Класс пресмыкающиеся, или рептилии (Reptilia)
Общий очерк (А. Г. Банников, М. Н. Денисова)
Пресмыкающиеся по сравнению с земноводными представляют собой следующий этап приспособления позвоночных животных к жизни на суше. Это первые настоящие наземные позвоночные, характеризующиеся тем, что размножаются на суше яйцами, дышат только легкими, механизм дыхания у них всасывательного типа (при помощи изменения объема грудной клетки), хорошо развиты проводящие дыхательные пути, кожа покрыта роговыми чешуйками или щитками, кожных желез почти нет, в желудочке сердца имеется неполная или полная перегородка, вместо общего артериального ствола от сердца отходят три самостоятельных сосуда, почки тазовые (метанефрос). У пресмыкающихся увеличивается подвижность, что сопровождается прогрессивным развитием скелета и мускулатуры: меняется положение различных отделов конечностей по отношению друг к другу и к телу, укрепляются пояса конечностей, позвоночник расчленяется на шейный, грудной, поясничный, крестцовый и хвостовой отделы, увеличивается подвижность головы. Череп пресмыкающихся, как и у птиц, в отличие от других позвоночных животных, соединяется с позвоночником одним (непарным) мыщелком. В скелете свободных конечностей характерны межзапястные (интеркарпальные) и межпредплюсневые (интертарзальные) сочленения.
В поясе передних конечностей у них есть своеобразная покровная кость-надгрудинник.
К примитивным особенностям пресмыкающихся как обитателей суши следует отнести наличие двух дуг аорты, смешанную кровь в артериях туловищного отдела, низкий уровень обмена веществ и непостоянную температуру тела.
Современные пресмыкающиеся представляют собой лишь разрозненные остатки богатого и разнообразного мира рептилий, населявшего Землю в мезозойскую эру.
Сейчас насчитывается около 6000 видов рептилий, т. е. почти втрое больше, чем современных земноводных.
Ныне живущих пресмыкающихся делят на 4 отряда: Чешуйчатые, Черепахи, Крокодилы и Клювоголовые.
Наиболее многочисленный отряд чешуйчатых (Squamata), включающий около 5700 видов,- единственная ныне процветающая группа пресмыкающихся, широко распространенная по земному шару и составляющая основную массу рептилий нашей фауны. К этому отряду относятся ящерицы, хамелеоны, амфисбены и змеи.
Значительно меньше черепах (Chelonia) - около 200 видов, представленных в животном мире нашей страны несколькими видами. Это очень древняя группа рептилий, дожившая до наших дней благодаря своеобразному защитному приспособлению - панцирю, в который заковано их тело.
Крокодилы (Crocodylia), которых известно около 20 видов, населяют материковые и прибрежные водоемы тропиков. Они представляют собой прямых потомков древних высокоорганизованных пресмыкающихся мезозоя.
Единственный вид современных клювоголовых (Rhynchocephalia) - гаттерия имеет много чрезвычайно примитивных черт и сохранилась только в Новой Зеландии и на прилежащих мелких островах.
Развитие рептилий, даже тех, которые обитают в воде, не связано с водной средой.
Развитие волокнистых скорлуповых оболочек, видимо, у пресмыкающихся было первым важнейшим преобразованием яйца в ряду приспособлений к наземному развитию. Среди ныне живущих пресмыкающихся можно наблюдать разные стадии изменения яйцевых оболочек, которые служат приспособлением к развитию на суше. Более примитивными формами в этом отношении оказываются скорлуповые оболочки яиц у ящериц и змей, представленные относительно мягкой, пергаментообразной, волокнистой оболочкой, которая довольно близка по химическому составу к оболочкам амфибий. Волокнистая скорлуповая оболочка значительно задерживает высыхание яиц, но предохранить их от этого полностью не может. Развитие идет нормально лишь при влажности почвы не ниже 25%.
Появление скорлуповой оболочки имеет значение не только как защита от высыхания, но и как приспособление к развитию яйца в новых условиях. Так, перенос развития на сушу может осуществиться в полной мере только тогда, когда выпадает личиночная стадия, требующая для своего существования водной среды. Исчезновение личиночной стадии обычно сопровождается увеличением в яйце запаса питательных веществ, обеспечивающего полное развитие зародыша. Увеличение же размеров яйца, особенно в воздушной среде, где удельный вес всякого тела сильно возрастает, возможно только в том случае, если имеется твердая оболочка, не позволяющая яйцу растекаться и сохраняющая его целостность. Следовательно, появление скорлуповой оболочки у яиц ящериц и змей обеспечило не только некоторую защиту их от высыхания, но и увеличение желтка и выпадение личиночной стадии в развитии.
Однако яйцо чешуйчатых еще примитивно. Большая часть воды, необходимая для развития зародыша, воспринимается им из окружающей среды.
Дальнейшим шагом в приспособлении яиц к развитию на суше было развитие белковой оболочки, выделяемой стенками яйцеводов. В ней сосредоточивается запас всей воды, необходимой для развития зародыша. Такая оболочка покрывает яйца черепах и крокодилов. Позднейшие исследования показали, что в яйцах змей (и, видимо, также ящериц) на ранних стадиях развития имеется тонкий слой белковой оболочки. В этот период еще не развиты ни амнион, ни аллантоис. Тонкий слой белка выполняет защитную роль и снабжает желток влагой.
Понятно, что белковая оболочка не могла бы осуществлять своей функции резервуара воды, если бы твердая наружная оболочка не защищала ее хотя бы частично от высыхания. Следовательно, появление скорлуповой оболочки яйца обеспечило не только возможность увеличения желтка, но и появление таких приспособлений в яйце, которые обеспечивают зародыш необходимой для развития водой. С другой стороны, отсутствие необходимости черпать извне воду, нужную для развития зародыша, создает предпосылку для дальнейшего преобразования скорлуповой оболочки. Волокнистая оболочка заменяется у черепах и крокодилов известковой, совершенно не пропускающей воду. Яйца, покрытые известковой оболочкой, прекрасно защищены от высыхания и могут развиваться на суше в любых условиях.
Однако твердые скорлуповые оболочки, ликвидировавшие опасность высыхания яйца, сами по себе представляют серьезную угрозу для развивающегося организма. Увеличивающийся в размерах зародыш может быть раздавлен или поврежден в результате соприкосновения с твердой скорлупой. В связи с этим у пресмыкающихся, так же как и у других наземных позвоночных, образуются особые эмбриональные приспособления, защищающие зародыш от соприкосновения с твердой скорлупой. На ранних стадиях развития начинает образовываться кольцевая складка (рис. 76 ). Она, все больше увеличиваясь, обрастает зародыш, края ее сходятся и срастаются. В результате зародыш оказывается заключенным в амниотическую полость, в которой скапливается особая амниотическая жидкость. Таким образом, зародыш оказывается защищенным от соприкосновения со скорлуповой оболочкой.
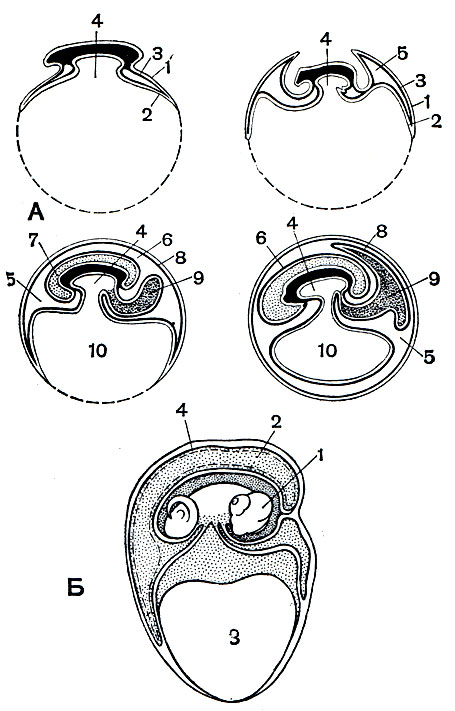
Рис. 76. Последовательные стадии развития зародышевых оболочек у амниот (А) и развивающаяся ящерица (Б). А. 1 - эктодерма; 2 - энтодерма; 3 - мезодерма; 4 - полость кишечника; 5 - внезародышевая полость тела; 6 - амнион; 7 - амниотическая полость, заполненная амниотической жидкостью, в которой плавает зародыш; 8 - сероза; 9 - аллантоис; 10 - желточный мешок. Б. 1 - амнион; 2 - аллантоис; 3 - желток; 4 - сероза
Помещающийся в амниотической полости зародыш испытывает недостаток в кислороде. Затруднено также и выделение продуктов жизнедеятельности развивающегося организма. Следствием образования амниона оказывается развитие и другого зародышевого органа - аллантоиса, или зародышевого мочевого пузыря. Он выполняет функцию органа дыхания, так как его стенки, имеющие богатую сеть кровеносных сосудов, прилегают к оболочкам яйца. Последние благодаря пористости скорлуповой оболочки не препятствуют проникновению кислорода внутрь яйца к кровеносным сосудам аллантоиса. Кроме того, зародыш выделяет в аллантоис продукты распада.
Затруднения с выделением продуктов жизнедеятельности зародыша, развивающегося в замкнутом яйце, решаются не только благодаря развитию аллантоиса, но и вследствие изменения характера обмена веществ в яйце. Основным источником энергии в яйцах земноводных оказываются белки. Продукт их распада - это мочевина, которая легкорастворима и, оставаясь по соседству с зародышем, может проникать обратно в его ткани" отравляя его.
Основу запаса питательных веществ в яйцах пресмыкающихся составляют жиры, распадающиеся на углекислый газ и воду. Газообразные продукты обмена веществ легко выделяются из яиц, развивающихся в воздушной среде, наружу, не причиняя вреда зародышу. Однако и у пресмыкающихся в процессе жизнедеятельности зародыша образуются продукты распада не только жиров, но и белков. Конечным продуктом распада белков у них оказывается не мочевина, а мочевая кислота, отличающаяся слабой способностью диффузии и не могущая поэтому причинить вред зародышу.
Органы размножения - яичники и семенники - пресмыкающихся существенно не отличаются от таковых у земноводных. Изменения в яичниках связаны только с большими размерами откладываемых яиц. Выводные протоки половых желез у представителей этих двух классов, так же как и у всех других наземных позвоночных, гомологичные,т. е. одинаковые по происхождению. Яйцевод, представлен мюллеровым, а семяпровод - вольфовым каналом. Яйцеводы пресмыкающихся отличаются от яйцеводов земноводных изменениями гистологического строения их стенок, выделяющих скорлуповую и белковую оболочки, отсутствующие у земноводных. Что касается вольфова канала, то он перестает выполнять функцию мочеточника и служит исключительно семяпроводом, исчезая в связи с этим у самок.
Черты организации взрослого пресмыкающегося также иллюстрируют дальнейшее приспособление к жизни на суше.
Органы дыхания рептилий отличаются от таковых у земноводных большей дифференцировкой. Они погружены глубже внутрь тела и соединяются с окружающей средой посредством хорошо развитых дыхательных путей. От гортани отходит длинная дыхательная трубка, или трахея, поддерживаемая многочисленными хрящевыми кольцами. Сзади трахея разделяется на две трубки (бронхи), ведущие каждая в соответствующее легкое. Развитие дыхательных путей защищает легкие от иссушающего и охлаждающего действия воздуха, поступающего в организм из окружающей среды. Легкие разрастаются вперед в виде выступов по бокам от бронхов, так что последние открываются не в передний конец легкого, а на боковой его стенке. От этого путь воздуха в легкие становится более извилистым.
Стенки легких у пресмыкающихся имеют несколько более губчатое строение, чем у земноводных. Однако значительное увеличение дыхательной поверхности у пресмыкающихся достигается не за счет этого. Самое существенное преобразование в строении легкого заключается в том, что в нем сокращаются размеры внутренней полости, вытесняемой перекладинами, отходящими от спинной и брюшной поверхности органа. В результате от внутренней полости остается только сравнительно узкий центральный ход, служащий как бы продолжением бронха. Перекладины, вдающиеся в полость легкого, так же как и стенки его, имеют ячеистое строение и богаты кровеносными сосудами. От каждой такой перекладины, разделяющей легкие на камеры, отходят перекладины, разбивающие каждую из этих камер на более мелкие участки. Внутренними краями этих перекладин образуются добавочные дыхательные пути - бронхи второго порядка, делящиеся иногда и дальше на бронхи третьего порядка.
Преобразования в системе органов дыхания приводят к тому, что легкие у рептилий приобретают значение единственного органа дыхания.
Сохранение ребер у пресмыкающихся обеспечило возникновение нового механизма дыхания, отсутствующего у земноводных. Вдох и выдох у них происходит благодаря увеличению и сокращению объема грудной клетки, ограниченной ребрами. Изменение объема трудной клетки производится специальными дыхательными мышцами и возможно благодаря подвижному сочленению ребер как с позвоночником, так и с грудиной.
Вентиляция легких у пресмыкающихся достигает значительно большей интенсивности, чем у земноводных, показывается весьма важным обстоятельством в приспособлении позвоночных животных к жизни на суше.
Совершенствование наземных органов дыхания у пресмыкающихся происходит параллельно с совершенствованием системы органов кровообращения. Основные преобразования сводятся к дальнейшему разделению артериального и венозного тока крови. У большинства пресмыкающихся сердце, так же как и у земноводных, трехкамерное, однако желудочек разделен неполной перегородкой на две половины: правую - венозную и левую - артериальную. При сокращении желудочка его перегородка, прикрепленная к брюшной стенке, доходит до спинной стенки желудочка, вполне разобщая правую и левую его половины. У крокодилов имеются и вовсе обособившиеся правый и левый желудочки, отделенные друг от друга полной перегородкой.
Разделению потоков крови в сердце способствует также распадение у пресмыкающихся общего артериального ствола на три самостоятельных сосуда: на легочную артерию, делящуюся затем на два сосуда, и на правую и левую дуги аорты (рис. 77). Каждая дуга аорты загибается назад вокруг пищевода, и, сойдясь друг с другом, они соединяются в непарную спинную аорту. Спинная аорта тянется назад, отсылая по пути артерии ко всем органам. От правой дуги аорты, отходящей от левого артериального желудочка, ответвляются общим стволом правая и левая сонные артерии, от правой же дуги отходят и обе подключичные артерии, несущие кровь к передним конечностям. Благодаря этому передний отдел тела получает артериальную кровь. Однако у всех пресмыкающихся, в том числе и у крокодилов, полного разделения артериальной и венозной крови не происходит, так как эти два потока смешиваются в спинной аорте.
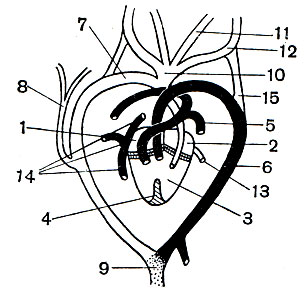
Рис. 77. Сердце и основные сосуды ящерицы (светлым - обозначены сосуды с артериальной кровью; точками - со смешанной кровью; черным - с венозной): 1 - правое предсердие; 2 - левое предсердие; 3 - желудочек; 4 - неполная перегородка, разделяющая желудочек на правую и левую половины; 5 - легочная артерия; 6 - левая дуга аорты; 7 - правая дуга аорты; 8 - подключичная артерия; 9 - спинная аорта; 10 - общий ствол сонных артерий; 11 - левая внутренняя сонная артерия; 12 - левая наружная сонная артерия; 13 - легочная вена; 14 - задняя полая вена и передние полые вены; 15 - сонный придаток
Кожа, освободившаяся от функции дыхания, претерпевает существенные изменения, направленные к защите организма от высыхания. Кожные железы у пресмыкающихся отсутствуют, так как необходимость в смачивании кожи отпала. Испарение влаги с поверхности тела уменьшилось, так как все тело этих животных покрыто роговыми чешуями.
Полный разрыв с водной средой приводит к тому, что осмотическое давление в теле пресмыкающихся становится независимым от окружающей среды. Ороговение кожи, которое делает ее непроницаемой для воды, снимает угрозу изменения осмотического давления даже тогда, когда пресмыкающиеся вторично переходят к водному образу жизни.
Так как вода поступает в тело пресмыкающихся только произвольно вместе с пищей, то осморегулирующая функция почек почти полностью отпадает. У пресмыкающихся нет надобности, как у земноводных, выводить постоянно возникающий избыток воды из организма. Наоборот, у них, как у сухопутных животных, появляется необходимость экономно расходовать воду, находящуюся в организме. Туловищные почки (мезонефрос) земноводных заменяются у пресмыкающихся тазовыми (метанефрос). Тазовые почки не имеют связи с вольфовым каналом, и у них развиваются собственные мочеточники, впадающие в клоаку. При этом вольфов канал у самок редуцируется, а у самцов выполняет функцию семяпроводов.
Особенности строения тазовых почек пресмыкающихся заключаются в том, что общая фильтрационная площадь клубочков у них уменьшается, а протяженность канальцев увеличивается. Чем меньше клубочки, тем менее интенсивно идет фильтрация воды из организма, В канальцах же, имеющих значительную длину, большая часть воды, отфильтрованной клубочками, всасывается обратно. Следовательно, выделение продуктов жизнедеятельности у пресмыкающихся происходит с затратой минимума воды, входящей в состав организма.
У широко расселившихся по суше многочисленных пресмыкающихся, в отличие от земноводных, наблюдается большая дифференциация пищеварительного тракта, позволяющая полнее использовать попавшуюся животному добычу. Желудок пресмыкающихся имеет толстые мышечные стенки и четко обособлен, так же как и отделы кишечника. На границе тонкой и толстой кишок расположена слаборазвитая слепая кишка, отсутствующая у земноводных. Большая печень рептилий имеет желчный пузырь.
Поджелудочная железа в виде длинного плотного тела лежит в петле двенадцатиперстной кишки.
Несмотря на прогрессивное развитие органов дыхания, кровообращения, пищеварения и покровов, пресмыкающиеся остаются все-таки на таком уровне развития, что теплокровность организма не может быть обеспечена и, подобно земноводным, температура тела этих животных зависит от температуры окружающей среды.
У пресмыкающихся идет дальнейшее совершенствование возможностей ориентации в окружающей среде, и благодаря дальнейшему развитию органов чувств они получают более полную информацию из внешнего мира.
В органах зрения пресмыкающихся, в их ресничном теле развивается поперечнополосатая мускулатура. При аккомодации хрусталик теперь не только перемещается, но и изменяет свою кривизну.
В органах слуха также происходят прогрессивные изменения, выражающиеся в больших размерах улитки и в том, что в слуховой капсуле ниже овального окна появляется второе отверстие, затянутое перепонкой,- круглое окно. Благодаря этому эндолимфа, заполняющая внутреннее ухо, приобретает большую подвижность, что ведет к лучшей передаче ею звуковых колебаний от барабанной перепонки через слуховую косточку к воспринимающим рецепторам во внутреннем ухе.
Органы обоняния по сравнению с аналогичными органами земноводных отличаются более развитой складчатостью, образуемой носовыми раковинами, полным обособлением от органа обоняния якобсонова органа, хорошо развитого у ящериц и змей, и удлинением носоглоточного хода в связи с развитием вторичного нёба. Внутренние ноздри - хоаны отодвигаются ближе к глотке, что обеспечивает возможность свободного дыхания в то время, когда пища находится во рту.
Головной мозг пресмыкающихся продолжает развиваться, приобретая свойства, характерные для наземных животных. Передний мозг у них сблизился с промежуточным, а с обонятельными луковицами, как правило, соединяется длинным обонятельным трактом. Передний мозг перестал быть преимущественно обонятельным и выполняет уже ведущую роль в организации поведения; эта функция перешла к нему от среднего мозга. Размеры переднего мозга стали больше, они заметно превосходят размеры других частей головного мозга. Увеличение переднего мозга идет главным образом за счет развития подкорковых узлов - полосатых тел. При этом прогрессивно развивается и первичный мозговой свод - архипаллиум. У пресмыкающихся четко дифференцируются закладки вторичного мозгового свода - неопаллиума, едва намечающегося у земноводных. Развитие вторичного мозгового свода знаменует собой дальнейшее увеличение числа нервных клеток, которые не только выстилают желудочки переднего мозга, но и появляются на его боковых поверхностях. Развивающийся мозговой свод наползает на промежуточный и средний мозг, которые становятся видимыми только снизу. По сравнению с земноводными мозжечок у пресмыкающихся более выпуклый вследствие разрастания верхней поверхности. Сильнее развивается и корпус мозжечка. Теменной орган у ящериц и гаттерии достигает исключительного развития. В нем можно различить образования, сходные с хрусталиком и сетчаткой. Наблюдения показывают, что у многих ящериц этот орган даже функционирует как светочувствительный аппарат, особенно остро реагирующий на длинные световые волны.
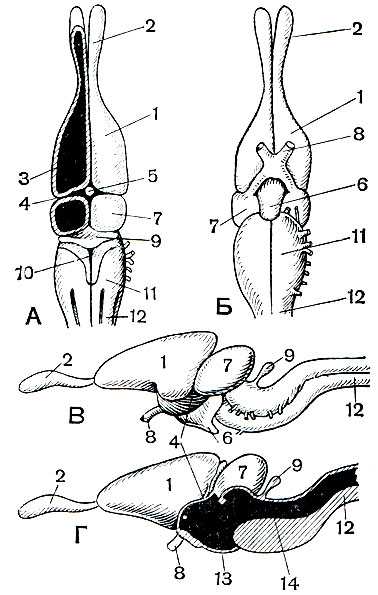
Рис. 78. Головной мозг пресмыкающихся: А - вид сверху; Б - вид снизу; В и Г - вид сбоку (Г - в разрезе): 1 - полушарие переднего мозга; 2 - обонятельная доля; 3 - полосатое тело; 4 - промежуточный мозг; 5 - теменной орган; 6 - гипофиз; 7 - средний мозг; 8 - перекрест зрительных нервов (хиазма); 9 - мозжечок; 10 - четвертый желудочек; 11 - продолговатый мозг; 12 - спинной мозг; 13 - третий желудочек; 14 - сильвиев водопровод
Общее совершенствование строения конечностей у наземных позвоночных животных сводится к тому, что у них относительно удлиняются в передней конечности плечо и предплечье, в задней - бедро и голень, а размеры конечных отделов - площадь ступни и кисти - уменьшаются. Изменяется также и положение отдельных частей конечности по отношению к телу.
Такое удлинение конечностей, сокращение их площади опоры и изменение положения отдельных частей по отношению к телу и друг к другу способствуют переходу от ползания к хождению и бегу. Не-одинаковое расположение отделов в передней и задней конечностях соответствует их различной функции: при передвижении животного передняя конечность тянет тело, а задняя толкает его. У низших наземных позвоночных наблюдаются различия в положении только предплечья и голени, а у высших эти различия распространяются на плечо и бедро.
По характеру описанных выше строения и положения конечностей пресмыкающиеся занимают промежуточное положение между земноводными и млекопитающими.
Кроме того, у пресмыкающихся ребра срастаются с грудиной, что значительно укрепляет пояс передних конечностей, который оказывается благодаря этому связанным с позвоночником. Тазовый пояс рептилий по сравнению с таковым у земноводных также прочнее укреплен, так как срастается не с одним, а с двумя крестцовыми позвонками.
Туловищный отдел позвоночника разделяется у пресмыкающихся уже на два отдела: грудной и поясничный. Грудные позвонки отличаются от поясничных тем, что к ним прикрепляются ребра. Развивается и крестцовый отдел за счет увеличения числа крестцовых позвонков. Удлиняется шейный отдел, и преобразовывается строение двух первых шейных позвонков. Первый из них (атлант) приобретает форму кольца, а второй (эпистрофей) оказывается снабженным зубовидным отростком, входящим в кольцо атланта. Благодаря такому устройству голова приобретает способность не только двигаться в вертикальной плоскости относительно первого шейного позвонка, но и поворачиваться. В последнем случае череп вместе с первым шейным позвонком может поворачиваться на зубовидном отростке второго позвонка. Увеличение подвижности головы, несущей органы чувств, наряду с удлинением шейного отдела способствует лучшей ориентации животного во внешней среде.
В отличие от современных земноводных череп пресмыкающихся весь костный. Исходно, так же как у вымерших земноводных, череп пресмыкающихся имел вид сплошного костного образования, как бы панциря, покрывавшего голову сверху, снизу и с боков. Оставались только отверстия для глаз, ноздрей и теменного органа. Такой тип черепа с небольшими изменениями сохранился у современных черепах. Череп других ныне живущих пресмыкающихся претерпевает частичное исчезновение костного панциря, одевающего череп, так как сплошной панцирь стесняет возможность развития челюстной мускулатуры, расположенной между костным панцирем и собственно черепной коробкой. При этом исчезновение панциря происходило различными путями у различных групп пресмыкающихся, определив подразделение класса рептилий на подклассы.
Приобретение пресмыкающимися способности размножаться на суше путем откладки крупных яиц, одетых плотной оболочкой, освободило их от необходимости возвращаться в водоем - они стали вполне наземными животными.
Ороговение кожи и потеря кожей дыхательной функции сделали пресмыкающихся независимыми и от влажности окружающей среды, позволив им заселить области с низкой влажностью. Ороговевшая кожа также сделала доступными для рептилий засоленные почвы и морскую воду - места обитания, совершенно недоступные земноводным. Вместе с этим произошло усиление дыхательной функции легких за счет усложнения их внутреннего строения и замены глоточного дыхания дыхательными движениями грудной клетки.
Расселившись во все климатические области земного шара, за исключением приполярных областей, пресмыкающиеся дали очень большое разнообразие жизненных форм. Среди них есть наземные, подземные, водные, древесные. Среди ископаемых рептилий были такие, которые имели крылья и летали в воздухе.
Однако сохранившаяся от предков зависимость температуры тела от температуры окружающей среды, т. е. пойкилотермия, связанная с низким уровнем обмена веществ, определяет как специфику распространения, так и многие биологические черты пресмыкающихся.
Пресмыкающиеся, в отличие от земноводных, широко расселились не только во влажных тропиках, но и по пустыням, которые очень благоприятны по температурным условиям. Однако по мере движения к полюсам число видов рептилий падает. Например, у нас в Средней Азии обитает около 50 видов пресмыкающихся, в средней части Европы - 15 видов, а у Северного полярного круга - всего 2 вида.
Правильная сезонная или суточная смена периодов покоя и активности носит у пресмыкающихся несколько иной характер, чем у земноводных. Цикличность рептилий - это биологическое приспособление к неблагоприятным условиям температуры и кормности мест обитания. С условием влажности в такой степени, как у земноводных, она не связана.
Суточная цикличность связана с температурой. Пресмыкающиеся активны при оптимальных для каждого вида температурах. Оптимум же у всех рептилий лежит в области высоких температур и колеблется у разных видов в пределах 20-40°С.
Будучи теплолюбивыми, большинство пресмыкающихся умеренных широт - дневные, немногие виды - сумеречные и только гекконы - ночные животные. В тропических пустынях, напротив, очень много ночных видов рептилий, так как днем там очень жарко.
Даже в умеренных широтах днем температура почвы бывает выше оптимальной, а в Средней Азии она нередко достигает 60°С и выше. При такой температуре все рептилии могли бы погибнуть через несколько минут от перегрева. Но, перемещаясь в течение суток, они выбирают те участки биотопа, где температура в данное время ближе всего к оптимуму. Утром держатся на солнцепечных склонах барханов, затем переходят на теневые склоны, при нагревании почвы взбираются на гребень бархана или куст и т. д. Путем таких перемещений рептилии избегают перегрева и поддерживают температуру тела на высоком и относительно постоянном уровне, близком к температуре тела гомойотермных (теплокровных) животных, т. е. они оказываются "экологически теплокровными" животными. Особенность почвы удерживать дневное тепло используется ночными видами рептилий. Так, сцинковый геккон, охотясь ночью, время от времени закапывается в теплый песок, т. е. активность его распадается на ряд периодов.
Будучи биологическим приспособлением, суточный цикл активности меняется в разные сезоны в зависимости от условий температуры. Так, весной пресмыкающиеся активны только в середине дня - в наиболее теплые часы суток. В середине лета активность рептилий многих видов приурочена, напротив, только к утренним и вечерним часам.
Большое значение в терморегуляции пресмыкающихся имеет солнечная радиация. Так, в горах при температуре воздуха минус 5°С и поверхности почвы плюс 5°С температура тела ящерицы может достигать на солнце 19°С.
Зависимость активности от температуры у змей не столь очевидна. Проглатывая целиком очень большую добычу (до 2/3-3/4 своего веса) и медленно переваривая ее, змеи могут быть неактивны 5-8 дней. Голодная же змея выходит из убежища и при довольно низких темпера-турах, далеких от оптимума.
Сезонная цикличность, как и у земноводных, служит приспособлением, позволяющим обитать в зонах, пригодных для существования лишь в теплый период года. У рептилий тропиков нет правильной сезонной смены периодов активности и покоя. Она слабо выражена в субтропиках и появляется лишь в умеренных широтах, где пресмыкающиеся впадают в зимнюю спячку.
Продолжительность активного периода меняется в зависимости от температурных условий. Так, живородящая ящерица (Lacerta vivipara) в Южной Европе активна 9 месяцев, в средней полосе Европейской части СССР - 5,5 месяца, а на севере - еще меньше.
У пресмыкающихся разных видов продолжительность зимней спячки различна и связана с холодоустойчивостью вида. Так, обыкновенная гадюка (Vipera berus) появляется весной раньше других обитающих в данной местности змей и позже них уходит на зимовку.
При благоприятных температурных условиях большинство пресмыкающихся может не впадать в зимнюю спячку. Например, в теплых постройках человека в Средней Азии обычно впадающие в зимнюю спячку агамы и гекконы оказываются активными всю зиму. В связи с относительно теплыми зимами и частыми оттепелями в Средней Азии гюрза, эфа, щитомордник, ряд видов ящурок и круглоголовок регулярно появляются на поверхности земли и греются на солнце, так что у них нет непрерывной зимней спячки.
Характер убежищ, где зимуют пресмыкающиеся, весьма различен. Большинство зимует в норах грызунов, трещинах почвы, в пустотах, образовавшихся на месте сгнивших корней, и т. д. Излюбленные места зимовок живородящей ящерицы - полости, образовавшиеся в болотистых местах под дерном при опускании уровня грунтовых вод. Ужи в большом количестве собираются на зиму в навозных кучах и подвалах. Полозы нередко зимуют в пещерах, болотные черепахи - на дне водоемов.
Большинство пресмыкающихся зимует поодиночке или небольшими группами в 2-3 особи. Однако некоторые образуют зимой большие скопления. Так, обыкновенные ужи собираются в клубки по нескольку сотен и даже тысяч особей; в клубках зимующих гадюк и щитомордников находили десятки змей. Среди ящериц нашей фауны известны большие скопления зимующих золотистых мабуй (Mabuya aurata) в Туркмении.
К периоду зимовки у пресмыкающихся накапливаются запасные питательные вещества, а интенсивность обмена веществ резко падает.
У некоторых пресмыкающихся наблюдается летняя спячка, однако она определяется не влажностью, как у земноводных, а отсутствием полноценного корма. Так, в. пустынях Средней Азии в летнюю спячку впадает среднеазиатская черепаха (Testudo horsfieldi). В конце мая или начале июня, вслед за высыханием растительности, которой они питаются, черепахи выкапывают норы и впадают в оцепенение. В тех местах, где растительность не высыхает, например на орошаемых полях, черепахи активны все лето. Следовательно, цикличность в этом случае зависит от кормности мест обитания.
Колебания численности пресмыкающихся по годам относительно невелики. Для немногих изученных в этом отношении видов известно изменение численности за смежные годы в 2-3 раза. Основной причиной падения численности является гибель в местах зимовок. В отдельных случаях численность падает в результате деятельности хищников.
Корма пресмыкающихся, как и способы добывания пищи, весьма разнообразны; среди пресмыкающихся встречаются насекомоядные, рыбоядные, хищники, растительноядные и т. д.
Насекомыми кормится большинство ящериц. Круглоголовки (Phrynocephalus) в основном ловят муравьев, мелких жуков, ящурки (Eremias) - прямокрылых и полужесткокрылых, настоящие ящерицы (Lacerta) питаются жуками, паукообразными и моллюсками. Моллюски составляют основу питания веретеницы и желтопузика. Самые крупные ящерицы - вараны, кроме насекомых, ловят мышевидных грызунов и птиц. Насекомыми кормятся немногие змеи, например слепозмейка, эйренисы (Eirenis). Большую долю они составляют в питании степной гадюки. Головоногими моллюсками и ракообразными кормится морская пеламида (Pelamis); ракообразными, моллюсками, насекомыми, реже рыбой - пресноводные-черепахи.
Основу питания большого числа змей составляют позвоночные животные. Обыкновенный уж и большеглазый полоз - (Ptyas) ловят главным образом земноводных; водяной уж (Natrix tessellata) и морские черепахи - рыб; стрела-змея, краснополосый полоз (Coluber rhodorhachis), волкозуб (Lycodon), афганский литоринх (Lytorhynchus) - ящериц; гадюки - мышевидных грызунов. Грызуны составляют основу питания эфы (Echis carinatus), гюрзы (Vipera lebetina) и большинства наших полозов. Некоторые полозы, например амурский (Elaphe schrenki), хорошо лазая по деревьям, ловят в гнездах птенцов и взрослых, птицы составляют заметную долю также в питании гюрзы. Есть змеи, пожирающие других змей. Так, распространенная у нас в Нижнем Поволжье и на Кавказе ящеричная змея (Malpolon monspessulanus) кормится не только ящерицами, но и степными гадюками, предпочитая последних.
Змеи вообще наиболее специализированы по питанию. Так, например, африканская яичная змея (Dasypeltis) кормится только яйцами птиц, в связи с чем у нее имеются своеобразные приспособления. Зубной аппарат этой змеи развит очень слабо, зато передние позвонки имеют направленные вперед отростки, которые прободают пищевод, вдаваясь в его полость. Змея заглатывает яйцо целиком, и оно, проходя через пищевод, разрезается отростками позвонков, после чего жидкое содержимое яйца стекает по пищеводу, скорлупа же выбрасывается через рот. Крупные тропические удавы нападают на таких больших животных, как обезьяны и мелкие копытные.
Неядовитые змеи пожирают добычу живьем (например, ужи - лягушек) или предварительно душат ее, обвиваясь вокруг жертвы кольцами. Однако, сдавливая добычу, они не ломают ее костей, которые могли бы поранить змею при заглатывании добычи. Ядовитые змеи предварительно убивают жертву молниеносным броском, вонзая в нее свои ядовитые зубы, после чего заглатывают целиком.
Реже всего встречаются растительноядные пресмыкающиеся. Растительноядные почти все сухопутные черепахи, в том числе и наши - среднеазиатская и средиземноморская черепахи (Testudo horsfieldi, Т. graeca). В питании немногих ящериц, главным образом тропических агам и игуан, также значительную роль могут играть растительные корма. Среди наших ящериц агамы иногда питаются цветками и плодами, а длинноногие сцинки (Eumeces schneideri) - ягодами шелковицы.
Большинство пресмыкающихся активно разыскивает добычу. Так, пользуясь слухом и зрением, отыскивают добычу гекконы, сцинки, ящурки, ящерицы, вараны. Отыскивают добычу, используя главным образом осязание, кобра, эфа, щитомордник, стрела-змея, полозы. Напротив, подстерегают свою добычу, подобно кошкам, агамы, круглоголовки, а из змей - гюрза, удавчики.
У пресмыкающихся как настоящих наземных позвоночных животных размножение и развитие происходит на суше.
Даже ведущие вполне водный образ жизни крокодилы, морские змеи и морские черепахи выходят в период размножения на сушу.
Оплодотворение у рептилий внутреннее, и у всех, кроме гаттерии, имеются органы совокупления. У большинства видов в период размножения наблюдается "гон" - повышенная активность, нередко сопровождаемая у черепах и ящериц драками самцов.
Большинство пресмыкающихся откладывает небольшое число (8-16) крупных яиц, одетых кожистой или твердой известковой скорлупой (черепахи, крокодилы), Благодаря плотным кожистым и известковым оболочкам яйца пресмыкающихся могут развиваться на суше при незначительной влажности (от 3 до 15%). Они откладываются обычно в почву или, как у гекконов и у некоторых агам и игуан, в трещины скал и под кору деревьев. Крокодил откладывает яйца в почву или в гнезда из сухих листьев и травы.
Скорость развития яиц зависит от температуры и в связи с этим может изменяться в 2-3 раза. У большинства видов нашей фауны развитие начинается лишь при 12-15°С, но температура в 40-42°С оказывается губительной; обычно температура в местах развития яиц бывает равна 20-30°С. Пресмыкающиеся пустынь чаще откладывают яйца в теневых местах и на значительной глубине. Напротив, ящерицы в умеренных широтах выбирают хорошо прогреваемые солнцем места. Обыкновенный уж откладывает яйца в кучи гниющих веществ, где температура оказывается на 3-5°С выше температуры окружающей среды. В умеренных широтах срок развития яиц пресмыкающихся - 2-3 месяца.
У змей бывает одна кладка яиц в год, у гекконов - две. Ящерицы и черепахи откладывают по 3-4 и более кладок в год. Число яиц в кладках зависит от количества кладок, размера яиц и возраста животных. Всего пресмыкающиеся откладывают несколько десятков яиц в год, т. е. значительно меньше, чем земноводные. Меньшая плодовитость рептилий связана с более совершенным строением яйца.

Таблица 17. Черепаха матамата (Chelus fimbriata)
В тех местах, где много животных, поедающих яйца, кладки охраняются родителями (крокодилы, кобры, некоторые удавы и др.). У питонов, обвивающих кладку своим телом, наблюдаются явления "насиживания"; при этом температура тела змеи поднимается на 2-4°С выше температуры окружающего воздуха.
Среди пресмыкающихся довольно широко развито живорождение. Оно встречается только у форм с мягкими яйцевыми оболочками, благодаря которым яйца сохраняют возможность водного обмена со средой. У черепах и крокодилов, яйца которых обладают развитой белковой оболочкой и скорлупой, живорождение не наблюдается. Первый шаг к живорождению - задержка оплодотворенных яиц в яйцеводах, где и происходит частичное развитие. Так, у прыткой ящерицы яйца могут задерживаться в яйцеводах на 15-20 дней и в отложенных яйцах оказывается зародыш, сформированный более чем на 1/3. На 30 дней может происходить задержка у обыкновенного ужа, так что в отложенном яйце оказывается наполовину сформированный эмбрион. При этом чем севернее район, тем, как правило, происходит более длительная задержка яиц в яйцеводах.
У других видов, например у живородящей ящерицы, веретеницы, медянки и др., яйца задерживаются в яйцеводах до момента выклева зародышей. Такое явление получило название яйцеживорождения, поскольку развитие идет за счет запасных питательных веществ в яйце, а не за счет материнского организма.

Таблица 18. Гаттерия (Sphenodon punetatus)

Таблица 18. Гаттерия (Sphenodon punetatus)
Наконец, у некоторых видов сцинков, ящериц и гадюк яйцевые волокнистые оболочки редуцируются и часть яйцевода соприкасается с хорионом. Хорион может образовать выступы-ворсинки, внедряющиеся в стенки яйцевода. Образуется подобие плаценты млекопитающих. При этом заметно уменьшается количество желтка в яйцах, и питание идет в значительной мере за счет материнского организма (Lygosoma, Seps, Vipera berus и др.). Подобные случаи носят название истинного живорождения.
Основной причиной, вызывающей яйцеживорождение у пресмыкающихся, следует считать холодный климат. В этом можно убедиться, обратившись к карте распространения живородящих форм. Процент живородящих видов повышается при движении к северу и в горы. Весьма характерно, что виды, распространенные далее всего к северу, так же как и виды, поднимающиеся выше в горы, живородящи. Примером могут служить тадюки и живородящая ящерица. При этом один и тот же вид, например тибетская круглоголовка (Phrynocephalus teobaldi), на высоте 2000 м яйцекладущ, а на высоте 4000 м оказывается живородящим.
У пресмыкающихся защитные приспособления не только носят пассивный характер, как у земноводных, но и могут принимать форму активной защиты.
К пассивным защитным приспособлениям относится приспособительная окраска, достигающая у многих видов большого совершенства. Преобладающее число рептилий окрашено под цвет субстрата, на котором они постоянно держатся. При этом часто встречается расчленяющая окраска и приспособительная окраска глаз. Особого совершенства достигла приспособительная окраска у плетевидных змей, гекконов и хамелеонов. Последние получили широкую известность благодаря своей способности быстро менять окраску в зависимости от условий окружающей среды.
Приспособительное значение имеет контрастная и яркая окраска многих ящериц и змей. При опасности такие пресмыкающиеся выставляют ярко окрашенную часть тела напоказ, принимая при этом устрашающие позы. У кобр хорошо известны складки кожи по бокам шеи, которые змея расширяет, поворачиваясь к противнику контрастными полосами, или очками. Раздувают шею, хотя и не в такой степени, многие полозы и ужи. Агамы и круглоголовки открывают при опасности рот и раздувают горловой мешок. Ушастая круглоголовка дополнительно раскрывает складки кожи в углах рта, которые наливаются кровью и становятся красно-синими. Огромных размеров достигает такая складка у австралийской плащеносной ящерицы, принимающей при опасности соответствующую позу.
К пассивным способам защиты относится панцирь черепах, получивший наибольшее развитие у сухопутных форм, вынужденных в связи с растительноядностью быть активными большую часть суток.
Активная защита состоит в приспособительном поведении. Некоторые круглоголовки, а из змей эфа, спасаясь от врага, закапываются в песок, начиная быстро перемещать тело то влево, то вправо, и как бы утопают в нем. Эфа, как и некоторые другие змеи, если не может сразу скрыться, ползет вспять, но благодаря особым движениям тела создает впечатление нападения.
Ряд рептилий издает предостерегающе-устрашающие звуки. Громко шипят многие сухопутные черепахи. Шипят почти все змеи, а гремучие змеи шелестят роговыми кольцами хвоста. Скрипящий звук хвостом издает сцинковый геккон.
Большинство круглоголовок и ящурок роет на охотничьем участке большое количество защитных нор, куда они скрываются при опасности.
Наконец, довольно широкое распространение имеет активная защита - нападение. Так действуют при опасности почти все круглоголовки. Нападает варан, который способен нанести не только значительную и долго болящую рану зубами, но и очень сильный удар хвостом. Из ящериц - ядозубы (Heloderma) могут при укусе вызвать тяжелое состояние отравления у человека.
Ядовитые железы - довольно частое явление у змей. Для человека ядовитыми следует считать около 450 видов, из них 10 видов обитает в пределах СССР. Видоизмененная слюнная железа, выделяющая яд, появляется у ряда видов, не имеющих ядовитых зубов, например у удавчиков и ужей. Яд последних очень слаб, но на холоднокровных действует подобно яду кобры.
Для настоящих ядовитых змей характерно появление специализированных зубов. Ядовитые зубы по своему расположению могут быть двух типов: заднебороздчатые (Opistoglypha) и переднебороздчатые (Proteroglypha). У змей с зубами первого типа, например у стрелы-змеи или ящеричной змеи, ядовитые зубы сидят на заднем конце верхнечелюстной кости и имеют бороздку для стекания капли яда. Этот яд действует на холоднокровных. У змей с зубами второго типа, например у гадюк, кобры, щитомордника, ядовитые зубы сидят на переднем конце верхнечелюстной кости, сильно увеличены и чаще имеют внутренний канал. Яд этих змей действует и на теплокровных животных. На различных животных яд действует по-разному. Так, одно и то же количество яда кобры может убить 10 змей, 25 собак, 60 лошадей и 300 тыс. голубей.
В пустынях и степях, где численность пресмыкающихся высока, они играют заметную роль в биоценозах. Большинство ящериц и змей, уничтожая значительное количество вредных насекомых, моллюсков и грызунов, приносит пользу сельскому хозяйству. Многие ящерицы в пустынях служат кормом для ряда важных промысловых зверей, например лисицы и хорька.
Некоторые виды приносят местами заметный вред. Среднеазиатская черепаха в ряде районов оказывается вредителем на бахчах и особенно в питомниках фисташки; вредная деятельность этой черепахи усугубляется прокапыванием ею насыпей и стен оросительной системы.
На рыборазводных станциях серьезный вред приносят водяные ужи, уничтожающие большое количество мальков промысловых рыб: карпа, лосося, осетра. В районах рыборазведения водяные ужи подлежат полному уничтожению, что лучше всего делать ранней весной на местах их зимовок.
Вероятна отрицательная роль ряда пресмыкающихся, особенно сухопутных черепах, некоторых ящериц и змей, в распространении ряда заболеваний человека и домашних животных, поскольку на них живут нимфы и взрослые иксодовые клещи.
В некоторых странах серьезный вред человеку приносят ядовитые змеи. Количество жертв от ядовитых змей обычно сильно преувеличено, но все же в жарких странах, особенно в Юго-Восточной Азии и Южной Америке, ежегодно от укусов змей умирают тысячи людей. В Западной Европе далеко не ежегодно регистрируются единичные случаи смерти людей от укусов ядовитых змей. В СССР в республиках Средней Азии, где наблюдается наибольшая численность ядовитых змей, ежегодно от их укусов умирает до 12 человек и смертельные случаи составляют около 6% укушенных (главным образом дети, укушенные в лицо). Прежде, до применения сывороток, укусы наиболее ядовитых змей - кобры, гюрзы, эфы - давали от 15 до 30% смертельных случаев, гадюки - 5-6%. Сейчас смертельные исходы бывают в 1,5-2% случаев укусов (см. очерк "Змеи").
В зависимости от характера действия яда различают две группы ядовитых змей. Человек, укушенный змеей I группы, к которой относятся гюрза, эфа, гадюка, ощущает сильную боль, окружающая ранку ткань краснеет и опухает. Развивающийся отек распространяется от места укуса на всю конечность. Появляется сонливость, выступает холодный пот. В ряде случаев бывает лихорадочное состояние, тошнота, бред. Вследствие подкожных точечных кровоизлияний на теле часто появляются фиолетово-красные пятна. В тяжелых случаях наступает некроз тканей, образуются нарывы и язвы. Может наступить смерть.
Человек, укушенный змеей II группы, яд которой действует через нервную систему (к ним относится кобра), обычно не чувствует боли в месте укуса. Однако вскоре после укуса развивается слабость в конечностях - начинается паралич. Распространяясь, паралич захватывает все органы, затрудняет дыхание, речь, опускаются веки, парализуются губы, язык. Лицо синеет. В тяжелых случаях наступает смерть.
Большое значение для лечения укушенного имеет своевременное оказание помощи. С укушенного места следует снять остатки яда крепким раствором марганцовокислого калия, давать больному крепкий чай, кофе и обеспечить полный покой и тепло. Больного необходимо как можно скорее доставить в больницу, где ему будет введена специальная сыворотка. Сыворотка хранится в ампулах или в сухом виде с указанием дозировки. Обычно вводят под кожу 10 см3, но в тяжелых случаях прибегают к венозным инъекциям большими дозами.
Увеличивать ранку надрезами для усиления кровотечения бесполезно. Только вред могут принести прижигание, прикладывание всевозможных "змеиных" камней, земли и т. д. Наложение жгута - опасное и практически также бесполезное средство, так как оно не задерживает всасывания яда. В большинстве случаев тяжелые последствия после укуса ядовитых змей бывают вызваны не действием яда, а прижиганиями, кровопусканиями, перетягиванием конечностей жгутом выше места укуса и т. д.
Змеиный яд применяется для приготовления лечебных сывороток против укусов змей. Сыворотка приготавливается из крови лошади, в организм которой вводят возрастающие дозы яда в течение 16 месяцев. В результате кровь приобретает такой иммунитет, что животное может переносить впрыскивание 80-кратной смертельной дозы.
Изготовляют моновалентные сыворотки, специфичные по отношению к яду одного вида змей, и поливалентные, действующие против яда нескольких видов змей.
Из яда гадюк приготавливают также лечебные препараты, например випратокс, випраксин, випразид и другие, применяемые при лечении радикулита, ишиаса, ревматизма. Из яда гюрзы и щитомордника изготовляют кровоостанавливающие средства (лебетокс), из яда гремучей змеи - препараты, излечивающие эпилепсию, яд кобры употребляют для изготовления препаратов, применяемых при лечении бронхиальной астмы и ряда других заболеваний. Яд змей широко применяется в биохимии.
Для получения яда организуют питомники змей. Первый такой питомник - серпентарий - был создан в Бразилии около города Сан-Пауло (Бутантан); он и сейчас самый крупный в мире. Подобные питомники имеются в Индии, Индонезии, Южной Африке и других странах. У нас питомники организованы в Ташкенте, Фрунзе, Бадхызе. В питомниках яд у змей берут постоянно, не убивая их (см. очерк "Змеи").
Пресмыкающиеся служат источником сырья для промышленности. Издавна кожа крокодила, крупных змей и ящериц идет на изготовление чемоданов, портфелей, обуви и т. д. Особенно ценится кожа крокодила. В начале этого века крокодилов заготовляли по 500 тыс. экземпляров в год. Местами численность их настолько сократилась, что они были взяты под охрану и сделаны попытки искусственного разведения их. Так, фермы Флориды содержали до 12 тыс. крокодилов, поставляя на рынки до 3000 экземпляров в год. Среди наших видов как сырье для кожевенной промышленности имеют значение вараны и крупные полозы. Используется панцирь некоторых морских черепах, особенно биссы, роговые щитки которых в горячей воде и под давлением спрессовываются в однородную массу. Из этой высококачественной по пластическим свойствам красивой массы изготовляют гребни, оправы для очков и т. д.
Некоторые рептилии употребляются человеком в пищу, наиболее часто мясо и яйца черепах. Существует регулярный промысел мягкокожистых черепах и некоторых видов морских черепах, особенно зеленых. В Северной Америке промышляют террапинов, которых стали разводить и на фермах. Среди наших видов вполне съедобно мясо средиземноморской, среднеазиатской и дальневосточной мягкокожистой черепах. Можно употреблять в пищу желтки яиц всех наших черепах.
Мясо черепах идет также на откорм пушных зверей. Так, звероводческие хозяйства Казахстана ежегодно заготовляют более 100 тыс. среднеазиатских черепах.
Ископаемые пресмыкающиеся представляют исключительный интерес, так как к ним принадлежат многочисленные группы, когда-то господствовавшие на земном шаре. Древние группы этого класса дали начало не только современным пресмыкающимся, но и птицам и млекопитающим.
Древнейшие пресмыкающиеся, относящиеся к отрядукотилозавров, или целъночерепных (Cotylosauria), из подкласса анапсид, известны уже из верхних каменноугольных отложений, но только в пермский период они достигли значительного развития, а в триасе уже вымерли. Котилозавры были массивными животными на толстых пятипалых ногах и имели длину тела от нескольких десятков сантиметров до нескольких метров. Череп был покрыт сплошным панцирем из кожных костей с отверстиями лишь для ноздрей, глаз и теменного органа. Такое строение черепа, а также многие другие признаки указывают на чрезвычайную близость котилозавров к примитивным стегоцефалам, которые, несомненно, были их предками.
Наиболее примитивной из до сих пор известных анапсид, а следовательно, и рептилий вообще является нижнепермская сеймурия (Seymouria). Это сравнительно небольшое (до 0,5 м длины) пресмыкающееся обладало целым рядом признаков, характерных для земноводных: шея была почти не выражена, длинные острые зубы еще сохраняли примитивное строение, имелся лишь один крестцовый позвонок, а кости черепа обнаруживали замечательное сходство даже в деталях с черепной крышкой стегоцефалов.
Ископаемые остатки сеймуриоморфных рептилий, найденные в СССР (Kotlasia и др.), дали возможность советским палеонтологам определить их систематическое положение как представителей особого подклассабатрахозавров (Batrachosauria), занимающего промежуточное положение между земноводными и котилозаврами.
Котилозавры - весьма разнообразная группа. Самыми крупными представителями ее являются неуклюжие растительноядные парейазавры (Pareiasaurus), достигавшие 2-3 м длины. Позднее скелеты их были найдены в Южной Африке и у нас на Северной Двине.
Котилозавры были исходной группой, давшей начало всем прочим основным группам пресмыкающихся. Эволюция в основном протекала по пути возникновения более подвижных форм: конечности стали удлиняться, в образовании крестца приняло участие не менее двух позвонков, весь скелет, сохраняя свою прочность, стал легче, в частности первоначально сплошной костный панцирь черепа стал редуцироваться путем возникновения височных ям, что не только облегчило череп, но, что особенно важно, способствовало усилению мышц, сжимающих челюсти, так как, если в костной пластинке, к которой прикрепляются мускулы, образуется отверстие, мышца при своем сокращении может несколько выпячиваться в это отверстие. Редукция черепного панциря пошла двумя основными путями: путем образования одной височной ямы, ограниченной снизу скуловой дугой, и путем образования двух височных ям, в результате чего образовались две скуловые дуги. Таким образом, всех пресмыкающихся можно разделить на три группы: 1) анапсиды - с цельным черепным панцирем (котилозавры и черепахи); 2) синапсиды - с одной скуловой дугой (звероподобные, плезиозавры и, возможно, ихтиозавры) и 3) диапсиды - с двумя дугами (все прочие пресмыкающиеся). Первая и вторая группы содержат по одному подклассу, последняя распадается на ряд подклассов и множество отрядов.
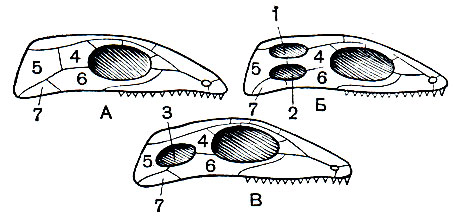
Рис. 79. Схема редукции черепного панциря у пресмыкающихся: А - стегальный тип черепа, лишенный височных ям, свойственный анапсидам; Б - тип черепа с двумя височными ямами, свойственный диапсидам; В - тип черепа с одной височной ямой, свойственный синапсидам: 1 - верхняя височная яма; 2 - боковая височная яма; 3 - височная яма; 4 - заглазничная кость; 5 - чешуйчатая кость; 6 - скуловая кость; 7 - квадратноскуловая кость
Группа анапсид является древнейшей ветвью рептилий, имеющих по строению черепа много общих черт с ископаемыми стегоцефалами, так как не только многие ранние формы их (котилозавры), но даже некоторые современные (некоторые черепахи) имеют сплошной черепной панцирь.
Черепахи являются единственными ныне живущими представителями этой древнейшей группы рептилий. Они отделились, по-видимому, непосредственно от котилозавров. Уже в триасе эта древняя группа вполне сложилась и благодаря своей крайней специализации дожила до настоящего времени, почти не изменившись, хотя в процессе эволюции некоторые группы черепах по нескольку раз переходили от наземного образа жизни к водному, в связи с чем они то почти утрачивали костные щиты, то вновь приобретали их.
От группыкотилозавров отделились морские ископаемые рептилии - ихтиозавры и плезиозавры, вместе с другими более редкими формами составившие два самостоятельных подкласса: Ихтиоптеригии (Ichthyopterygia) и Синаптозавры (Synaptosauria).
Плезиозавры (Plesiosauria), относящиеся к синаптозаврам, были морскими пресмыкающимися. Они имели широкое, бочонкообразное, уплощенное туловище, две пары мощных конечностей, видоизмененных в плавательные ласты, очень длинную шею, заканчивающуюся маленькой головкой, и короткий хвост. Кожа была голая. Многочисленные острые зубы сидели в отдельных ячейках. Размеры этих животных варьировали в очень широких пределах: некоторые виды имели лишь полметра длины, но были и гиганты, достигавшие 15 м. Характерная особенность их скелета заключалась в недоразвитии спинных частей поясов конечностей (лопатки, подвздошной кости) и исключительной мощности брюшных отделов поясов (коракоида, брюшного отростка лопатки, лобковой и седалищной костей), а также брюшных ребер. Все это свидетельствует об исключительно сильном развитии мышц, приводящих в движение ласты, которые служили лишь для гребли и не могли поддерживать тело вне воды.
Хотя в пределах подклассасинаптозавров переход от наземных форм к водным восстановлен довольно отчетливо, но происхождение группы в целом во многом еще неясно.
В то время как плезиозавры, приспособившись к водной жизни, еще сохранили облик наземных животных, ихтиозавры (Ichthyosauria), относящиеся к ихтиоптеригиям, приобрели сходство с рыбами и дельфинами. Тело ихтиозавров было веретенообразное, шея не выражена, голова удлиненная, хвост с большим плавником, конечности в виде коротких ластов, причем задние значительно меньше передних. Кожа была голая, многочисленные острые зубы (приспособленные к питанию рыбой) сидели в общей борозде, имелась лишь одна скуловая дуга, но крайне своеобразного строения. Размеры варьировали от 1 до 13 м.
Группа диапсид включает два подкласса: лепидозавров и архозавров.
Наиболее раннюю (верхняя пермь) и наиболее примитивную группу лепидозавров составляет отрядэозухий (Eosuchia). Они еще очень плохо изучены, лучше других известна Iounginia - мелкое пресмыкающееся, напоминающее телосложением ящерицу, со сравнительно слабыми конечностями, имевшими обычное рептильное строение. Примитивные черты ее выражаются главным образом в строении черепа, зубы расположены как на челюстях, так и на нёбе.
Первые клювоголовые (Rhynchocephalia) известны с раннего триаса. Некоторые из них были чрезвычайно близки к современной гаттерии. Отличаются клювоголовые от эозухий присутствием рогового клюва и тем, что зубы их приращены к кости, тогда как челюстные зубы эозухий сидели в отдельных ячейках. По последнему признаку клювоголовые даже примитивнее эозухий и, следовательно, должны были произойти от каких-то еще не найденных примитивных форм последней группы.
Чешуйчатые (Squamata), именно ящерицы, известны лишь с самого конца юры. От основного ствола чешуйчатых - ящериц - уже в начале мела, видимо, отделились мозазавры (Mosasauria). Это были морские пресмыкающиеся, имевшие длинное змеевидное тело и две пары конечностей, видоизмененных в ласты. Некоторые представители этого отряда достигали длины 15 м. В конце мела они бесследно вымерли. Несколько позже мозазавров (конец мела) от ящериц отделилась новая ветвь - змеи.
По всей вероятности, от эозухий же берет начало большая прогрессивная ветвь архозавров (Archosauria) - именно псевдозухии, распавшиеся впоследствии на три основные ветви - водную (крокодилы), наземную (динозавры) и воздушную (крылатые ящеры). Наряду с двумя типичными височными дугами наиболее характерной чертой этой группы была тенденция перехода к "двуногости", т. е. передвижению на одних задних конечностях. Правда, некоторые из наиболее примитивных архозавров только начали изменяться в этом направлении, и потомки их пошли по другому пути, а представители ряда групп вторично вернулись к передвижению на четырех конечностях. Но в последнем случае прошлая история оставила след на строении их таза и самих задних конечностей.
Псевдозухии (Pseudosuchia) впервые появились лишь в начале триаса. Ранние формы были небольшими животными, но уже с относительно длинными задними ногами, которые, по-видимому, одни и служили им для передвижения. Зубы, имевшиеся только на челюстях, сидели в отдельных ячейках, вдоль спины почти всегда располагались в несколько рядов костные пластинки. Эти мелкие формы, типичными представителями которых являются орнитозухи, и ведущие, по-видимому, древесную жизнь Scleromochlus были очень многочисленны и дали начало не только ветвям, получившим расцвет позже - в юре и мелу, но и ряду сильно специализированных групп, бесследно вымерших еще в триасе.
Наконец, псевдозухии, в частности если не сам орнитозух, то формы, близкие к нему, могли быть предками птиц.
Крокодилы (Crocodylia) очень близки к некоторым триасовым псевдозухиям, например белодону (Belodon, или Phytosaurus). Начиная с юры появляются уже настоящие крокодилы, но окончательно выработался современный тип крокодилов лишь в течение мелового периода. На этом длинном пути эволюции можно шаг за шагом проследить, как развивался характерный признак крокодилов - вторичное нёбо. Сперва появились лишь горизонтальные отростки на верхнечелюстных и нёбных костях, затем эти нёбные отростки сошлись, а еще позже к ним присоединились нёбные отростки крыловидных костей, и одновременно с этим процессом шло передвижение ноздрей вперед, а вторичных хоан - назад.
Динозавры (Dinosauria) - наиболее многочисленная и разнообразная группа пресмыкающихся, когда-либо жившая на свете. Сюда относились и небольшие формы, размером с кошку и мельче, и гиганты, достигавшие почти 30 м длины и 40-50 т веса, легкие и массивные, подвижные и неуклюжие, хищные и травоядные, лишенные чешуй и покрытые костным панцирем с различными выростами. Многие из них бегали скачками на одних задних конечностях, опираясь на хвост, другие передвигались на всех четырех. Голова у динозавров была обычно сравнительно мала, полость же черепной коробки - совсем крошечная. Зато спинномозговой канал в области крестца был очень широк, что указывает на местное расширение спинного мозга.
Динозавры разделялись на две большие группы - ящеротазовых и птицетазовых, которые возникли совершенно самостоятельно от псевдозухий. Различия их заключаются главным образом в строении пояса задних конечностей.
Ящеротазовые (Saurischia), родственные связи которых с псевдозухиями не вызывают сомнения, первоначально были только хищными. В дальнейшем, хотя большинство форм продолжало оставаться хищными, часть превратилась в растительноядных. Хищные, хотя и достигали огромных размеров (до 10 м длины), имели сравнительно легкое телосложение и мощный череп с острыми зубами. Передние конечности их, служившие, по-видимому, лишь для схватывания добычи, были сильно уменьшены, и животное должно было передвигаться, прыгая на задних конечностях и опираясь на хвост. Типичный представитель таких форм - цератозавр (Ceratosaurus).
В противоположность хищным растительноядные формы передвигались на обеих парах конечностей, которые были у них почти равной длины и оканчивались пятью пальцами, по-видимому, покрытыми роговыми образованиями вроде копыт. К ним относились самые крупные четвероногие животные, когда-либо жившие на земном шаре, например бронтозавр, достигавший свыше 20 м длины и, вероятно, 30 т веса, и диплодок. Последний был стройнее и, несомненно, значительно легче, но зато превосходил бронтозавра длиной, которая у одного экземпляра превышала 26 м; наконец, неуклюжий брахиозавр, длиной около 24 м, должно быть, весил около 50 т. Хотя полые кости облегчали вес этих животных, все же трудно допустить, чтобы такие гиганты могли свободно двигаться по суше. По-видимому, они вели лишь полуназемную жизнь и, подобно современным бегемотам, большую часть времени проводили в воде. На это указывают и их очень слабые зубы, пригодные для поедания лишь мягкой водной растительности, и то, что, например, у диплодока ноздри и глаза были сдвинуты кверху, так что животное могло видеть и дышать, выставив из воды только часть головы.
Птицетазовые (Ornithischia), имевшие пояс задних конечностей, чрезвычайно похожий на птичий, никогда не достигали таких огромных размеров. Зато они были еще разнообразнее. Большинство этих животных вторично вернулось к передвижению на четырех ногах и обычно имело хорошо развитый панцирь, иногда осложненный различного рода выростами в виде рогов, шипов и т. д. Все они с самого начала и до конца оставались растительноядными, и большинство сохранило лишь задние зубы, тогда как передняя часть челюстей была, по-видимому, покрыта роговым клювом.
Как характерных представителей различных групп птицетазовых можно указать игуанодонов, стегозавров и трицератопсов.
Игуанодоны (Iguanodon), достигавшие 5-9 м высоты, бегали на одних задних ногах и были лишены панциря, но зато 1-й палец передних конечностей у них представлял собой костный шип, который мог служить хорошим орудием защиты.
Стегозавр имел крошечную голову, двойной ряд высоких треугольных костных пластин на спине, и несколько острых шипов сидело на хвосте.
Трицератопс (Triceratops) внешне походил на носорога: на конце его морды располагался большой рог, кроме того, пара рогов возвышалась над глазами, а по заднему, расширенному краю черепа сидели многочисленные заостренные отростки.
Птеродактили (Pterosauria), подобно птицам и летучим мышам, были настоящими летающими животными. Их передние конечности представляли собой настоящие крылья, но крайне своеобразного устройства: не только предплечье, но и сращенные друг с другом пястные кости были сильно удлинены, первые три пальца имели нормальное строение и размеры, пятый отсутствовал, тогда как четвертый достигал чрезвычайной длины и между ним и боками тела была натянута тонкая летательная перепонка. Челюсти были выдвинуты, у некоторых форм имелись зубы, у других - беззубый клюв. Птеродактили обнаруживают ряд общих черт с птицами: сращенные грудные позвонки, большую грудину с килем, сложный крестец, полые кости, лишенную швов черепную коробку, большие глаза. Питались крылатые ящеры, по-видимому, рыбой и жили, вероятно, по прибрежным скалам, так как, судя по строению задних конечностей, подняться с ровной поверхности они не могли. К птеродактилям относят довольно разнообразные формы: сравнительно примитивную группу рамфоринхов, имевших длинный хвост, и собственно птеродактилей с зачаточным хвостом. Размеры колебались от величины воробья до гигантского птеранодона, размах крыльев которого достигал 7 м.
Группа синапсид составляет самостоятельный подкласс рептилий, как особая боковая ветвь, отделившаяся от древних котилозавров. Они характеризуются усилением челюстного аппарата путем образования своеобразной височной впадины для очень мощной челюстной мускулатуры и прогрессивной дифференцировкой зубной системы - разнозубостью, или гетеродонтностью. Это связывает их с высшим классом позвоночных - млекопитающими.
Звероподобные (Theromorpha) - это та группа, примитивные представители которой были еще очень близки к котилозаврам. Отличие их заключается главным образом в присутствии скуловой дуги и более легком телосложении. Звероподобные появились в конце каменноугольного периода, а начиная с нижней перми стали очень многочисленными и в течение всего этого периода вместе с котилозаврами были почти единственными представителями своего класса. Несмотря на все свое разнообразие, все звероподобные были строго наземными животными, передвигающимися исключительно при помощи обеих пар конечностей.
Наиболее примитивные представители пеликозавров (например, Varanops) имели небольшие размеры и внешне должны были напоминать ящериц. Однако зубы их, хотя и однородные, уже сидели в отдельных ячейках.
Зверообразные (Therapsida), сменившие пеликозавров со средней перми, объединяли чрезвычайно разнообразных животных, из которых многие были узкоспециализированы. У - более поздних форм теменное отверстие исчезло, зубы дифференцировались на резцы, клыки и коренные, образовалось вторичное нёбо, один мыщелок разделился на два,, зубная кость сильно увеличилась, прочие же кости нижней челюсти уменьшились.
Причины вымирания древних рептилий до сих пор не вполне ясны. Наиболее правдоподобное объяснение этого явления следующее. В процессе борьбы за существование отдельные формы все более приспосабливались к определенным условиям среды, все более специализировались. Такая специализация крайне полезна, но лишь до тех пор, пока продолжают существовать те условия, к которым приспособился организм. Стоит же им измениться - такие животные оказываются в худших условиях, чем менее специализированные формы, которые их и вытесняют в борьбе за существование. Кроме того, в борьбе за существование некоторые группы могут приобретать свойства, повышающие их общую жизнедеятельность. В противоположность узкой адаптации, или идиоадаптации, это явление получило название ароморфоза. Например, теплокровность дала возможность организмам, приобретшим это свойство, меньше зависеть от климата по сравнению с животными с переменной температурой тела. В течение долгой мезозойской эры происходили лишь незначительные изменения ландшафтов и климата, в связи с чем пресмыкающиеся все более и более специализировались и процветали. Но в конце этой эры земная поверхность стала претерпевать такие огромные горообразовательные процессы и связанные с ними климатические изменения, что большинство пресмыкающихся не смогло их пережить и бесследно вымерло к концу мезозоя, получившего название эпохи великого вымирания. Однако было бы ошибкой объяснить этот процесс исключительно физико-географическими причинами. Не меньшую роль сыграла и борьба за существование с другими животными, именно с птицами и млекопитающими, которые благодаря теплокровности и высокоразвитому мозгу оказались лучше приспособленными к этим внешним явлениям и вышли победителями в жизненной борьбе.
Подкласс Анапеиды (Anapsida) + Отряд Сеймуриоморфы (Seymouriomorpha) + Отряд Котилозавры (Cotylosauria) Отряд Черепахи (Testudines, или Chelonia) + Подкласс Ихтиоптеригии (Ichthyopterygia) + Отряд Мезозавры (Mesosauria) + Отряд Ихтиозавры (Ichtyosauria) + Подкласс Синаптозавры (Synaptosauria) + Отряд Проторозавры (Protorosauria) + Отряд Плезиозавры (Sauropterygia) +Подкласс Лепидозавры (Lepidosauria) + Отряд Эозухии (Eosuchia) Отряд Клювоголовые (Rhynchocephalia) Отряд Чешуйчатые (Squamata) + Подкласс Архозавры (Archosauria) + Отряд Текодонты (Thecodontia) Отряд Крокодилы (Crocodylia) + Отряд Птерозавры (Pterosauria) + Отряд Ящеротазовые динозавры (Saurischia) + Отряд Птицетазовые динозавры (Ornithischia) + Подкласс Синапсиды (Synapsida) + Отряд Пеликозавры (Pelycosauria) + Отряд Терапсиды (Therapsida)
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'