточных берегов Северной Америки от Баффинова и Гудзонова заливов на юг по американскому побережью примерно до 35° с. ш. Обычен у Скандинавии, Исландии, на юг до Бискайского залива. Есть в южной части Балтийского моря. По Мурманскому побережью редок. Второй участок ареала приурочен к северной части Тихого океана,, где тюлени обитают в прибрежных районах открытого океана и морях Беринговом, Охотском и Японском на юг до берегов Корейского полуострова включительно, а по восточному побережью до Калифорнии.
У обыкновенного тюленя существенно различаются две географические расы. Звери, населяющие Атлантику, определенно избегают льдов, размножаются и линяют на берегах летом (в конце мая - в июне). Весьма примечательно, что у этой более теплолюбивой расы детеныши сменяют свой первый, эмбриональный меховой покров в утробе матери либо в первые часы после рождения. Эта раса наиболее привязана к прибрежным районам и ведет в общем оседлый образ жизни. Тюлени тихоокеанской расы (особенно те, которые держатся у азиатского побережья) льдов не избегают, а щенка и линька происходят на больших, обычно дрейфующих льдинах. Иные у них и сроки размножения. Деторождение бывает у берегов советского Приморья (в конце февраля - начале марта), в Татарском проливе (в середине марта) и в Беринговом море (в апреле).
Детеныши у дальневосточной ларги рождаются покрытые густым длинным почти чисто-белым мехом, который сохраняется 3-4 недели (стадия белька). После окончания молочного кормления, длящегося около 3-4 недель, бывает спаривание, и, таким образом, беременность длится примерно 11 месяцев. Однако имплантация зародыша наступает только в сентябре, и, следовательно, латентная фаза беременности длится 2-3 месяца. Некоторые особи достигают половой зрелости в три года, но большинство - к четырем годам.
Линька происходит на льдах с середины мая до начала июля. В это время ларга образует залежки в десятки, а иногда и сотни голов. Ларга - очень осторожный зверь с хорошо развитыми слухом и зрением. По льдам она передвигается более легко, чем лахтак, и при опасности ее движения несколько напоминают прыжки.
После исчезновения льдов ларга держится в прибрежных водах, особенно близ устьев рек, куда заплывают для нереста лососевые рыбы, которыми тюлени кормятся. Кроме того, ларга часто поедает сельдь, корюшку, мойву, навагу. Вообще это преимущественно рыбоядный зверь, местами существенно вредящий рыболовству.
В конце лета и осенью обыкновенные тюлени образуют береговые залежки, которые заметны на выступающих из воды рифах, обнажающихся во время отлива отмелях и косах. Как и у лахтака, эти залежки формируются ежедневно, а распадаются во время прилива.
Кольчатая нерпа, на Дальнем Востоке - акиба (Ph. hispida), - один из самых мелких, многочисленных и широко распространенных видов тюленей (рис. 213). Длина тела у него обычно в пределах 110-140 см, наиболее крупные звери достигают длины 150 см. Масса, как и у других тюленей, сильно варьирует по сезонам в связи с накоплением жира. Наибольшей она бывает в осенне-зимнее время, когда большинство зверей (взрослых) достигают 40-80 кг. Самцы у этого вида лишь незначительно крупнее самок. Полового диморфизма в окраске нет. Общий фон окраски несколько изменчив индивидуально от светло-серебристого до темно-серого. По этому фону расположены темные, неправильной формы пятна, окаймленные светлыми кольцами.

Рис. 213. Кольчатая нерпа (Phoca hispida)
Волосяной покров относительно густой и длинный, и шкуры нерпы используются не только как кожевенное сырье, но и для пошива меховых изделий, например курток.
Нерпа распространена в Северном Ледовитом океане, преимущественно в его окраинных морях и в морях северных частей Атлантического и Тихого океанов, там, где хотя бы зимой бывают льды. К югу она встречается до берегов Норвегии, Балтийского моря, по Атлантическому побережью Северной Америки до залива Св. Лаврентия, а по Тихоокеанскому побережью - до полуострова Аляска, по азиатскому побережью - до северной части Татарского пролива. Есть в озерах Ладожском (СССР) и Сайманском (Финляндия).
Хотя нерпа не связана, подобно морскому зайцу, с мелководьями, она большую часть года явно тяготеет к прибрежным водам, особенно к таким, где берега изрезаны заливами, где есть острова. Больших миграций не совершает, однако в зависимости от времени года ее концентрации наблюдаются в несколько отличных местах. Летом держится преимущественно в прибрежных водах и местами образует на камнях или галечных косах незначительные залежки. Осенью по мере замерзания моря большая часть зверей уходит из прибрежной зоны в глубь моря и держится на дрейфующих льдах. Меньшее их число остается на зиму у берегов и держится в заливах и бухтах. В этом случае еще в начале замерзания моря нерпа проделывает в молодом льду отверстия - лазки, через которые выходит из воды. Бывают отверстия и меньшего размера, используемые лишь для того, чтобы дышать через них. Нередко отверстие лазки заносится толстым слоем снега, в котором нерпа устраивает нору без выходного отверстия наружу. В таком удобном месте она отдыхает, будучи невидимой для врагов, главным образом белых медведей.
Наибольшие скопления наблюдаются весной на дрейфующих льдах во время щенки, линьки и спаривания. Особо характерно это для морей Дальнего Востока, где за один день плавания во льдах можно наблюдать многие сотни, а иногда и тысячи зверей. Чаще нерпы лежат группами по 10-20 голов, но бывают скопления в сотню и более зверей. Они остаются на льдах до их исчезновения. Нерпы менее осторожны, чем ларги.
Щенка происходит на льдах с конца февраля до начала мая в зависимости от района. Звери, зимовавшие в прибрежной области, чаще щенятся в снеговых норах. Иногда такие норы сооружаются и на дрейфующих льдах. В иных случаях для деторождения выбираются крепкие льдины, имеющие торосы, среди которых и укрывается новорожденный. Детеныш рождается длиной около 50 см и массой около 4,5 кг. Он покрыт густой, длинной (2-2,5 см), мягкой шерстью молочно-белого или слегка сероватого цвета, сохраняющейся около 2-3 недель (белек).
Молочное кормление длится около месяца, и в это время детеныши не сходят в воду, но самка регулярно покидает белька и кормится в море.
Половая зрелость у немногих самок наступает на четвертом году жизни, у большинства - на пятом году, большая часть самцов становятся половозрелыми в возрасте 5-7 лет.
Вскоре после щенки у взрослых наступает линька, длящаяся до конца лета, а иногда и до осени. Линяющие звери особенно неохотно сходят со льдов в море, и добыча их наиболее легка. Однако в это время упитанность зверей наименьшая и добыча их не столь выгодна.
Спаривание бывает после окончания молочного кормления (в июле - августе). При беременности в первые 3-3,5 месяца зародыш не имплантируется и почти не развивается.
Состав пищи весьма разнообразен: различные ракообразные и массовые виды рыб - мойва, навага, корюшка. Существенного вреда для рыбного хозяйства не приносит.
Промысловое значение существенно, особенно в морях Дальнего Востока. Основными продуктами промысла являются жир и шкура, идущая на изготовление кожевенных и меховых изделий. Выход сала с одного зверя от 6 до 20 кг. Наиболее упитанными нерпы бывают к концу зимы, наименее - летом, во время линьки.
Байкальская нерпа (Ph. sibirica) систематически, несомненно, близка к только что рассмотренной кольчатой нерпе, но отличается от нее одноцветной окраской. Верх тела этой нерпы буровато-серый с серебристым оттенком; низ тела несколько светлее. Волосяной покров, как и у обыкновенной нерпы, относительно длинный и густой. Длина тела взрослых от 110 до 150 см, а по некоторым сообщениям - до 160 см. Масса, как и у всех тюленей, сильно варьирует по сезонам: от 60 до 100 кг.
Встречается только в озере Байкал, из которого заходит в реки, например в Ангару и Селенгу.
Происхождение байкальской нерпы до сего времени не выяснено. Вероятно, она проникла в озеро еще в третичное время через систему существовавших тогда крупных, связанных между собой и с морем внутренних бассейнов, одним из членов и последним звеном развития которой есть Байкал.
Байкальская нерпа распространена в северных частях Байкала более, чем в южных. Особенно заметно это зимой и в начале весны. На поверхность льда нерпа зимой не выходит и дышит в продушинах, которые сооружает в молодом, еще тонком льду.
Беременные самки нерпы устраивают во льду лазки и сооружают в толще снега цору, в которой и щенятся. Щенные самки сосредоточиваются преимущественно близ восточных берегов.
Щенка бывает в феврале - марте. Детеныш (длиной около 60-70 см и массой 3-3,5 кг) родится одетый бельковым мехом сохраняющимся у него в течение месяца.
Молочное кормление длится более месяца, по некоторым сведениям - до 3 месяцев.
В конце апреля - в мае наблюдается массовый выход зверей всех возрастов на лед, где они образуют залежки. Больших скоплений байкальские нерпы не образуют.
В мае - июне на льду происходит спаривание, а вслед за этим линька. После исчезновения льдов держатся чаще в прибрежной зоне, а местами на камнях и косах образуют малочисленные залежки.
Байкальская нерпа питается почти исключительно непромысловой рыбой (бычками, голомянкой) и рыболовству не вредит.
Таблица 35. Ластоногие: 1 - 4 - группа гренландских тюленей (Pagophoca groenlandica): 1 - самка, 2 - самец, 3 - белек, 4 - серка; 5 - полосатый тюлень (Histriophoca fasciata); 6 - каспийская нерпа (Phoca caspica); 7 - тюлень-монах (Monachus monachus); 8 - морской леопард (Hydrurga leptonyx)

Таблица 35. Ластоногие: 1 - 4 - группа гренландских тюленей (Pagophoca groenlandica): 1 - самка, 2 - самец, 3 - белек, 4 - серка; 5 - полосатый тюлень (Histriophoca fasciata); 6 - каспийская нерпа (Phoca caspica); 7 - тюлень-монах (Monachus monachus); 8 - морской леопард (Hydrurga leptonyx)
Каспийская нерпа (Ph. caspica) по размерам неотличима существенно от других нерп (табл. 35). Длина тела 120-150 см, масса - 40-60 кг, а в период наибольшей упитанности у некоторых оробей может достигать 90 кг. Окрасу взрослых самцов с многочисленными разной величины темными пятнами, разбросанными по сизовато-серому общему фону.
Самки окрашены более тускло, и пятен у них меньше. Менее развита пятнистость у неполовозрелых зверей. Бельки имеют длину тела около 70 см и массу около 4 кг. Они покрыты густым мягким белым волосом, который сохраняется в течение 2-3 недель.
История происхождения каспийской нерпы полностью не выяснена. Наиболее вероятно, что она является потомком местных южных тюленей, населявших в раннетретичное время Сарматско-Понтический бассейн, одним из остатков которого является Каспийское море.
В современное время нерпа распространена по всему Каспийскому морю, но в связи с массовыми сезонными миграциями концентрируется в различных частях моря в зависимости от времени года. Летом основная масса нерпы держится в южной, глубоководной части моря, южнее устья Терека - на западном берегу моря и близ полуострова Мангышлак - на восточном. Большую часть времени они проводят на воде и лишь местами образуют береговые лежбища. В конце августа нерпы начинают мигрировать в северные части моря. Причем большая часть зверей идет вдоль восточного берега моря. Первыми идут половозрелые самки, затем взрослые самцы и последними - неполовозрелые звери. Массовый ход бывает в ноябре - декабре. В октябре - ноябре скопившиеся в северной части моря нерпы образуют большие береговые залежки на песчаных отмелях островов и кос. Они существуют до образования льда.
В январе самки, собравшиеся в стада (косяки), входят во льды, где образуют щенные залежки, которые обычно располагаются в центральных частях ледяных скоплений, на крепких льдах. Наиболее мощные залежки образуются в северо-восточной части моря. Период щенки в разные годы растягивается с конца января по апрель. Новорожденный лежит у лазки прямо на льду. Самки большую часть времени проводят в воде, выходя на лед только для кормления молодого. Молочное кормление длится около 4-5 недель.
Перед концом молочного кормления щенные самки начинают линять, собираясь при этом в большие косяки. В конце марта к линяющим самкам присоединяются самцы. Косяки линяющих зверей увеличиваются. Линька кончается к началу мая, ко времени исчезновения льдов. Не успевшие долинять на льдах нерпы образуют небольшие линные залежки па отмелях и косах.
Спаривание происходит на льду, вскоре после щенки, т. е. с конца февраля, и длится в течение почти всего марта. Лишь как исключение спаривание бывает на берегу. Половой зрелости самцы достигают, по-видимому, на третьем году, самки - на втором.
После окончания линьки происходит обратная массовая миграция тюленей из северных частей моря в южные, где они проводят лето.
Каспийская нерпа питается преимущественно непромысловыми видами рыб (бычками, атериной) и ракообразными. В последние годы в питании большое значение имеет акклиматизированный рачок - креветка леандр. Существенного вреда рыбному хозяйству нерпа не приносит.
Серый, или длинномордый, тюлень, тевяк (Halychoerus grypus) - это сравнительно крупный тюлень: самцы имеют длину тела 165-260 см и массу около 300 кг; сайки несколько мельче, длина тела у них 155-190 см, а масса примерно 150-200 кг. Морда сильно удлиненная, без уступа в области переносицы. Ноздри очень крупные, расположенные на самом конце морды. Общий фон окраски серый, по нему разбросаны темные пятна разной величины и интенсивности (рис. 214).

Рис. 214. Тевяк (Halychoerus grypus)
Область распространения в виде трех разобщенных участков. Один в Северо-Западной Атлантике - у американского побережья, в районе залива Св. Лаврентия и Гренландии; другой приурочен к Северо-Восточной Атлантике, к прибрежным водам Британских островов, Скандинавского полуострова, Мурманского побережья, Шпицбергена; наконец, третий участок - в Балтийском море" включая его заливы.
Этот тюлень ведет в общем оседлый образ жизни, и четко выраженных длительных миграций у него нет. Пища состоит преимущественно из рыбы (трески, камбалы, лосося, сельди), реже - из ракообразных. Условия размножения различны в разных частях ареала. Балтийские тевяки щенятся на льдах чаще в марте. На Мурманском побережье, на Британских островах деторождение бывает осенью - в начале зимы, обычно в ноябре, на берегу У атлантической популяции береговые залежки бывают весьма многочисленные (до 1000 голов). В это время наблюдается образование небольших гаремов. Спаривание происходит через две недели после щенения. Новорожденные одеты белым, густым и длинным мехом, сохраняющимся около недели.
Существенного промыслового значения не имеет. Местами вредит рыболовству (поедает рыбу и портит сети).
Гренландский тюлень, или лысун (Pagophoca groenlandica), средней величины, с весьма своеобразной окраской (табл. 35). Длина тела взрослых чаще 180-185 см, крайние вариации 150-193 см. Масса Туши в начале зимы достигает 160 кг. Самцы и самки по размерам практически неотличимы. У старых самцов общий фон окраски желтовато-белый, по бокам тела располагаются два симметричных полулунной формы черных поля; черный также и верх головы. У самцов меньшего возраста, а также у старых самок поля не черные, а темно-бурые. Молодые половозрелые самки окрашены в светло-серый общий тон, по которому разбросаны темные пятна неправильной формы.
Новорожденный имеет длину 85-95 см, покрыт белым густым и длинным мехом. Через 2-3 недели белый эмбриональный волос выпадает и заменяется коротким серым мехом. Линяющего детеныша называют хохлушей. По окончаний линьки при длине тела 110-125 см тюлененок называется серкой. В возрасте года и двух звери имеют пепельно-серую окраску с темными пятнами.
Гренландский тюлень населяет северные широты Атлантики и частично Северного Ледовитого океана от восточных окраин Канадского архипелага и полуострова Лабрадор на западе до западных районов Карского моря, а в некоторые годы-до западных частей моря Лаптевых на востоке. Северным пределом распространения служит граница тяжелых арктических паковых льдов. На юг этот вид распространен до Ньюфаундленской банки у американского побережья, южной оконечности Гренландии, северных берегов Исландии, южнее Шпицбергена, до Мурманского побережья, Белого моря, Чешской губы, Карского моря близ Новой Земли.
В пределах этой обширной области тюлени встречаются не сплошь, а в определенных районах, нахождение которых меняется по сезонам. Особенно заметно это в зимнее время - до начала весны, когда ясно вырисовываются три хорошо обособленных и, по-видимому, несмешивающихся стада, собирающихся для размножения и линьки в трех ограниченных по площади и далеко отстоящих райо-нах. Такими стадами являются: 1) беломорское, размножающееся преимущественно в Воронке и Горле Белого моря; 2) янмайенское, образующее детные залежки в районе острова Ян-Майен; 3) ньюфаундлендское, залежки которого приурочены к району острова Ньюфаундленд. На лето каждое стадо мигрирует на север к кромке пака в свой район арктических широт.
В территориальных водах СССР находятся залежки беломорского стада, которое летует к востоку от Шпицбергена, иногда проникая до западных частей моря Лаптевых. Осенью тюлени массами начинают двигаться на юг к местам щенки и линьки. В декабре они появляются на востоке Мурманского побережья и в Горле Белого моря. Первыми подходят половозрелые самки, самцы же и неполовозрелые запаздывают. В начале февраля начинаю! формироваться щенные залежки. Самки выбирают обширные и крепкие льдины, покрытые снегом. Торосистых и битых льдов самки избегают и щенятся на них лишь при отсутствии подходящих льдин. Щенка начинается с февраля. Самка рождает одного (очень редко двух) бельков длиной около 85 см и массой около 8 кг, Первые 7-10 дней она часто лежит на льдине вместе с детенышем, позже выходит на лед только для молочного кормления молодого, а остальное время проводит в воде. Лактация длится около 4 недель.
Ньюфаундлендское стадо летует рассредоточенно от Лабрадора до Гренландии. Арктику тюлени этого стада покидают в октябре - ноябре и двигаются вдоль Лабрадора на юг. В середине февраля беременные самки появляются на льдах залива Св. Лаврентия и у Ньюфаундленда и образуют щенные залежки. Общая площадь их примерно 260 км2, а плотность залегания маток в прошлом была около 2-3 тысяч на 1 км2. Щенка бывает в конце февраля -начале марта. Примерно в эти же сроки размножаются и тюлени ян-майенского стада.
Покрытие самок происходит в конце молочного кормления, в конце марта - первой половине апреля. Беременность длится 11 месяцев, но в развитии зародыша бывает латентная фаза, приходящаяся на первые 2-2,5 месяца.
Во второй половине марта половозрелые самцы (лысуны) собираются в большие стада в Мезенском заливе, в Горле Белого моря и образуют на льдах линные залежки. Несколько позже к залежкам лысунов присоединяются самки (утельги) и неполовозрелые звери (серуны). Нередко в одной залежке бывает по нескольку тысяч голов. Залежки, образующиеся в глубине Горла и в Мезенском заливе, с течением времени постепенно выносятся к северу и к апрелю - началу мая они оказываются у кромки дрейфующих льдов. Кроме того, в начале мая наблюдается активное кочевание уже вылинявших тюленей из Белого моря к северу. Выйдя из Белого моря, тюлени задерживаются на некоторое время у восточных берегов Мурманского побережья, где усиленно питаются, а затем идут к местам летовок.
В районе Ян-Майена линные залежки наблюдаются в апреле - начале мая. У Ньюфаундленда лысуны начинают линять в начале апреля, самки - во второй половине этого месяца.
Половой зрелости самки достигают в возрасте 4-8 лет, самцы - около 8-9 лет. Потенциальная продолжительность жизни около 30 лет.
В период летовки пища тюленей состоит из ракообразных, моллюсков, рыбы.
Осенью и в начале зимы в рационе, наоборот, преобладает рыба (сайка, мойва, сельдь, морской окунь), ракообразных гюлени поедают реже. Во время линьки не кормятся и сильно худеют. Наиболее упитанными бывают осенью и в начале зимы.
Промысловое значение гренландского гюленя очень велико,, и история его добывания насчитывает несколько веков.
Человек использует сало и кожу тюленей, а при добыче бельков - мех, хорошо поддающийся имитации под мех бобра, выдры, норки. В Белом море судовой промысел тюленей запрещен.
Полосатый тюлень, или крылатка (Histriophoca fasciata), имеет среднюю величину и очень своеобразную окраску. У взрослых самцов общий фон темно-коричневый, часто почти черный. На этом фоне располагаются белые полосы шириной 10-12 см. Одна полоса кольцом перепоясывает тело, другая кольцеобразно охватывает область крестца, наконец полосы в виде овалов есть и на боках тела, где они окружают основание передних ластов (табл. 35). Самки имеют тот же тип окраски, но общий фон у них светлее, коричневато-бурый, иногда почти серый. Неполовозрелые звери после первой линьки одноцветно серые. Новорожденный одет длинным густым, белым мехом, сохраняющимся около двух недель.
Длина тела у половозрелых 150-190 см (чаще до 180 см). Масса - 70-90 кг. Самцы и самки примерно одинаковых размеров. Детеныш родится с длиной тела 70-80 см.
Распространение этого тюленя недостаточно выяснено. Известно, что весной и в начале лета он залегает на льдах в Охотском и Беринговом морях и в южных районах Чукотского моря. Изредка бывает в это время и на льдах северной части Татарского пролива. Предпочитает открытые участки моря, но при дрейфе льдов может оказаться и в прибрежных районах. Весенне-летнее залегание на льдах связано с щенкой, спариванием и линькой.
После исчезновения льдов крылатка уходит в открытые части морей, но ее точные места нахождения осенью и зимой не установлены.
Залегая на льдах, крылатка выбирает только крепкие и обязательно чистые белые льдины. Не избегает она и льдов с торосами, но таких, где есть ровные площадки, на которых зверь и ложится. Высота льдов существенного значения не имеет. Тюлень этот удивительно ловок и даже на высоко возвышающиеся льдины вскакивает энергичным красивым прыжком. Другая характерная черта поведения - это большое беспокойство. Для того чтобы залечь на льдине, тюлень этот то впрыгивает на нее, то вновь уходит в воду. Выйдя на льдину, он некоторое время быстро передвигается по ней, долго выбирая подходящее место, и только потом засыпает.
Находясь на льду, крылатка малоосторожна, и подкрасться к ней на близкое расстояние легче, чем к многим другим тюленям.
Щенка бывает в марте - апреле. Белек в воду не сходит и при опасности прячется среди торосов. На чистых белых льдах его окраска сливается с общим фоном местности и только большие темные глаза выдают присутствие затаившегося звереныша.
Спаривание бывает на льдах в июне - июле (местами в мае - июне). Половая зрелость наступает ранее, чем у других северных тюленей, уже со второго года жизни, но чаще в 3-4 года.
Линька проходит в мае - июне очень бурно, и вместе со старым волосом лоскутами сходит верхний слой эпидермиса.
Взрослые питаются преимущественно рыбой (минтаем, треской), головоногими моллюсками, реже - ракообразными. Молодые звери, начинающие самостоятельно кормиться, поедают главным образом ракообразных.
Промысловое значение этого тюленя довольно велико. Однако залежки ее распространены весьма спорадически, и отыскание их требует много времени.
Тюлень-монах (Monachus monachus) - довольно крупный зверь, длина тела которого достигает почти 3 м (чаще всего 240-275 см), масса около 300 кг; окраска черновато-коричневая. На нижней поверхности тела располагается ромбовидное вытянутое пятно длиной около 75 см грязно-белого цвета. Тюлень этот редок и встречается спорадически в прибрежных водах Атлантического океана: у африканского побережья в Средиземном море и в немногих районах Черного моря - у берегов Турции и Румынии. В СССР, вероятно, единственное место, где держится небольшое число этих тюленей, находится в прибрежных районах между дельтой Дуная и островом Змеиным. Биология изучена плохо. Известно, что размножаются они на берегу, выбирая для этого уединенные песчаные или галечные косы или участки каменистого берега в заливчиках. Щенка происходит, видимо, в растянутые сроки, с июля по сентябрь. Детеныш рождается не с белой длинной шерстью, как у многих других (северных) тюленей, а с короткой темно-бурой. Около 2 месяцев, в период молочного кормления, он находится на берегу. Взрослые кормятся рыбой и крупными ракообразными. В связи с малочисленностью никакого промыслового значения не имеет. Необходимы мероприятия по охране этого единственного в Черном море вида тюленей (табл. 35).
В районе Вест-Индских островов и Карибского моря распространен другой вид тюленя-монаха - тропический (M. tropicalis); в районе Гавайских островов - гавайский тюлень-монах (M. shauinslandi). Это редкие звери, не имеющие хозяйственного значения.
Хохлач (Cystophora cristata) - сравнительно крупный тюлень с причудливой пестрой окраской. Наиболее крупные самцы достигают в длину почти 3 м (чаще 200-280 см), масса самца около 300 кг. Самки заметно мельче; длиной 170-230 см и массой около 150 кг. Общий тон окраски серый, по которому разбросаны темные буро-коричневые или почти черные пятна крайне разнообразных причудливых очертаний (рис. 215). На спине пятна расположены чаще, чем на брюхе, где они нередко соединяются. Новорожденный не имеет белькового наряда и покрыт коротким жестким мехом. На спинной стороне он серого цвета, на брюхе почти белого. Эмбриональный мех сменяется еще в утробе матери.

Рис. 215. Хохлач (Cystophora cristata)
У самцов на верхней части морды имеется очень своеобразный парный полый кожистый вырост, полость которого сообщается с носовой полостью. При возбуждении эта полость наполняется воздухом, в связи с чем верх головы приобретает весьма причудливые очертания.
Хохлач - арктический вид тюленя, населяющий северные районы Атлантического океана и смежные окраины Северного Ледовитого океана. Он встречается у западного побережья Канадского архипелага (в Баффиновом заливе и Девисовом проливе к югу до района острова Ньюфаундленд), у побережья Гренландии, особенно в Датском проливе, на восток примерно до Шпицбергена. В СССР изредка бывает в северных частях Белого моря.
В отличие от ряда других тюленей хохлач не связан непосредственно с прибрежными водами и придерживается преимущественно районов близ кромки арктических льдов. Питается рыбой (треска, сельдь, морской окунь) и головоногими моллюсками. Подобно гренландскому тюленю в период размножения сосредоточивается в немногих ограниченных по площади областях. Основными из них будут районы близ островов Ньюфаундленд и Ян-Майен, где на льдах образуются щенные залежки. Однако столь же большого, как у гренландского тюленя, сгущения зверя при этом не образуется.
Сроки щенки в указанных двух районах несколько различны. На ньюфаундлендских залежках щенка происходит в конце февраля - начале марта, на ян-майенских залежках - в середине марта. Детеныши у хохлача, не имеющие белькового наряда, кормятся молоком около 2-3 недель. После окончания лактации бывает спаривание. Беременность с латентной фазой, и общая ее длительность около 11 месяцев.
Линные залежки образуются главным образом в Датском проливе (между Гренландией и Исландией) в июне - начале июля. Промысловое значение хохлача очень большое.
Южный морской слон (Mirounga leonina) - один из наиболее крупных тюленей: длина тела самца может достигать 5,5 м (по некоторым данным, и более того), масса его до 2,5 т. Самки заметно мельче, длина тела у них обычно менее 3 м. Как и у хохлача, к которому морской слон систематически близок, у самцов этого вида развит кожистый мешок, расположенный на верхней стороне морды. Когда зверь возбужден, мешок несколько распрямляется и длина его достигает 60-80 см. При спокойном состоянии зверя длина мешка сокращается вдвое. Некоторое сходство этого мешка с хоботом слона, а также крупные размеры зверя и послужили причиной тому, что этого тюленя называют морским слоном (табл. 39).
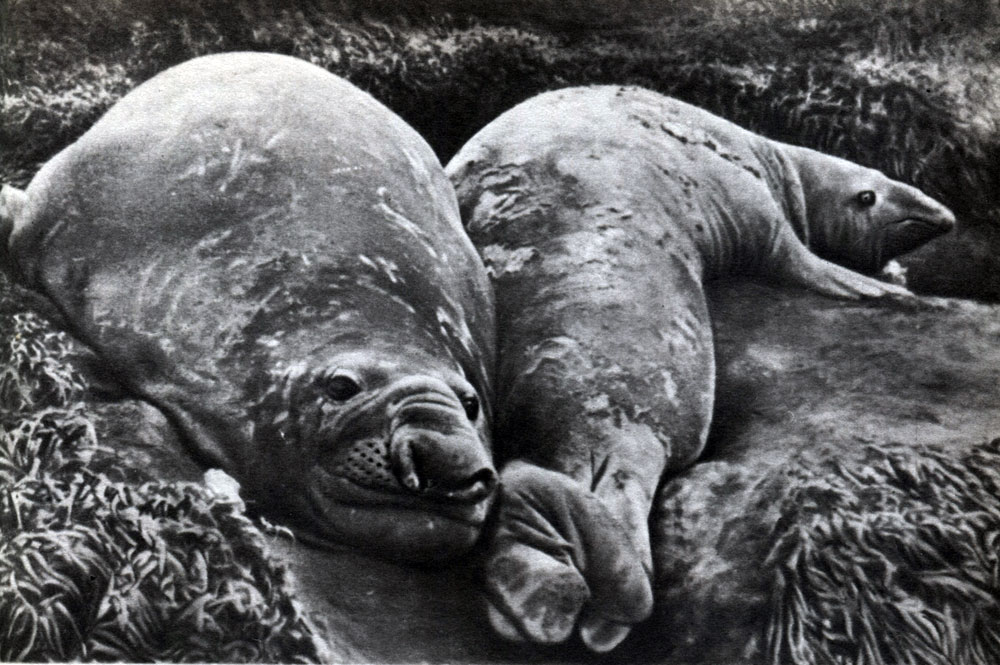
Таблица 39. Ластоногие. Южные морские слоны (Mirounga leonina), самцы
Мех взрослых короткий, жесткий, буро-коричневый. Новорожденные покрыты густым черным мехом, который в возрасте 1-2 месяцев заменяется серебристо- серым. Подкожный жировой слой развит очень сильно. Так, взрослый самец длиной 4,06 м имел массу 1980 кг, причем на долю подкожного сала приходилось 34% от общей массы, на долю мяса - 21%, костей - 15%, шкуры - 6%. Средний выход жира с одного тюленя 420 кг. Столь сильная ожиренность зверя хорошо заметна при его передвижении по суше: туловище тюленя сотрясается подобно студенистой массе.
Этот вид тюленя распространен в южном полушарии, в субантарктических водах. Его лежбища располагаются на Фолклендских, Южных Оркнейских, Южных Шетландских островах, на островах Кергелен, Южная Георгия. Есть лежбища и на побережье Южной Америки (Патагония, Чили, Огненная Земля). Во многих районах численность морских слонов в недалеком прошлом заметно уменьшилась. Прекращение промысла заметно способствовало восстановлению поголовья этого замечательного тюленя, и в настоящее время вновь производится его добыча, но в ограниченных пределах. Наиболее крупные стада на островах Южная Георгия и Кергелен (примерно по 250-260 тысяч голов).
Морские слоны - широко мигрирующие звери. Летом они держатся на береговых лежбищах, где происходит деторождение, спаривание и линька. На зиму большинство уходит на север, в более теплые воды. И только незначительное число остается в районах береговых лежбищ. Пути миграций основной массы зверей и места их зимовок точно не известны. Для выяснения этого в недавнее время широко развернуто мечение преимущественно молодых зверей на лежбищах.
Лежбища слонов располагаются на песчано-галечных пляжах, нередко в бухточках и заливах. Неразмножающиеся звери залегают и на значительном отдалении от моря (на несколько сотен метров) обычно по берегам ручьев. В этом случае они менее прихотливы к грунту и ложатся на участки, прикрытые травой или мхами, иногда несколько заболоченные. О характере формирования лежбищ существуют разные мнения. Одни исследователи полагают, что первыми к берегам подходят половозрелые самцы, а примерно через две недели - плодующие самки, из которых самцы формируют гаремы. Рождение детенышей бывает уже в гаремах.
По другим сведениям, к лежбищам первоначально подходят взрослые самки, которые вылезают на берег и щенятся в отдалении, а несколько позже к ним подходят самцы. Гаремы, согласно этим наблюдениям, формируются лишь после того, как пройдет щенка. Можно предполагать, что в разных районах возможна разная последовательность образования лежбищ и формирования гаремов.
Половозрелые звери подходят к лежбищам весной, в конце августа - начале сентября. Неполовозрелые особи запаздывают примерно на месяц. Замечено, что сроки появления зверей сильно растянуты, и роды наблюдаются с конца августа до начала ноября, но чаще всего с конца сентября до второй декады октября. Родится, как правило, один детеныш длиной 75-80 см и массой 15-20 кг. Спаривание бывает вскоре после родов, беременность длится около 11 месяцев. Молочное кормление продолжается около месяца, после чего детеныши часто покидают семейные лежбища и залегают отдельно от взрослых. После окончания лактации детеныши несколько недель не сходят в воду, ничего не едят и существуют за счет подкожного жира.
Во время образования гаремов между самцами бывают драки. При этом они громко ревут, распрямляют "хобот", которым как бы размахивают, бросаются друг на друга и наносят клыками подчас сильные раны. При этом часто повреждают "хобот". Обычно малоподвижные, как бы флегматичные, самцы во время драки преображаются, проявляя поразительную ловкость и энергичность. Порой они выпрямляются почти во весь рост и, энергично действуя хвостовой частью тела, выделывают удивительные пируэты, временами почти полностью отрываясь от земли. В остальное время слоны на лежбищах большей частью спят, они мало обращают внимания на посторонние звуки, и к ним можно подойти вплотную.
Раньше всех покидает лежбища приплод текущего года. Это бывает в середине лета, когда молодым 2-3 месяца. В ноябре гаремные лежбища постепенно распадаются. Сильно истощенные самки некоторое время откармливаются в море, после чего образуют линные залежки. Примерно в это же время, т. е. в ноябре, у берегов скопляются неполовозрелые слоны, у которых вскоре тоже начинается линька. Лишь часть их лежит на прибрежных отмелях, а большинство отходит на 100-200 м (иногда и более того) в глубь суши и располагается на лугах и торфяниках, нередко сырых. Позже всех, в марте, происходит линька у половозрелых самцов. Закончив линьку, звери всех возрастных групп покидают сушу. Большинство зверей уходит в открытое море, где проводит зиму. В районе лежбищ остаются лишь немногие слоны.
В районе лежбищ слоны кормятся главным образом головоногими моллюсками, реже - рыбой. Характер питания в морской период жизни точно не известен, но полагают, что в это время головоногие моллюски важная составная часть их рациона.
Морские слоны еще несколько столетий назад привлекали внимание промышленников, снаряжавших зверобойные корабли в субантарктические районы. Зверей добывали на многочисленных береговых лежбищах - на островах Южная Георгия, Кергелен, Южные Шетландские и др. Однако в настоящее время промысел морских слонов международным соглашением строго ограничен.
Северный морской слон (Mirounga angustirostris) внешне и по образу жизни весьма близок к своему южному собрату и отличается от него главным образом более крупными размерами. В настоящее время незначительные, но увеличивающиеся по численности стада держатся у острова Гвадалупе и у побережья Калифорнии.
Тюлень Уэдделла (Leptonychotes weddelli) - типичный обитатель антарктических вод. Среди настоящих тюленей Антарктики это один из самых многочисленных видов. Это довольно крупный зверь, длина тела которого достигает 300 см, при этом самцы несколько мельче самок (длина до 260 см). Общая окраска телесно- серовато-коричневая, у многих почти черная с серебристо-серыми, иногда почти белыми овальными пятнами на брюхе и боках. Волосяной покров короткий, жесткий, без подшерстка. Подкожный жировой слой, наоборот, развит очень сильно, толщина его у взрослых достигает 7 см, а общая масса подкожного жира у наиболее сильно ожиревших зверей составляет почти 30% от массы тела. Несомненно, это служит важным приспособлением к обитанию при низких антарктических температурах.
Тюлень Уэдделла (рис. 216) распространен близ антарктического континента и расположенных поблизости островов. Известны лишь немногие случаи встречи этих зверей на субантарктических островах и даже у берегов Австралии и Новой Зеландии. Сравнительно с многими другими видами тюленей, он не совершает больших миграций и держится преимущественно в прибрежных водах,, где летом на льдах или на берегу образует немногочисленные залежки (по 50-200, редко более голов). В конце осени тюлени держатся у кромки льдов и устраивают в молодых льдинах отверстия - продушины, через которые дышат в течение долгой антарктической зимы. Продушины регулярно покрываются льдом, и столь же регулярно тюлени возобновляют их. Эту работу они проделывают зубами, и поэтому у старых животных клыки и резцы бывают сломанные.

Рис. 216. Тюлень Уэдделла (Leptonychotes weddelli)
На поверхность льда тюлени зимой выходят очень редко, что, видимо, связано с низкими температурами воздуха и сильными ветрами.
Размножение бывает весной, в сентябре - октябре, на прибрежных или крупных плавающих льдах, на которых тюлени образуют небольшие скопления. Новорожденные имеют длину тела 120-130 см и массу около 25 кг. Они покрыты густым, мягким и длинным мехом рыжевато-серого цвета с небольшими более темными пятнами. Такой мех сохраняется в течение 1,5 месяца. В воду молодые тюлени сходят, еще не закончив молочное кормление, примерно в возрасте 6 недель.
Спаривание бывает вскоре после окончания периода молочного кормления, беременность длится около 10 месяцев.
Кормятся преимущественно головоногими моллюсками и рыбой. Ныряют при добыче пищи на значительные глубины. С помощью специального прибора, прикрепленного к спине животного, удалось установить погружение самок на 320-395 м, а самцов на 335-350 м.
Тюлени Уэдделла мало боятся человека, и к ним можно подойти вплотную. В. А. Арсеньев указывает, что при приближении к зверям, лежащим на льдине, они лишь поднимают головы и издают при этом короткий свист.
Промысел этого вида тюленей развит очень слабо.
Тюленъ-крабоед (Lobodon carcinophagus) - также типичный антарктический вид, при этом наиболее многочисленный из настоящих тюленей (табл. 39). По размерам тела он несколько мельче предыдущего вида, длина взрослого около 2-2,5 м. Самки и самцы по размерам и окраске неразличимы, но она меняется по сезонам. Во второй половине зимы и в начале весны общий цвет меха серебристо-серый. Осенью после линьки тюлени имеют серовато-коричневую окраску с редкими светлыми пятнами.
Таблица 39 а. Ластоногие. Тюлень-крабоед (Lobodon carcinophagus)
Крабоеды придерживаются области пакового льда, северный предел которого определяет северную границу распространения этого тюленя. Очень редко отдельные звери заходят на север до Австралии и Новой Зеландии. К югу вид встречается до берегового припая Антарктиды. В отличие от тюленя Уэдделла крабоед большую часть года, в том числе и зиму, держится на дрейфующих льдах. Летом, когда у побережья материка мало плавающих льдов, они образуют и береговые залежки. Осенью тюлени в большинстве откочевывают к северу, к кромке плавающих льдов, где и зимуют.
Кормятся мелкими ракообразными, в связи с чем в зубном аппарате есть черты специализации. Многовершинные зубы верхней челюсти входят в промежутки между такими же по устройству зубами нижней челюсти, в результате чего образуется своего рода решетка, свободно пропускающая воду, но задерживающая рачков (рис. 217).
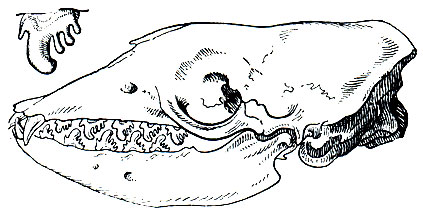
Рис. 217. Череп и зубная система тюленя-крабоеда (Lobodon carcinophagus)
Щенка бывает ранней весной, в сентябре. Новорожденный длиной около 115 см, одет пушистым, густым мехом серовато- коричневого цвета. Период молочного кормления всего около 2-3 недель. Полагают, что молодые крабоеды начинают сходить в воду раньше, чем детеныши большинства других тюленей, быть может, даже в возрасте 2-3 недель.
Крабоед очень энергичное и ловкое животное. Наблюдавшие его советские зоологи (например, В. Арсеньев и В. Земский) особенно поражались той непостижимой ловкости, с которой он выпрыгивает из воды даже на высокие льдины. Они полагают, что эта способность возникла у крабоедов в связи с постоянной угрозой со стороны косаток, от которых они спасаются, выпрыгивая на лед. У многих тюленей на шкурах обнаруживали многочисленные шрамы и свежие раны.
Промысел тюленя-крабоеда развит слабо, хотя местами, например в районе Фолклендских островов, имеются значительные концентрации зверя.
Тюлень Росса (Ommatophoca rossi) - очень редкий зверь, обитающий в самых труднодоступных южных антарктических водах. Скоплений не образует и на льдах держится одиночно (рис. 218).
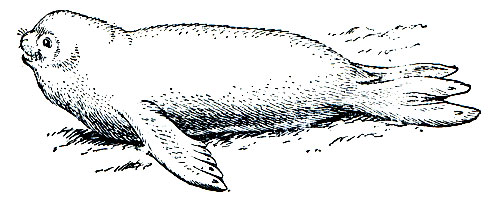
Рис. 218. Тюлень Росса (Ommatophoca rossi)
Внешне этот тюлень легко отличим от других антарктических видов ластоногих. Туловище у него короткое и относительно толстое. Особенно характерна очень толстая, в складках шея, в которую он почти полностью может втягивать голову. Общая окраска меха темно-коричневая, почти черная, более светлая на боках и брюхе. Подкожный жировой слой развит так сильно, что дополняет впечатление о неуклюжести зверя (табл. 39).
Таблица 39 б. Ластоногие. Тюлень Росса (Ommatophoca rossi)
Тюлень Росса способен издавать громкие, мелодичные звуки, природа которых неизвестна. Людей не боится, были случаи, когда к тюленю подходили вплотную и трогали его руками.
Образ жизни почти не известен. При вскрытии желудков в них обнаруживали головоногих моллюсков, реже - ракообразных. Промысел запрещен международным соглашением.
Морской леопард (Hydrurga leptonyx) - антарктический тюлень с очень своеобразной внешностью (табл. 35), прямо противоположной таковой тюленя Росса. Туловище у леопарда относительно длинное, тонкое и стройное. Шея также тонка и длинна. Голова небольшая, и, по мнению некоторых (например, В. Земского), она несколько похожа на голову змеи. В отличие от других тюленей у морского леопарда самцы мельче самок. Максимальная длина тела самцов 3,1 м, самок - 3,6 м. Спина и бока серые, живот почти белый. Граница между участками с окраской очень резкая. На боках располагаются темные пятна. Мех очень короткий, а жировой подкожный слой развит слабее, чем у других антарктических тюленей.
Среди антарктических тюленей морской леопард имеет наиболее широкое распространение, хотя численность его нигде не бывает высокой. Держится, среди льдов, на побережье материка и островов, на плавучих льдах. Образ жизни преимущественно одиночный, и лишь в период размножения иногда наблюдаются небольшие группы тюленей. Отмечены миграции: летом на юг к побережью Антарктиды, зимой - на север. Были случаи обнаружения этого вида у Австралии, Огненной Земли, Патагонии.
Размножаются чаще на льдах, реже на берегу. На Фолклендских островах щенка бывает в сентябре - октябре, на Южной Георгии - в конце августа - начале сентября. В отличие от многих других тюленей окраска новорожденного неотличима существенно от окраски взрослого.
Морской леопард - хищник. Он поедает рыбу, головоногих моллюсков, пингвинов, а изредка и тюленей других видов. Ест также мясо убитых китов. Распространенное ранее мнение о нападении на людей ошибочно. Только при преследовании тюлень может броситься на человека.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'