Отряд Бесхвостые земноводные (Anura) (М. Н. Денисова)
Бесхвостые земноводные представляют собой наиболее высокоорганизованный и богатый представителями отряд. Разнообразие видов бесхвостых амфибий в 6,5 раза больше, чем хвостатых, и в 32 раза больше, чем безногих. Ныне живущих бесхвостых насчитывается около 1800 видов. Однако, несмотря на свою многочисленность, все они имеют довольно однообразное строение. Туловище у них короткое, шея не выражена, хвоста нет, парные конечности хорошо развиты, причем задние конечности в два-три раза больше передних и служат для характерного передвижения прыжками.
Однообразие в строении бесхвостых связано с приспособлением к передвижению прыжками, что первично было присуще всем представителям этого отряда. Одновременный толчок задними ногами повлек за собой выработку укороченного, клинообразного тела и удлиненных задних конечностей. При этом возник дополнительный отдел - предплюсна как эластичная вставка, продолжающая действие мускулатуры при прыжке (рис. 46). Образование узкого таза в виде характерного диска есть также приспособление к наиболее полному использованию силы при прыжке, поскольку при таком типе движения целесообразно иметь точки приложения сил максимально сдвинутыми. С этим же связано характерное удлинение подвздошных костей, способствующих перенесению точек приложения сил к центру тяжести животного.
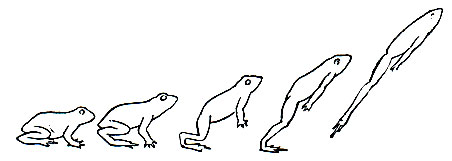
Рис. 46. Шесть положений лягушки во время прыжка
Грудина состоит из нескольких частей. В плечевом поясе имеется ключица. Хвостовые позвонки срослись в одну длинную косточку - уростиль. Лобные и теменные кости слиты в парную лобнотеменную кость. Жаберные дуги изменены в подъязычный аппарат"
Кроме внутреннего, у бесхвостых есть еще и среднее ухо с барабанной перепонкой, иногда скрытой под кожей. Ни жабры, ни жаберные щели у взрослых никогда не сохраняются. Кровеносная система характеризуется отсутствием связи между дугой аорты и сонными и легочными артериями и исчезновением третьей артериальной дуги.
Ныне живущих бесхвостых амфибий объединяют в 176 родов, 12 семейств и 5 подотрядов. В основу деления на подотряды и их наименования положены особенности сочленения позвонков в туловище.
Подотряд Amphicoela содержит наиболее примитивных бесхвостых земноводных, имеющих двояковогнутые (амфицельные) позвонки; у них сохраняются рудименты хвоста, имеющие значение при размножении (Ascaphus, Северная Америка), или рудименты хвостовых мышц (Liopelma, Новая Зеландия).
Представители подотряда Opisthocoela характеризуются задневогнутыми (опистоцельными) позвонками, отсутствием свободного выбрасывающегося языка и наличием ребер. Сюда относятся жерлянки (Bombina), жабы-повитухи (Alytes), африканские шпорцевые лягушки (Xenopus) и американские пипы (Pipa).
Подотряд Anomocoela объединяет чесночниц (Pelobates), крестовок (Pelodytes) и ряд экзотических форм. Представители отряда характеризуются передневогнутыми (процельными) позвонками или процельными позвонками со свободными вставочными дисками. Крестцовый позвонок слит или сочленяется с уростилем одним бугорком. Свободных ребер нет.
Подотряд Procoela включает жаб (Bufonidae), квакш (Hylidae) и короткоголовых (Brachycephalidae). Для видов этого подотряда характерны передневогнутые (процельные) позвонки и двойной сочленовный бугорок между крестцом и уростилем. Ребер нет.
Подотряд Diplasiocoela объединяет наибольшее число видов; сюда относятся настоящие лягушки (Ranidae), узкоротые (Brevicipitidae) и веслоногие лягушки (Polypedatidae). Первые 7 туловищных позвонков у них процельные, последний - амфицельный или реже все процельные, так же как и крестцовый. Ребер нет.
Бесхвостые земноводные - главным образом сухопутные формы. Число вторично вернувшихся к водному образу жизни по сравнению с хвостатыми очень невелико и не превышает 15%; лишь одно небольшое семейство пиповых (Pipidae) целиком состоит из постоянно живущих в воде видов. У некоторых водных форм редуцирована барабанная перепонка (жерлянки) и язык (пиповые). У наземных форм язык, выбрасывающийся наружу, играет важную роль в захватывании пищи. У водных форм эта функция языка выпадает. Видимо, в связи с этим он и редуцируется. У африканской шпорцевой лягушки (Xenopus) редуцированы и веки и сохраняется орган боковой линии.
Среди сухопутных бесхвостых есть роющие и древесные.
Древесные формы встречаются среди бесхвостых весьма часто. В 6 семействах из 12 есть древесные виды, а одно из самых больших семейств - квакши (Hylidae) - более чем на 90% состоит из древесных форм; многие виды лягушек и жаб ведут также древесный образ жизни; у живущих на деревьях видов, как, например, у квакши обыкновенной (Hyla arborea), концы пальцев расширены в диски. На этих дисках образуется эпидермальная щетка и особые железы, выделяющие липкий секрет, с помощью которого животное удерживается на вертикальных поверхностях. У некоторых древесных лягушек Южной Америки и Юго-Восточной Азии чрезвычайно развиты перепонки между удлиненными пальцами передних и задних лап. Среди таких лягушек особенно известны веслоногие лягушки (Polypedates, или Rhacophorus) из Юго-Восточной Азии. Некоторые из них, прыгая за насекомыми, пролетают планирующим полетом до 12 м и более с дерева на дерево.
Роющие формы, так же как и древесные, весьма многочисленны в отряде бесхвостых. Они встречаются среди представителей шести семейств, и одно из них - семейство чесночниц (Pelobatidae) - состоит почти целиком из видов, ведущих роющий образ жизни. Роющие виды многочисленны среди настоящих жаб (Bufonidae) и узкоротых (Brevicipitidae) Африки и Австралии. Роют бесхвостые земноводные задними лапами, совершая попеременные движения ногами в стороны, так что погружаются в землю задним концом тела. Переход к роющему образу жизни повлек за собой укорочение конечностей, что лишило животных возможности совершать большие прыжки. Жабы и чесночницы чаще передвигаются шагом, чем прыгают. Чередующиеся движения правой и левой лап привели также к образованию подвижного сочленения в виде скользящих суставных поверхностей между подвздошными костями и крестцовыми позвонками при значительном расширении поперечных отростков последних. Наконец, появились острые роговые мозоли на основании первого пальца, принимающие, например у чесночниц, вид лопаты. С роющим образом жизни связаны кожные окостенения на голове, как у нашей чесночницы. Эти образования могут принимать причудливую форму костяного шлема.

Таблица 7. Квакша обыкновенная (Hyla arbrea)

Таблица 7а. Квакша изменчивая (Hyla versicolor)
Бесхвостые распространены очень широко по всем материкам (кроме Антарктиды) и по всем материковым островам, исключая самые северные.
Самые древние ископаемые остатки бесхвостых амфибий известны из нижнего триаса северной части Мадагаскара. Здесь найден единственный представитель особого отряда Proanura - Protobatrachus massinoti. У этого первичнобесхвостого, длина тела которого около 10 см, было 16 позвонков и все туловищные, за исключением первого, несли ребра. Между короткими подвздошными костями лежали 3 хвостовых позвонка. Локтевая и лучевая, а также большая и малая берцовые кости еще не были слиты.
Начиная с верхней юры и нижнего мела появляются уже типичные бесхвостые амфибии различных подотрядов.
Семейство Гладконогие (Liopelmidae)
К этому семейству относится два вида самых древних представителей из ныне живущих бесхвостых земноводных, каждый из которых выделяется в особый род. Это - гладконог, или хвостатая лягушка (Ascaphus truei), из Северной Америки и и лиопелъма (Liopelma hochstetteri), живущая в водоемах Новой Зеландии. Они обладают некоторыми чертами строения, свойственными хвостатым земноводным. Например, позвонки у них двояковогнутые (амфицельные); в течение всей жизни у них сохраняются ребра и в тазовом поясе имеется предлобковый хрящ; предгрудины нет, а небольшая грудина целиком хрящевая.
Замечательная внешняя особенность хвостатой лягушки, или гладконога (рис. 47), состоит в том, что самец имеет хвост. В отличие от саламандр и тритонов хвост у гладконога не имеет скелета и сухожилий, но снабжен двумя мускулами, имеющимися и у представителей хвостатых. У самки хвоста нет, а есть лишь короткая, малозаметная анальная трубка. Максимальная длина тела этого животного - 50 мм, но среди взрослых встречаются особи, достигающие всего лишь 30 мм. Кожа у хвостатой лягушки может быть гладкой или покрытой редко разбросанными бородавками. Бурая поверхность тела неравномерно покрыта черными пятнами, по бокам головы помещаются бурые продолговатые околоушные железы. Между глазами зеленая или желтая полоса, морда зеленоватая или рыжевато-коричневая. Низ тела желтовато-белый, границы его с более темным или малиновым оттенком. Соответственно господствующей окраске горных пород в местах обитания животного окраска его может быть темнее или светлее. Большие глаза отличаются вертикальным зрачком. Пальцы передних конечностей, длинные и тонкие, совершенно свободны от перепонок. На задних конечностях пальцы соединены перепонкой только в основании.
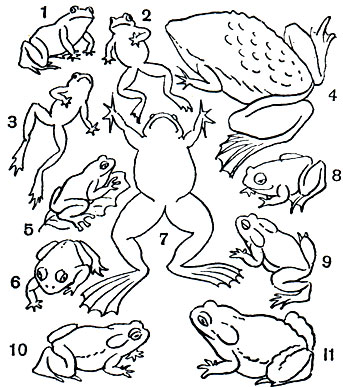
Таблица 8. Бесхвостые земноводные: 1 - жерлянка дальневосточная (Bombina orientalis); 2 - жерлянка краснобрюхая (Bombina bombina); 3 - жерлянка желтобрюхая (Bombina variegata); 4 - пипа суринамская (Pipa pipa); 5 - квакша обыкновенная (Hyla arborea); 6 - лопатоног скафиопус (Scaphiopus); 7 - шпорцевая лягушка (Xenopus laevis); 8 - чесночница обыкновенная (Pelobates fuscus); 9 - жаба камышовая (Bufo calamita); 10 - жаба зеленая (Bufo viridis); 11 - жаба серая (Bufo bufo)

Таблица 8. Бесхвостые земноводные: 1 - жерлянка дальневосточная (Bombina orientalis); 2 - жерлянка краснобрюхая (Bombina bombina); 3 - жерлянка желтобрюхая (Bombina variegata); 4 - пипа суринамская (Pipa pipa); 5 - квакша обыкновенная (Hyla arborea); 6 - лопатоног скафиопус (Scaphiopus); 7 - шпорцевая лягушка (Xenopus laevis); 8 - чесночница обыкновенная (Pelobates fuscus); 9 - жаба камышовая (Bufo calamita); 10 - жаба зеленая (Bufo viridis); 11 - жаба серая (Bufo bufo)
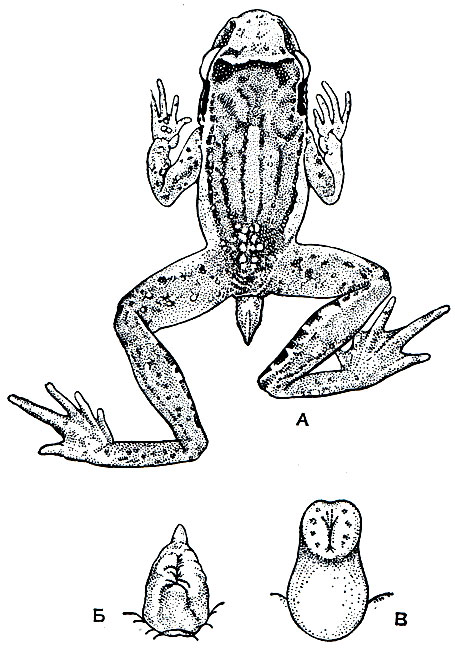
Рис. 47. Самец хвостатой лягушки (Ascaphus truei) с клоакальным придатком: А - общий вид; Б - придаток с брюшной стороны; В - он же, полностью растянутый, видны внутренние шипики
Хвостатая лягушка встречается в США и Канаде; ареал ее, шириной примерно в 160 км, расположен вдоль морского побережья штатов Вашингтон, Орегон, Калифорния, по южной границе Британской Колумбии и в штате Монтана.
Места обитания - лесные районы с быстротекущими горными ручьями, отличающимися очень холодной водой. Большую часть жизни хвостатые лягушки проводят в ручьях, прячась под камнями. Держатся они поодиночке. Изредка после периода сильных дождей животные покидают ручей и выходят на сушу, в лес. Как далеко они отходят от ручьев и сколько времени могут оставаться на суше, неизвестно.
Хвостатые лягушки - хороший образец приспособления к жизни в горных ручьях. Легкие их небольшие, а кожа богата кровеносными сосудами и является основным органом дыхания. В быстро текущих шумных ручьях звуки почти не слышны. В связи с этим самцы хвостатых лягушек не устраивают "брачных концертов". Соответственно у обоих полов совершенно отсутствует среднее ухо и евстахиевы трубы. Удивительным приспособлением к жизни в быстро текущих водах у этого вида оказывается внутреннее оплодотворение. Хвост - это своеобразный совокупительный орган, по которому семя стекает в клоаку самки.
Не имея возможности издавать призывные крики, самец активно разыскивает самку, ползая по дну ручья. Встретившись с нею, он обхватывает ее впереди задних лап, в паховой области. В связи с более активным передвижением и обхватыванием самки передние и задние конечности у самцов длиннее и толще, чем у самок. Брачные мозоли самцов, шершавые наросты на передних лапах, помогающие удерживать самку, располагаются у хвостатых лягушек на внутренней стороне первого пальца, на предплечье и у некоторых даже на плече. Вторичные половые признаки бывают резко выражены лишь во время спаривания.
Яйца хвостатой лягушки крупны и достигают 5 мм в диаметре, а с оболочкой - 8 мм. Они лишены пигмента. Самка откладывает их в ручьи комком, прикрепляя к нижней стороне камней. Кладка невелика, не более 50 яиц. Это свидетельствует о высокой выживаемости потомства, чему в первую очередь способствует внутреннее оплодотворение.
Контуры тела головастиков, если смотреть сверху, имеют клиновидную форму, хвост у них длинный, хвостовой гребень низкий и никогда не простирается на тело, как это бывает у многих головастиков, живущих в стоячих водоемах. Дыхательное отверстие находится на нижней стороне тела, около заднепроходного отверстия. Окраска головастиков темная, испещренная пятнами. Хвост по цвету не отличается от тела; кончик его белый. Рот очень большой, круглый; на верхней губе помещаются 2-3, на нижней 7-10 непрерывных рядов губных зубов. С помощью больших присасывающихся губ головастики прикрепляются к камням и соскабливают с их поверхности водоросли. Большую часть личиночной жизни они и проводят таким образом, предпочитая валуны, лежащие на самом быстром течении. Головастики могут плавать даже против течения, но недолго. В особенно быстро текущих ручьях у них хвосты длиннее, а ротовые диски уже.
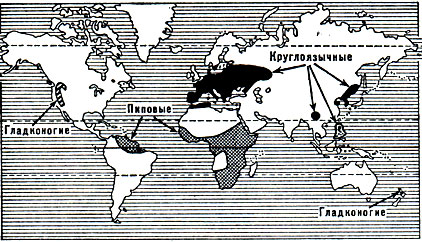
Рис. 48. Области распространения семейств гладконогих, круглоязычных, пиповых
Период размножения у хвостатой лягушки длится с мая до октября. Икра развивается около месяца. С момента вылупления головастиков до конца метаморфоза проходит около года. На севере ареала, в Канаде, где вода холоднее, были обнаружены двухгодовалые головастики. В горах Северного Айдахо спаривание хвостатых лягушек наблюдали только в сентябре. Икра выметывалась лишь на следующий год. Головастики вылуплялись в августе - сентябре и достигали длины 13-15 мм. Все личиночное развитие продолжается здесь 3 года, собственно метаморфоз - 60 дней. К этому времени головастики вырастают до 50 мм. Возможно, что здесь половозрелые самки откладывают икру один раз в два года, но вдвое большую порцию, чем на Тихоокеанском побережье.
Единственный вид другого рода этого семейства - Leiopelma hochstetteri живет в водоемах Новой Зеландии и достигает всего лишь 33 мм длины.
Семейство Круглоязычные (Discoglossidae)
Это семейство объединяет древних, примитивных бесхвостых земноводных, населяющих Европу и Азию. Оно включает 8 видов, принадлежащих к 4 родам.
К примитивным особенностям строения представителей этого семейства относится наличие коротких ребер и вогнутые с задней поверхности позвонки, что приближает круглоязычных к высшим хвостатым земноводным. У них толстый дискообразный язык, прикрепленный всей своей нижней поверхностью и не принимающий участия в схватывании добычи.
К роду жерлянок (Bombina) относится 4 вида. Для них характерно отсутствие барабанной перепонки, трехгранный зрачок, бугорчатая кожа, красное или желтое брюшко с резкими черными пятнами.
Краснобрюхая жерлянка (Bombina bombina) сверху светло-серого, буроватого или черного цвета с темными, реже зелеными пятнами. Брюшко ярко-оранжевое с синевато-черными пятнами. Кожа на брюшке и на спине бугорчатая. Концы пальцев, если смотреть сверху, темные. У самцов есть внутренние резонаторы. Размеры краснобрюхой жерлянки изменяются от 41 (в Румынии) до 60 мм (в Курской области). По длине самки больше самцов, а по весу равны им. В брачное время самцы имеют черные мозоли на первом и втором пальцах передней конечности и на внутренней части предплечья.
Распространена в Центральной и Восточной Европе: на востоке - до Урала, на юге - до Дуная, на севере - до Южной Швеции. Селится на равнинах, избегая гор. Живет в воде, чаще всего в болотах, озерах, старицах, канавах, лужах речных пойм, на рисовых полях. Предпочитает мелкие, хорошо прогреваемые водоемы, покрытые ряской, с глинистым дном. Песчаных берегов и быстрого течения жерлянка не любит. Отсутствует в солоноватых озерах.
В течение весенне-летнего сезона жерлянки могут совершать перемещения из одного водоема, в котором они кормятся, в другой на расстояние до 700 м. Один из водоемов, отличающийся постоянством уровня воды, становится для половозрелых животных местом нереста. Скорость передвижения жерлянок в среднем 33 м в сутки.
Жерлянки активны при температуре воды от 10 до 30°. Наибольшая активность наблюдается при температуре от 16 до 23°. Предпочитаемая температура, установленная экспериментально, оказалась равной 21°.
Обычно краснобрюхая жерлянка не бывает особенно многочисленной и составляет лишь небольшую долю от общего количества бесхвостых амфибий той или иной местности. Так, в Беловежской пуще на ее долю приходилось 0,4%, а в Волжско-Камском заповеднике - 0,7%.
В период размножения, когда происходит концентрация взрослых животных в нерестовых водоемах, плотность поселения жерлянок резко возрастает. Так, в Волжско-Камском заповеднике на 1 га водного зеркала насчитывали до 74 экземпляров (28% от общего количества земноводных в водоеме). После окончания икрометания, когда жерлянки рассеиваются по другим водоемам, плотность их поселения снижается почти в два раза, и в это время они составляют около 12% от общего количества водных амфибий.
В связи с вырубанием лесов краснобрюхая жерлянка, вероятно, продвигается к северу, увеличиваясь в некоторых местах в числе. В 40-х годах нашего века в десятки раз увеличилась численность этого вида под Москвой.
Индивидуального участка обитания краснобрюхие жерлянки не имеют. В какой-то степени можно говорить об участке обитания популяции, включающем места зимовки, нерестовые и кормовые водоемы. В одном и том же водоеме известны встречи одних и тех же меченых жерлянок в течение 2-4 сезонов подряд.
Жерлянка кормится преимущественно в дневные часы. Ее "брачные песни" слышатся также на протяжении всего дня, заметно стихая ночью. Однако на протяжении светлого времени суток наблюдается три пика активности: с 11 до 12, с 16 до 17 и самый большой с 19 до 20 часов. По мере изменения продолжительности дня часы максимальной активности жерлянок несколько сдвигаются. Миграции происходят ночью при 92-100% относительной влажности припочвенного слоя воздуха. Сеголетки, выходящие осенью из водоемов, совершают передвижения и днем.
Обитая всю жизнь в водоемах, жерлянки в значительной мере живут за счет водных беспозвоночных, питаясь ими в большей степени, чем зеленые лягушки, часто живущие сними бок о бок. Так, в средней полосе нашей страны у прудовой лягушки водные корма составляют 17%, у озерной - 23% , а у жерлянки - 43%. В Предкавказье у озерной лягушки водные корма составляют 12% , а у жерлянки - 76%. Однако в разных местах пища жерлянки не остается однотипной. В Волжско-Камском крае и в Казахстане водных животных жерлянка поедает меньше - от 11 до 30%.
В Словакии и в Предкавказье в питании жерлянки преобладают двукрылые, главным образом их личинки; в Волжско-Камском крае и в Казахстане - жуки, а в Воронежском заповеднике - дождевые черви.
В средней полосе в связи со значительной ролью в питании жерлянки как наземных животных, так и водных набор кормов у нее разнообразный и суммарный процент преобладающих компонентов невелик (50%). В Предкавказье, где водные животные играют ведущую роль, специализация в питании возрастает и доля доминирующих кормов оказывается уже равной 88%. Летающие формы представлены в питании этого вида незначительно (8%).
Состав кормов у сеголеток тот же, что и у взрослых, но преобладают более мелкие животные. Наполненность желудка жерлянки не превышает 5 - 7% от веса тела, а в осеннее время снижается до 0,7%.
На зимовку из водоемов жерлянки уходят в Белгородской и Курской областях в третьей декаде ноября, в Волжско-Камском заповеднике - на месяц раньше. Зимуют на суше в норах полевок, водяной крысы, выхухоли, в песчаных ямах, в рыхлой наносной почве по берегам водоемов, под жилыми постройками и в погребах. В окрестностях Владимира, Горького, в Смоленской области, под Киевом и в Казахстане известны зимовки в водоемах. В Волжско-Камском заповеднике жерлянки были найдены в ямах на глубине 40-60 см вместе с жабами и тритонами. Над ямой располагался слой листьев толщиной до 40 см и слой снега 83 см.
Продолжительность спячки у жерлянки невелика, около 150 дней. Выход с зимовки в разных местах происходит во второй половине марта - в конце апреля.
Спустя 15-20 дней после выхода с зимовок у жерлянок наступает период размножения. Разгар тока бывает после того, как сойдут талые воды и зазеленеют луга. Голос - короткое, минорное "ук" или "унк". Токующий самец лежит распластавшись на воде, время от времени передвигаясь толчками. При крике тело жерлянки вибрирует и по воде расходятся круги. Испуганная жерлянка ныряет, но может "укать" и под водой. Голоса их звучат особенно интенсивно во время спаривания.
Икрометание растянуто и происходит в течение всего лета. Однако если судить по интенсивности "пения", то в Средней Европе наблюдается две волны спаривания: в конце апреля и в середине мая. Между этими двумя пиками на протяжении примерно недели голоса жерлянок слышатся редко. Так же редко слышны они и после 20 мая. Изменение интенсивности пения жерлянок совпадает с ходом изменения температуры. Минимальная температура, при которой начинают слышаться их голоса, равна примерно 14°. Последние спаривающиеся жерлянки наблюдались в Волжско-Камском заповеднике в начале июля.
Самец во время спаривания охватывает самку впереди основания бедер. Соотношение полов в разгар нереста близко 1:1.
Икра откладывается ночью, порциями по 2-80 штук. Общее количество икринок, откладываемых одной самкой,- 80-100 (до 300). В яичниках жерлянки насчитывали до 500 икринок. Возможно, часть из них рассасывается. Диаметр яйца около 2 мм; диаметр икринки - 7-8 мм.
Для откладки икры жерлянка выбирает хорошо прогреваемые мелководные участки водоемов, поросшие растительностью, которая служит местом прикрепления икры. Прудовые и озерные лягушки, откладывающие икру одновременно с жерлянками, придерживаются более глубоких мест. Температура воды в период икрометания 16-21°.
Головастики вылупляются через 4-10 дней и имеют длину 3,5-5 мм. Через 3-4 недели длина их достигает 33-48 мм. Они отличаются широким хвостовым плавником, так как держатся не на мелководьях, а в толще воды и передвигаются на относительно большие расстояния. По некоторым данным, головастики жерлянок - хищники, но охотно питаются также и трупами животных. По другим наблюдениям, они питаются водорослями. Смертность яиц и личинок примерно 46%.
Развитие головастиков занимает около 3 месяцев. В Волжско-Камском крае сеголетки появляются в середине июля - начале августа. В это время их длина 11,5-24,6 мм и вес 0,4-1,4 г. Через 2-2,5 месяца после метаморфоза, к моменту выхода из водоемов на зимовку, размер молодых животных увеличивается на 0,8-2,7 мм, а вес - на 0,3 г.
Под Москвой сеголетки составляют в августе 96,9% популяции, годовалые - 2,1%, старшие возрасты - 1-3%. Основная масса сеголеток гибнет в период выхода на сушу и во время первой зимовки, которую переживают всего 2-6%. Годовалых жерлянок погибает примерно 40%, так что в природе больше двух лет живут лишь немногие жерлянки, В Волжско-Камском заповеднике среди повторно встреченных меченых жерлянок лишь одна была обнаружена на пятом году жизни. В неволе они жили до 29 лет.
Наибольшая гибель жерлянок наблюдается во время зимовок и при перемещениях к ним и обратно. Икра и головастики гибнут при пересыхании водоемов. Влияние этих факторов определяет и изменение численности жерлянок по годам. Воздействие хищников на этот вид, по всей вероятности, незначительно, так как кожные железы жерлянки выделяют пенистый секрет - фринолицин, который более ядовит, чем секрет других наших амфибий, в том числе обыкновенной жабы. Привозимые с экскурсий в одной посуде с жерлянками обыкновенные тритоны и остромордые лягушки обычно гибнут через несколько часов. Несмотря на ядовитые выделения, жерлянка все-таки становится добычей некоторых животных. Ее поедают прудовые лягушки, ужи, гадюки, малая выпь, рыжая и малая белая цапли, кваква, сарыч, малый подорлик, грач, сорокопут-жулан, еж и хорьки. Правда, во всех случаях жерлянки составляют незначительную долю в пищевом рационе этих животных и могут считаться вынужденной пищей. Только у кваквы встречаемость этого вида в желудках достигает 5-25%.
Потревоженная жерлянка принимает специфическую позу, поворачиваясь на спину, прогибая ее и выворачивая конечности так, что становится заметной оранжевая окраска нижней части тела. Эту окраску считают предупреждающей, отпугивающей хищников от ядовитого животного. Однако такую позу у краснобрюхой жерлянки не удается наблюдать часто. Она ярко выражена у желтобрюхой жерлянки.
Желтобрюхая жерлянка (Bombina variegata) обитает в Средней и Южной Европе на Апеннинском и Балканском полуостровах. На территории Советского Союза желтобрюхая жерлянка встречается в Ивано- Франковской, Львовской, Черновицкой и Закарпатской областях. Это ее восточная граница распространения. В горы поднимается до 1900 м, но особенно многочисленна на высоте 200-300 м над уровнем моря.
По ряду признаков ее считают более древней, чем краснобрюхая жерлянка. Внешне она отличается цветом брюшка, общий фон которого лимонно-желтый или оранжево-желтый. Концы пальцев как сверху, так и снизу желтые. Максимальная длина 50 мм.
Весь активный период желтобрюхая жерлянка проводит в воде или возле воды, в выборе водоемов очень неприхотлива. Изредка встречается в воде сероводородных источников, была обнаружена в дождевых лужах, расположенных среди глыб выходящей на поверхность земли каменной соли. Интересно, что в соленой воде держались не только взрослые жерлянки, но была отложена икра и наблюдались головастики разных возрастов. Малочувствительны жерлянки и к отходам нефти, загрязняющим воду.
Активны желтобрюхие жерлянки в светлые часы суток. В питании их преобладают наземные беспозвоночные, первое место среди которых занимают жуки, затем двукрылые, перепончатокрылые и дождевые черви.
На зимовку уходят в конце сентября - в октябре. Зимуют на суше в норах и под камнями. В ручейке от сточных промышленных вод в окрестностях Мукачево жерлянки оставались активными и в январе при морозах до -16°.
Размножаться начинают вскоре после пробуждения, на равнинах, с середины марта. Крик может быть передан как "хуу... хуу...".
Если суточный ритм "пения" желтобрюхих жерлянок в Центральной Европе удивительно точно совпадает с таковым у краснобрюхих, то об изменениях интенсивности пения в разные месяцы этого сказать нельзя. Прежде всего, у желтобрюхой жерлянки вместо двух максимумов в течение сезона наблюдается три, и все они несколько сдвинуты по сравнению с краснобрюхой жерлянкой на более позднее время. Первый пик приходится у нее на 27 апреля - 1 мая, второй - на 9-21 мая и третий - на 24-28 мая. Следовательно, размножение у этого вида еще больше растянуто, чем у краснобрюхой. Кладки икры встречаются до конца июля. Одна самка откладывает до 100 и более икринок. При температуре 15-18° головастики появляются через 10-12 дней после откладки икры и имеют длину 6-7 мм. Ко времени метаморфоза, который наступает через 2-2,5 месяца, они достигают длины 50 мм. Поздно вылупившиеся головастики зимуют.
В Северо-Восточном Китае, Корее и на Дальнем Востоке живет дальневосточная жерлянка (Bombina orientalis). На севере в нашей стране она доходит примерно до 50° с. ш. и на запад - до 130° в. д.
Брюшко у нее оранжевое или красное с черными пятнышками, сверху она серая, чаще с темными пятнышками. Концы пальцев сверху светлые. Голень примерно равна или длиннее ступни. Длина тела до 52 мм.
Держится в текучей воде под камнями, а в брачный период - в водоемах со стоячей или слабопроточной водой. Просыпается в мае, икру мечет с середины мая до конца июля. Икринки прикрепляются к камням. Одна самка откладывает около 100 икринок. Диаметр яйца в среднем 2 мм. На зимовку уходит в октябре.
В Китае в горах Юньнаня (1800-2600 м) живет самый крупный вид - большая жерлянка (Bombina maxima), длина тела ее достигает 68 мм.
В отличие от жерлянок представители другого рода этого же семейства - жабы-повитухи (Alytes) живут почти исключительно на суше.
Обыкновенная жаба-повитуха (Alytes obstetricans) встречается от Средней Европы до Пиренейского полуострова. Это небольшое животное длиною около 50 мм; с верхней стороны оно пепельно-серое с желтоватым, буроватым или зеленоватым оттенком; с нижней стороны - беловатое или желтовато-серое. Бородавки частью темные, черные или желтовато-белые, продольный ряд их, идущий от глаза к бедру, беловатый, иногда ярко-красный. В отличие от жерлянок имеет барабанную перепонку. Близкая к ней иберийская жаба-повитуха (Alytes cisternasii) населяет Испанию и Португалию.
Живет жаба-повитуха исключительно в холмистых и гористых местностях (до 2400 м). Предпочитает меловые почвы, часто встречается в старых каменоломнях. Но вообще места обитания весьма разнообразны, так же как и водоемы, в которых развиваются головастики. Активна ночью. Днем прячется в норах, ямах, зарывается в почву. Хорошо роет и передвигается по вертикальным плоскостям.
Корм жаба-повитуха, подобно лягушкам, собирает с растений, камней или ловит в воздухе, а не на земле, как это делают настоящие жабы. Питается различными беспозвоночными. Зимует на суше, в норах и пещерах.
Размножение во Франции продолжается с марта до августа; в Германии - с начала мая до конца июля. Голос самца повитухи звучит приятно, как звонкий стеклянный колокольчик. Икру выметывает 3-4 порциями, и одна самка откладывает всего 120-150 яиц. Яйца заключены в два четкообразных шнура, каждый из которых достигает длины 80-170 см. Яйца лежат на расстоянии 4-7 см друг от друга, и число их в одном шнуре от 18 до 54. Численность самцов в природе больше численности самок.
Самое удивительное в биологии жабы- повитухи - это особая форма заботы о потомстве. Спаривающийся самец двумя средними пальцами задней ноги схватывает конец яйцевого шнура, выходящего из клоаки самки, и, постепенно вытаскивая его, наматывает себе на бедра. Затем он носит икру на себе, пока не придет время вылупления головастиков. В отличие от большинства других бесхвостых земноводных откладка икры и оплодотворение у повитухи происходит, как правило, на суше. В искусственных условиях при высоких температурах (от 25 до 30°) спаривание происходит в воде, но в этом случае самец икру не наматывает на лапы.
В период откладывания яиц самцы яростно дерутся из-за самок. Там, где много повитух, один самец может спариться и носить икру от двух и даже от трех самок. Самец бродит со своей ношей повсюду, и она не мешает его нормальной жизнедеятельности. Яйца повитух сравнительно долго могут противостоять высыханию. Это объясняется особенностями строения их оболочек,, прозрачное слизистое вещество которых пронизано волокнами, образующими ряд слоев. Каждое волокно изгибается, некоторые ветвятся. Соседние слои волокон пересекаются под прямым углом, в результате оболочки яиц повитухи настолько тверды, что на ощупь кажутся кожистыми и надежно защищают развивающийся зародыш не только от высыхания, но и от механических повреждений. Однако они не способны разбухать в воде в такой степени, как лишенные волокон однородные оболочки яиц других земноводных.
Развитие икры в зависимости от погоды продолжается от 3 до 7 недель. К моменту вылупления головастиков самец направляется в водоем и начинает в нем торопливо плавать. Личинки в течение немногих минут покидают яйцевые оболочки, разорванные благодаря движениям их хвоста. Стряхнув с себя детенышей, самец снимает с ног пустые яйцевые шнуры и, не заботясь более о личинках, снова отправляется на сушу (рис. 49).
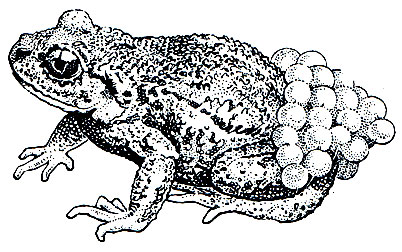
Рис. 49. Самец жабы-повитухи (Alytes obstetricans) с кладкой яиц на задних ногах
Вылупившиеся личинки могут жить в небольшом количестве воды. О характере их питания существуют разноречивые сообщения. Относительно небольшая длина их кишечника (только в 4 раза больше, чем у взрослых) свидетельствует о плотоядном типе питания. Однако есть указания на то, что головастики повитухи питаются растительной пищей. Некоторые полагают, что водоросли могут поддерживать жизнь головастиков и их рост, но их недостаточно для метаморфоза.
Превращение личинок заканчивается в конце июля - начале октября. Однако для повитухи очень характерно длительное развитие головастиков - в течение нескольких лет. Это наблюдается как в неволе, так и в естественных условиях. В Швейцарии вылупившиеся головастики имеют длину 16-17 мм и уже лишены наружных жабр, которые у зародыша, находящегося в яйце, достигают очень большой длины. Спустя восемь дней личинки достигают 32 мм длины, примерно через 4 месяца, т. е. в октябре,- 55, в марте следующего года - 65, в мае - 76 мм. В июне, т. е. через год, происходит метаморфоз. По наблюдениям, в неволе независимо от того, два года длилось личиночное развитие или три, к середине сентября рост головастиков прекращается и возобновляется лишь в апреле, несмотря на то что в течение этого времени потребление головастиками пищи не уменьшалось. Перезимовавшие головастики при метаморфозе превращаются в молодых повитух заметно более крупного размера, чем метаморфизировавшие тем же летом. В Пиренеях в озере на высоте 2400 м условия существования для головастиков повитухи очень неблагоприятны; в связи с этим задние конечности у них появляются только через 13-14 месяцев. Дальнейшее развитие длится несколько лет, а некоторые личинки имеют возраст до 20 лет.
В искусственных условиях ускорить метаморфоз можно усиленным освещением, высокой температурой, малым количеством воды и взбалтыванием ее, а также внезапно наступившим голодом.
Когда икра повитухи развивается в воде, то превращение наступает в том же году. Если личинок, вылупившихся из икры, созревшей на суше, не пускать в воду и заставить их прожить еще несколько недель на суше, то, попав в конце концов в водоем, они развиваются чрезвычайно ускоренно и быстро вновь выбираются на сушу. Увеличение продолжительности личиночной стадии может быть достигнуто преждевременным перенесением личинок в воду тогда, когда у них еще имеются наружные жабры, а также воздействием на них темнотой, низкой температурой, обилием спокойной воды, внезапным откармливанием после предварительного голодания. Длительное развитие личинок повитухи приводит к тому, что головастик перед метаморфозом составляет 174% длины взрослых.
Иногда самцы сбрасывают с себя комки икры при испуге или при протискивании в узкую щель между корнями и камнями. В потерянных комках икры личинки развиваются так же, как и в тех, которые самец таскает за собой. Личинки способны жить без воды на сырой земле до 4 недель. Кожа у них становится толще, рано развиваются кожные железы, обильно выделяющие слизь, быстро формируются легкие. При этом личинки скучиваются, благодаря чему лучше сохраняется влага.
Еще один вид этого семейства, относящийся к особому роду, - дискоязычная лягушка (Discoglossus pictus) - обитает на территории Юго-Западной Европы (Франция, Испания, Италия, острова Среди-земного моря) и Северо-Западной Африки (Тунис, Алжир, Марокко). Размером она до 75 мм и внешне напоминает травяную лягушку. Ее гладкая кожа серо-бурая или красноватая, обычно с темными пятнами, которые нередко образуют два-три продольных ряда. Нижняя поверхность тела светлая, часто с буроватыми крапинками. Барабанная перепонка скрыта под кожей. Язык круглый.
Держится она по берегам рек, ручьев и озер. Оптимальная температура для нее 25°. Негромкий голос самцов можно передать как быстро повторяющиеся "ха-а, ха-а" или "ва-а". Размножается в течение всего теплого периода, откладывая икру 5-6 раз. Всего самка за сезон откладывает до 6 тысяч мелких яиц. Кладки икры в виде лепешек лежат на дне водоема. Головастики развиваются около двух месяцев, и метаморфизировавшие сеголетки имеют в длину около 30 мм.
Последний вид семейства круглоязычных, также выделенный в особый род Barbourula, сравнительно недавно открыт и еще не изучен. Представители его обитают в водоемах Филиппинских островов и характеризуются тем, что пальцы передних лап соединены у них перепонкой.
Семейство Пиповые (Pipidae)
Подобно круглоязычным, у пиповых вогнутые сзади позвонки. Вероятно, в связи с давним переходом к полностью водному образу жизни у них исчез язык. К этому семейству относится всего 12 видов, обитающих в Африке (8 видов) и в Южной Америке (4 вида).
Широкой известностью пользуется род шпорцевых лягушек (Xenopus), распространенных в Южной Африке к югу от Сахары и имеющих ногти на трех внутренних пальцах задних лап.
Гладкая шпорцевая лягушка (Xenopus laevis) имеет маленькую приплюснутую голову, короткую круглую морду с обращенными кверху ноздрями и маленькими глазами, имеющими круглый зрачок. Верхнее веко у нее атрофировалось до небольшой кожной складки, что, видимо, так же как и редукция языка, связано с водным образом жизни. Под глазом располагается короткая осязательная нить, напоминающая осязательную нить безногих земноводных. Пальцы передних ног тонкие, сравнительно длинные, не соединены плавательной перепонкой. На задних же ногах развита широкая плавательная перепонка и три первых пальца вооружены острыми черными когтями. На теле имеются органы боковой линии. Сверху лягушка темно-бурого или оливково-зеленого цвета, часто с большими темными неправильной формы пятнами, снизу - беловатая, одноцветная или с бурыми пятнами. Самка отличается тем, что задний проход закрывается у нее тремя кожными клапанами. Длина взрослых особей до 80 мм.
Встречается в Южной Африке, на севере - до Анголы и Килиманджаро. Живет исключительно в воде и кормится только под водой, помогая себе засовывать пищу в рот передними лапами.
Спаривание и икрометание в неволе происходит в течение всего лета. В это время у самца развиваются так называемые "брачные щетки" в виде черной полосы по бокам пальцев и вдоль конечности до подмышечных впадин. Самец обхватывает самку перед задними конечностями. Спаривание происходит ночью. В это время самцы громко квакают, и голос их напоминает голос квакши. Откладка яиц начинается приблизительно через час после начала спаривания. Яйца откладываются большей частью по одному, реже по 3-4, быстро одно за другим. Одна самка может отложить до 15 000 икринок. Процесс икрометания своеобразен и обнаруживает интересные приспособления, увеличивающие возможность оплодотворения. Откладываемое яйцо сначала держится между тремя кожными лоскутами, окружающими клоаку самки. Спаривающаяся самка держится в это время задними ногами за какое-нибудь водное растение. Затем клоака ее приближается к переднему концу продольной борозды, которая тянется по брюху самца до его клоаки и ограничена двумя кожными складками. Яйцо быстро скользит по этой борозде кзади, проскальзывает под клоачным отверстием самца, оплодотворяется и повисает на растении, которое придерживает самка. Следующая икринка откладывается уже в другом месте. Яйца имеют в поперечнике около 3 мм и окружены прозрачной, чрезвычайно липкой студенистой оболочкой, наружный слой которой в течение немногих часов откладки икры твердеет и становится похожим на рог.
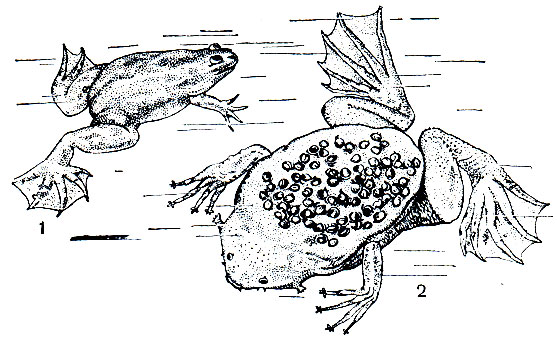
Рис. 50. Шпорцевая лягушка (Xenopus mülleri) - 1 и пипа суринамская (Pipa pipa) - 2 - с кладкой яиц на спине
Обычно на утро после спаривания оба животных линяют, причем, как у многих других бесхвостых земноводных, кожа отстает сначала на ногах, затем продвигается ко рту и наконец проглатывается.
При температуре 22° личинки вылупляются через двое суток. По строению рта личинка шпорцевых лягушек совершенно не похожа на головастиков других бесхвостых земноводных, а скорее напоминает личинок хвостатых. У нее нет воронки, окружающей ротовое отверстие, роговых челюстей и зубов. Питается она фильтруя пищу из воды. Цедильный аппарат помещается у головастика на жаберных дугах вместо внутренних жабр, которые не развиваются. Головастик заглатывает воду ртом и выпускает ее через жаберные отверстия, которых у него, в отличие от других бесхвостых, два, и располагаются они с обеих сторон тела. Кормящийся головастик держится в воде почти вертикально, совершая последней третью хвоста непрестанные волнообразные движения. В течение короткого времени личинки могут сделать воду, зеленую от находящихся в ней жгутиковых, совершенно прозрачной.
К моменту вылупления личинки уже утрачивают наружные жабры. Через два часа, после того как начинается самостоятельное питание, включается легочное дыхание. В первые дни жизни на нижней стороне головы у личинки имеется цементная железа, при помощи которой животное держится на месте. Вскоре этот орган исчезает. На третий-четвертый день после вылупления морда головастика становится клиновидной, а в углах рта появляются длинные осязательные нити, аналогичные органам равновесия у личинок тритонов. Предполагают, что эти усики ощущают приближение добычи, невидимой в мутной воде. Обычно головастик держит усики, вытянутыми вперед; лишь при быстром подъеме на поверхность воды они направляются кзади. От оплодотворения яйца до полного превращения при температуре 22° проходит 2,5-3 месяца. Собственно метаморфоз занимает из этого времени 15-20 дней. Превращение начинается с того, что появляются передние конечности. Они выходят с обеих сторон из покрывающего их мешочка над жаберным отверстием, а не из него, как это происходит у других бесхвостых. В результате образ жизни личинки не изменяется, тогда как у головастиков других бесхвостых прорыв передних конечностей прекращает дыхание внутренними жабрами.
Три дня спустя после появления передних конечностей задние лапы начинают проделывать плавательные движения, принимая характерное для взрослого животного положение. Теперь они быстро растут, и на трех внутренних пальцах появляются черные когти. Тело головастика приобретает окраску, похожую на окраску взрослой лягушки. Через 14 дней после появления передних конечностей щупальца начинают спадаться, в течение 4-6 часов рот приобретает свою окончательную форму, жаберные отверстия закрываются. Животное все чаще и чаще остается на дне, хвост постепенно уменьшается в размерах, и личинка превращается в лягушку, но не покидает водоема и переходит на питание животной пищей - мелкими ракообразными и другими водными животными. Теперь передние лапы, как и у взрослых, помогают им запихивать пищу в рот. Характер метаморфоза, протекающего медленно, также приближает шпорцевых лягушек к хвостатым земноводным. Половая зрелость у самца наступает на второй год.
Еще 4 более мелких вида этого рода известны из различных мест Восточной и Южной Африки.
В Камеруне, Габоне и Конго встречается два мелких вида (28-46 мм) другого рода - Hymenochirus. У них плавательная перепонка имеется и на передних ногах. Один из видов - Hymenochirus boettgeri - в неволе размножается круглый год. Размножение стимулируется светом и временным повышением температуры. Икра откладывается в поверхностном слое воды порциями примерно по 12 штук. Всего самки откладывают по 150-200 икринок. Они быстро расплываются по поверхности воды и прикрепляются к растениям. Диаметр икринки - 0,9 мм. Вылупление происходит через 24 часа. Головастики начинают свободно плавать через 4-5 дней, а еще через 27 дней превращаются во взрослых животных. Питание головастиков происходит в течение всего метаморфоза.
Представители другого рода - Пипа (Pipa), обитающие в Америке, вовсе лишены зубов и замечательны своей особенностью размножения.
Самая крупная из них - суринамская пипа (Pipa pipa) достигает 200 мм длины. У нее почти четырехугольное сплющенное тело, широкая, сливающаяся с туловищем голова, заостренная спереди, тонкие передние ноги с длинными четырехраздельными на концах пальцами, не соединенными плавательной перепонкой. Задние ноги значительно толще, довольно длинны и имеют плавательную перепонку. Кожа, особенно у старых животных, морщинистая. От углов рта и впереди глаза свисают кожные лоскуты. Глаза малы и обращены вверх. Окраска желтоватая, переходящая иногда в черновато-бурую, нижняя сторона тела светлее, иногда с белыми пятнами, иногда же украшена черной полосой, идущей вдоль живота.
Живет суринамская пипа в Бразилии, Гайане, Гвиане Французской и Суринаме. Ведет водный образ жизни. Несмотря на это, у нее сильно развитые легкие и ороговевшая кожа - признаки, обычно хорошо выраженные у наземных форм. Пипа часто встречается в оросительных каналах на плантациях, во время засухи держится в почти высохших лужах, но, когда в период дождей ливни затопляют все лесистые местности, пипы, не покидая воды, бродят по всему лесу.
На период дождей приходится также и откладка икры. Голос самца пипы напоминает тиканье часов. Спаривающийся самец держит самку над задними ногами. Спаривание продолжается сутки. Не исключено, что у этого вида оплодотворение внутреннее. Клоака самки в виде большого мешка представляет собой своеобразный яйцеклад, который спаривающаяся самка продвигает под самцом к себе на спину. Самец нажимает на яйцеклад и поодиночке выдавливает из него яйца. Таким способом они почти равномерно распределяются по спине самки и попадают в расположенные на ней ячейки. Каждая ячейка имеет глубину 10-15 мм и вскоре после попадания в нее яйца принимает шестигранную форму. Сверху ячейки покрыты кожистой крышечкой диаметром 5-6 мм, образующейся из яйцевых оболочек. Перегородки, разделяющие ячейки, очень тонки и богаты кровеносными сосудами.
Яйца пипы богаты желтком и достигают больших размеров - 6-7 мм в диаметре. В начальной стадии развития они весят 2,95 г, а к концу - 3,37 г. Вероятно, они получают воду, а может быть, и питательные вещества из организма матери. Количество откладываемых яиц колеблется от 40 до 114. Развитие продолжается 82 дня. Вполне сформировавшееся молодое животное разрывает ячейку и покидает ее. Самка стирает затем остатки ячеек о камни или растения и после линьки покрывается новой кожей.
Семейство Чесночницы (Pelobatidae)
Это довольно многочисленное семейство, объединяющее 12 родов и более 50 видов. Более 75% из них обитает в Азии, особенно на юго-востоке материка. Там же распространены и наиболее примитивные виды, в том числе и представители самого обширного рода Megophrys. Представители других родов этого семейства распространены в Европе (Pelobates, Pelodytes), Северной Америке (Scaphiopus) и на некоторых островах (Sooglossus, Nesobia, Nesomantis). Среди чесночниц есть виды, имеющие особые приспособления к жизни на высокогорье, например 4 вида рода Scutiger и 5 видов рода Aelurophryne, живущие в Гималаях, Сычуани и Тибете на высоте до 4000 м над уровнем моря. К этому же семейству относятся причудливые чесночницы Сычуаня рода Vibrissaphora, у самцов которых многочисленные кожные шипики образуют подобие усов на верхней губе. Многие чесночницы хорошо роют и ведут полуподземный образ жизни.
Род Megophrys, объединяющий около 25 видов, характерен свободными межпозвонковыми дисками. Все виды живут в горах Южной Азии и Малайского архипелага. Среди видов этого рода есть как очень мелкие, например М. kempii из Юго-Восточной Азии, имеющие длину тела до 15 мм, так и крупные, например М. carinensis, достигающие длины 150 мм. Последний вид питается как насекомыми, так и мелкими млекопитающими. Личинки представителей этого рода отличаются огромным расширением ротового поля, имеющим вид воронки (рис. 17,2). При помощи такого аппарата они неподвижно, почти вертикально висят на поверхности воды, подобно тому как это делают при помощи своих щупалец личинки шпорцевых лягушек.
Обыкновенная чесночница (Pelobates fuscus) известна лучше других. Спина ее окрашена в желто-бурый или светло-серый цвет с крупными и мелкими бурыми и черными пятнами с красными точками. Кожа гладкая, временами издает сильный запах, напоминающий запах чеснока. Лоб между глазами выпуклый. Очень характерен для чесночниц большой лопатообразный внутренний пяточный бугор, развивающийся в связи с их роющей деятельностью. Самцы имеют на плечах овальную железу. Длина тела до 80 мм.
Распространена от Средней Европы до Аральского моря и юга Западной Сибири; на юге до Крыма, Северного Кавказа, а на севере - до линии Ленинград - Казань.
Рис. 51. Азиатская рогатая чесночница (Megophrys nasuta)
Чесночницы приходят в водоемы только в период размножения, проводя все остальное время на суше. Они прекрасно роют и день проводят зарывшись в землю, покидая убежище вечером.
В связи с роющим образом жизни чесночницы распространены неравномерно. Они избегают каменистых почв, вследствие чего область их распространения ограничивается почти исключительно равнинами с более или менее рыхлой глинистой или песчаной почвой. В мягком грунте чесночница зарывается очень быстро и через 2-3 минуты совершенно исчезает из виду. Роет она задними ногами, погружаясь в землю задом почти в вертикальном положении.
Так как чесночницы много времени проводят под землей, у них меньше развиты легкие.
Охотятся они ночью, и степень их активности находится в прямой зависимости от влажности. Чем больше влажность, тем активнее чесночницы. При температуре 10-15° и 78-89% влажности активны лишь 11% животных, а при этой же температуре, но при 90-95% влажности - 52%.
Число видов беспозвоночных, поедаемых чесночницей, велико. Однако существенную роль в ее питании играют муравьи, жужелицы и пауки. Эти преобладающие корма составляют 87%. Среди других наших земноводных чесночница отличается наиболее высоким процентом наземных кормов (99%), что свидетельствует о том, что этот вид почти не питается в водоемах. В исследованных желудках обыкновенной чесночницы вовсе не обнаружено летающих форм.
Среди наших земноводных чесночница раньше всех других уходит на зимовку, и период зимней спячки у нее длится около 200 дней. Зимует она на суше, зарываясь в землю или используя норы грызунов, глубокие ходы кротов, гнезда береговых ласточек. Просыпается чесночница под Киевом в марте, а севернее - в апреле - начале мая.
Мечет икру ранней весной, вскоре после пробуждения, задерживаясь в водоемах очень недолго. Вся популяция выметывает икру на протяжении 20-25 дней. Самцы чесночниц "брачных мозолей" не имеют; так же как жерлянки, они держат самок в поясничйой области. В брачный период самцы под водой издают булькающий звук вроде "тук, тук, тук". Кладка чесночницы представляет собой толстые колбасовидные шнуры, образованные слившимися наружными оболочками и содержащие внутри некоторое количество беспорядочно расположенных яиц. Икрометание происходит под водой, и шнуры обволакивают подводные предметы: упавшие сучки, водоросли. Одна самка откладывает 1200-2300 яиц диаметром 1,5-2,5 мм.
Развитие головастиков у чесночницы продолжается дольше, чем у других наших земноводных,- 90-110 дней, а в некоторых случаях и дольше. Наружные жабры у них исчезают на 7-й день, почки конечностей становятся заметными на 34-й день. На 67-й день задние конечности разделяются на суставы. На 92-й день появляются передние конечности, а на 94-й начинает редуцироваться хвост. Однако растут головастики у чесночницы быстрее, чем у всех изученных в этом отношении наших земноводных. Средний прирост за день составляет у них 1,2 мм. Перед метаморфозом длина этих гигантских головастиков на 40% превышает длину взрослой самки и равна 73-175 мм.
Первое время головастики чесночницы ведут придонный образ жизни, в это время они малозаметны. Во второй половине личиночной жизни они плавают в толще воды. Питаются головастики растительными кормами. В связи с этим длина кишечника у них очень велика и почти в 6 раз превышает длину тела. У головастиков, питающихся растительной пищей, ротовая воронка глубокая, с крупной бахромой по краям, мощным роговым клювом и хорошо развитыми многочисленными зубчиками. Все эти признаки по сравнению с другими земноводными особенно ярко выражены у чесночницы. Число зубчиков на ротовой воронке у нее равно 1100, т. е. вдвое больше, чем у головастиков травяной лягушки или квакши.
Скорость развития головастиков зависит от внешних условий. В глубоких водоемах под Москвой температура может оказаться столь неблагоприятной для головастиков чесночницы, обычно обитающих южнее, что они не успевают метаморфизировать и зимуют на личиночной стадии. Влияет на скорость развития и характер пищи. Длительное развитие головастиков чесночницы обычно связывают с тем, что они питаются растительной пищей. Метаморфизировавшие сеголетки здесь же на берегу водоема закапываются в почву, где и остаются до весны следующего года, когда начинается их расселение.
Другой вид - сирийская чесночница (Pelobates syriacus) - живет в Сирии, Малой Азии и Закавказье. Сверху она светло-серого, иногда желтоватого цвета с темно-зеленой неправильной пятнистостью; напоминает по окраске зеленую жабу. Брюхо беловатое, без пятен. Кожа гладкая. Лоб между глазами плоский. Перепонки между пальцами задних ног с глубокими вырезами. Самцы мельче самок.
В отличие от обыкновенной чесночницы сирийская чесночница может жить на более плотных почвах. В Армении обитает на глинистых почвах с каменистыми включениями. В сутки 18-19 часов она проводит зарывшись в почву, обычно на глубине 10-15 см, иногда 25 см от поверхности земли. Постоянных убежищ не имеет. Зарывается там, где ее застанет рассвет. Весной и летом предпочитает зарываться в более влажную почву (30-35% влажности). Потеря воды, равная 40% от веса тела, приводит к гибели животного. Осенью чесночницы зарываются в более сухую почву, влажность которой составляет всего 3-5%. Объясняется это, вероятно, тем, что теплопроводность в сухих почвах значительно ниже и в них сравнительно теплее, чем во влажных.
В кормовом рационе сирийской чесночницы около 60% занимают насекомые, 30% - моллюски, 9% - дождевые черви. Среди насекомых преобладают жуки и прямокрылые.
В окрестностях Еревана в конце октября - начале ноября, когда средняя температура воздуха равна 7-8°, сирийская чесночница уходит на зимовку. Зимуют поодиночке. Весной появляются в первой половине марта примерно при той же температуре.
Икрометание происходит в постоянных, непересыхающих водоемах. В зависимости от высоты над уровнем моря оно бывает с конца марта до середины мая. Период икрометания длится 10-15 дней.
При температуре воды 17° головастики выводятся через 7 дней и имеют длину 3-4 мм. Покинув яйцевые оболочки, они опускаются на дно водоема. Когда головастики достигнут 7-8 мм, они прикрепляются развившейся присоской к растениям. В это время у них появляются зачатки наружных жабр в виде четырехлопастных пластинок. На 13-14-й день головастики достигают 12-13 мм, у них прорывается рот и они начинают свободно плавать. На 50-55-й день их длина достигает 143 мм и задние конечности у них формируются полностью. На 65-70-й день прорываются передние конечности, а спустя 5-6 дней начинает рассасываться хвост. Все развитие длится 70-85 дней. Иногда головастики зимуют. Длина тела сеголеток 33-35 мм, а перед уходом на зимовку 43-45 мм.
Третий вид этого рода - Pelobates cultripes - живет в Южной Франции, Испании, Португалии и Северо-Западной Африке.
В Северной Америке, от Юго-Западной Канады до Южной Мексики, распространены лопатоноги (Scaphiopus). У них такое же, как у чесночниц, плотное телосложение, мощный лопатообразный пяточный бугор и вертикальный зрачок. Различаются 6 видов этого рода размерами (от 38 до 72 мм) и окраской. По образу жизни напоминают наших чесночниц. День они проводят зарывшись в землю. Вечером выходят на добычу, и в это время суток весной и летом после дождя можно слышать их голоса. Способ спаривания и форма яйцевых кладок такие же, как и у наших чесночниц, но развитие проходит быстрее, и метаморфизировавшие сеголетки образуют массовые скопления по берегам водоемов.
Если по внешнему виду и образу жизни лопатоноги больше похожи на чесночниц, то по анатомическому строению они ближе к крестовкам, представляющим еще один род этого же семейства.
В Западном Закавказье живет кавказская крестовка (Pelodytes caucasicus). В отличие от чесночниц внутренний пяточный бугор у крестовок округлый, небольшой. Это горная форма, поднимающаяся до 2300 м и выше над уровнем моря. Живет около водоемов и в тенистых местах. Осторожна и малозаметна. Деятельна ночью.
Самцы кавказской крестовки в брачный период темнее самок. На спине у них редко разбросанные красные пятна, которые иногда находятся в кратерообразных углублениях. На спине, боках, вдоль нижнего края нижней челюсти и на мозолях расположены многочисленные роговые образования в виде заостренных и несколько изогнутых шипиков. Брюшко в нижней его части усеяно черными пятнами, которые тоже представляют собой роговые образования. Борозды и бугорки на спине покрыты сплошным роговым слоем, поверхность которого неровная и образует местами сильно выступающие утолщения. Все эти роговые образования в конце брачного периода исчезают при линьке, и их можно видеть на сброшенной кожице. После линьки меняется и окраска самца. На спине постепенно вырисовывается контур светлоокрашенного косо лежащего креста, у нижних концов которого появляются два светлых пятна; верхняя часть брюшка принимает мраморную окраску; полосы на задних ногах становятся ярче. Самцы крупнее самок. Свое присутствие в водоеме они обнаруживают, издавая специфические горловые звуки.
Самка откладывает икру не только в маленьких водоемах со стоячей водой, но и в речках, выбирая места с меньшим течением. В типичных случаях кладка имеет форму цилиндра и содержит от 20 до 400 икринок. Вдоль кладки, посередине ее, проходит какой-нибудь предмет - прутик или травинка. Период икрометания в районе Боржоми длится со второй половины июня до середины сентября. Головастики, вышедшие из поздних кладок, зимуют. Питаются они водорослями и гниющими частями растений, а в период формирования задних конечностей кормятся трупами животных. Активны как ночью, так и днем.
Другой вид - пятнистая крестовка (Pelodytes punctatus) - обитает во Франции, Северо-Восточной Италии, некоторых местностях Португалии и Испании. Отличается очень растянутым периодом размножения. Возможны две кладки в году. Личинки встречаются круглый год.
Упомянем еще одного представителя этого семейства - живущую на Сейшельских островах сейшельскую чесночницу (Sooglossus seychellensis), которая отличается своеобразным способом размножения. В местах ее обитания текут лишь бурные потоки; видимо, в связи с этим ее развитие происходит на суше. Крупные яйца откладываются на землю, на влажные листья, а вылупившиеся головастики забираются на спину самца, сторожащего яйца, и развиваются там до конца метаморфоза. Дышат они через кожу, главным образом хвоста, а затем включаются легкие.
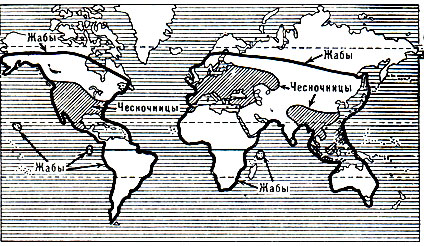
Рис. 52. Области распространения семейств жаб и чесночниц
Семейство Жабы (Bufonidae)
Семейство жаб очень большое, включает около 450 видов, распространенных во всех частях света. Подразделяется на 7 подсемейств. Большинство жаб полностью лишено челюстных зубов.
К подсемействуавстралийских жаб относится 49 наиболее примитивных видов, объединяемых в 16 родов.
Один из наиболее известных видов - пустынная австралийская жаба (Chiroleptes platicephalus) отличается способностью противопоставлять внутренний палец передних ног всем остальным, большой головой, сильно развитыми плавательными перепонками, соединяющими пальцы задних ног, и неявственной барабанной перепонкой. Верхняя челюсть у нее с зубами. Легкие у этой жабы очень сильно развиты и доходят до заднего конца тела. В пустынях Центральной Австралии она живет в котлованах с твердой глинистой почвой. Засушливый период проводит зарывшись в норе на глубину около 30 см. Тело жабы шарообразно раздувается от переполнения водой и занимает всю нору, стенки которой бывают влажными. Воду животное запасает в больших подкожных полостях и в полости тела. Местные жители отыскивают этих жаб в пустыне и используют их как запасы питьевой воды.
Широко распространена в Австралии лягушковидная жаба (Limnodynastes tasmaniensis), по внешнему виду действительно похожая на лягушку; отличается красивой окраской. Верхняя поверхность ее бурая, желтоватая или красноватая с неправильно расположенными большими темно-бурыми и зелеными пятнами, часто со светлыми краями. Светлая линия на спине желто-зеленая или оранжевая. У самцов зеленое горло. Откладывают икру в воду в виде белых пенистых комков. Икрометание, как у большинства австралийских бесхвостых, не приурочено к определенным срокам, а связано с дождями, обычно с июля до мая.
Некоторые австралийские жабы размножаются на суше. Так, жаба Crinia laevis откладывает яйца под землей в гладкостенных норах, которые животное выкапывает само, или использует готовые пустоты. Ложная жаба Биброна (Pseudophryne bibroni), как и другие представители этого рода, внешне похожа на жерлянку и по складу тела, и по яркой окраске брюха. Она откладывает яйца поодиночке на суше между камнями или среди пучков травы на берегу водоема. Период икрометания приходится на австралийское лето и осень. Из отложенных яиц, часто по истечении нескольких месяцев, при первом дожде выходят развившиеся головастики. Другой вид того же рода - P. corroboree имеет на спине характерные продольные черные полосы, чередующиеся с блестящими желтыми полосами, живет на границе снегов в альпийском поясе и многие месяцы проводит в зимней спячке. Размножается сходно с P. australis, которая откладывает около 20 яиц в ямки поблизости от воды. Потоки дождя уносят яйца в воду, где и развиваются выведшиеся из них головастики.
Единственный вид еще одного рода австралийских жаб - Myobatrachus живет в термитниках.
Подсемействонастоящих жаб близкородственно с австралийскими жабами. Основная масса видов (около 250) принадлежит к роду Bufo, представители которого распространены по всем континентам, кроме Австралии.
Зеленая жаба (Bufo viridis) окрашена сверху в светло-серо-оливковые тона с крупными темно-зелеными пятнами, отороченными узкой черной каймой. Кожа бугорчатая, по бокам головы два крупных скопления ядовитых желез - паротиды. Распространена в Европе, Северной Африке, в Европейской части СССР, в Крыму, на Кавказе, в Казахстане, в Средней Азии, Иране, Турции, Афганистане, на восток идет до Алтая, Западной Монголии, Западного Китая. У Балтийского моря доходит на севере до 59° с. ш., а на востоке ареала до 50° с. ш. Широко распространена в полупустынях, степях и широколиственных лесах, идет до северной границы смешанных лесов. Дальше других земноводных проникает в пустыни, высоко поднимается и в горы. В Гималаях найдена на высоте до 4500 м.
Максимальные размеры зеленой жабы - 140 мм. На Кавказе, в зоне оптимума для этого вида, средняя длина - 75 мм. К северу, югу, востоку, а возможно, и к западу размеры тела зеленых жаб уменьшаются.
Несмотря на широкое распространение, зеленая жаба обладает качествами, характеризующими ее как южное животное. Так, ее мышечная ткань отличается значительной теплоустойчивостью, и лучше всего этот вид в опытных условиях чувствует себя при 33°.
Зеленая жаба ведет наземный образ жизни, проводя в водоемах лишь незначительное время в период икрометания. Занимает наиболее сухие места обитания, недоступные другим земноводным. Специальными наблюдениями установлено, что 91 % зеленых жаб встречался в местах, где влажность припочвенного слоя воздуха равна 51-90%. При более высокой влажности - от 91 до 100% - встречено лишь 9% этих животных. Приспособленность к существованию в сухих местах обитания развивается у земноводных в двух направлениях: они обладают меньшей проницаемостью кожи для воды и способны терять большое количество влаги без вреда для организма. Проницаемость кожи для воды у зеленой жабы в 2 раза меньше, чем у остромордой лягушки, и в 3 раза меньше, чем у травяной. Зеленые жабы переносят потерю воды, равную 50% веса их тела, тогда как травяные лягушки погибают, потеряв воду в количестве 15% от веса тела. Проникновение воды через кожу регулируется консистенцией слизи, покрывающей ее. При недостатке влаги в окружающей среде кожа земноводных покрывается тонкой сухой, блестящей пленочкой, малопроницаемой для воды.
Зеленая жаба ведет ночной или сумеречный образ жизни, выбирая для этого самое влажное время суток. Вышедшие на охоту животные часто "купаются" в водоемах или в росе, пополняя запасы воды в теле.
Летом зеленые жабы начинают кормиться около 8 часов вечера. К 11-12 часам ночи их желудки бывают максимально наполнены. Осенью жабы выходят на охоту уже в 5 часов вечера. Иногда они бывают активны и днем.
Все корма зеленая жаба добывает на суше. Лишь 0,1% от общего числа пищевых объектов приходится на долю водных организмов. В Предкавказье ведущую роль в питании этого вида играют жуки, клопы, гусеницы и муравьи. В Армении почти половина пищевых объектов представлена жуками, большое значение имеют личинки мух, уховертки, гусеницы, клопы. В Дагестане 40% из числа всех съеденных животных составляют жуки, 22% - прямокрылые, 11% - клопы и 11% - муравьи. Летающих насекомых зеленая жаба добывает очень редко. Это связано с тем, что охотящиеся жабы передвигаются небольшими прыжками; создается впечатление, что животное идет "шажками". Из-за относительно коротких задних ног с менее развитой мускулатурой жаба не в состоянии делать сильных прыжков, устремляясь за добычей. У жабы малоподвижный язык, на значительном протяжении прикрепленный ко дну ротовой полости, способен лишь немного вываливаться набок. Его движения не могут компенсировать малую подвижность животного. Угол, образуемый телом и горизонтальной поверхностью, у жаб невелик. Это также способствует тому, что они берут добычу в "нижнем ярусе", с земли.

Таблица 9. Лягушка черно-пятнистая (Rana nigromaculata)

Таблица 9а. Лягушка прудовая (Rana esculenta)
Единственное средство защиты этого небыстро передвигающегося животного - ядовитые железы. Кожа спинной стороны тела жаб несет две крупные околоушные ядовитые железы - паротиды и множество мелких одиночных ядовитых желез. Ядовитые железы жаб лишены аппаратов, позволяющих наносить ранения и вводить свои выделения непосредственно в кровь, следовательно, ядовитые железы жаб пассивные. Мелкие одиночные ядовитые железы имеют открытый выводной проток. Когда хищник, преследующий жабу, хватает ее, из всех малых ядовитых желез рефлекторно выделяются вещества с резким специфическим запахом, чрезвычайно горьким вкусом, жгучим и рвотным действием. Нападающее животное оказывается вынужденным бросить жертву. До тех пор пока жаба не схвачена, как бы ее ни преследовали, выделения из желез не происходит. Протоки околоушных желез закрыты особыми пробками, препятствующими выделению секрета наружу. Это оказывается возможным только при надавливании на железу. Выделения околоушных желез, попав в организм хищника, отравляют его. Их защитное значение оказывается особенно велико в период спячки, когда все рефлекторные реакции резко понижаются и деятельность малых ядовитых желез сводится до минимума. Околоушные железы, выделяя яд при надавливании на них, действуют независимо от физиологического состояния организма. Для человека ядовитые выделения наших жаб не опасны.
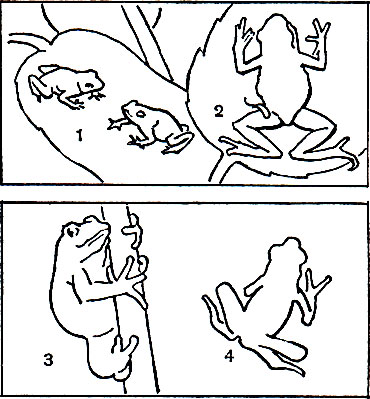
Таблица 10. Бесхвостые земноводные: 1 - древолаз маленький (Dendrobates pumilio); 2 - листолаз двуцветный (Phyllobates bicolor); 3 - африканская веслоногая лягушка (Hylambates maculatus); 4 - венесуэльский короткоголов (Atelopus cruciger)

Таблица 10. Бесхвостые земноводные: 1 - древолаз маленький (Dendrobates pumilio); 2 - листолаз двуцветный (Phyllobates bicolor)

Таблица 10. Бесхвостые земноводные: 3 - африканская веслоногая лягушка (Hylambates maculatus); 4 - венесуэльский короткоголов (Atelopus cruciger)
В пустынях у зеленой жабы, видимо, хорошо выражена летняя спячка, нередко переходящая в зимнюю. На зимовку она уходит раньше многих земноводных, вслед за чесночницами. Севернее и в горах активность ее прекращается раньше. Зеленые жабы начинают уходить на зимовку, когда среднесуточная температура воздуха снижается до 7-8°, и исчезают полностью, когда она снизится до 3-4°. Зимуют там же, где скрываются днем: в норах грызунов, ямах, под камнями, в щелях стен, зарываются в рыхлую землю на глубину 10-12 см. Спячку проводят поодиночке, иногда по 3-4 особи вместе.
В Армении одиночные выходы зеленых жаб наблюдаются при среднесуточной температуре 3-9°; массовое появление происходит только с повышением ее до 15-16°. В окрестностях Еревана это происходит в конце марта - в начале апреля, выше в горах и на северных границах распространения вида - в конце апреля - в мае. Средняя продолжительность зимовки - 185 дней; в окрестностях Еревана, на высоте 980 м, - 120 -130 дней; в окрестностях Севана, на высоте 1940 м,- 175 - 180 дней.
Проснувшись, зеленые жабы приступают в размножению. В это время половозрелые особи скапливаются в водоемах.
В Армении жабы достигают половозрелости на 4-й год. В это время длина их 50-55 мм.
У самца, обнимающего самку при спаривании, кисти лап помещаются у нее под мышками. Брачные мозоли расположены на трех средних пальцах передних лап и по размерам уступают этим образованиям у лягушек.
Отметав икру, жабы покидают водоем. Откладка икры разными особями происходит неодновременно, и благодаря этому икрометание растягивается до июля, а иногда и до августа. Кладка в виде шнура, в котором яйца расположены двумя рядами. Общая длина шнура может доходить до 7 м, а количество яиц в одной кладке достигает 12 800 штук. Яйцеклетки сравнительно мелки, диаметр их 1-1,5 мм. Икра откладывается около берега в самых разнообразных водоемах. Шнуры оплетают подводные предметы или лежат на дне. При средней температуре воды 23° головастики вылупляются на 5-6-й день, достигая 6-7 мм. Сначала они прикрепляются к студенистой массе шнуров, а затем к подводной растительности и другим предметам. Через 3-4 дня у них прорывается рог и они начинают интенсивно питаться.
Переходя к активному образу жизни, головастики держатся на дне и в связи с этим отличаются приплюснутой в спинно- брюшном направлении формой тела. Передний отдел головы у них несколько вытянут в хобот, а ноздри pi глаза сближены и отодвинуты назад. Эти особенности строения, видимо, связаны с тем, что они роются на дне в иле. Органы боковой линии у них развиты лучше, чем у головастиков лягушек, особенно зеленых. Их небольшой хвостовой плавник почти одинаков по ширине на всем протяжении и лишь слегка суживается к концу. Несмотря на то что головастики жаб, ведущие придонный образ жизни, обитают в условиях недостатка кислорода, наружные жабры у них коротки и не ветвятся.
Видимо, это связано с их роющим образом жизни. Однако сеть кровеносных сосудов в хвостовом плавнике у них значительно гуще, чем у головастиков- других видов. Это особенно важно, так как легочное дыхание у них включается поздно. Для них также характерна меньшая, чем у других видов, потребность в кислороде.
Головастики зеленой жабы могут жить в температурных пределах от 1° до 45. О питании их имеются отрывочные сведения, указывающие на то, что они поедают диатомовые и зеленые водоросли, гниющие остатки растений и животных, но предпочитают животную пищу. Ведут дневной образ жизни.
Головастики зеленой жабы отличаются самым быстрым развитием среди всех наших бесхвостых земноводных. На 5-й день у них исчезают наружные жабры, на 20-й появляются почки задних конечностей, на 30-й задние конечности разделяются на суставы, на 38-й появляются передние конечности, на 39-й начинается резорбция хвоста. Растут головастики не очень быстро. Средний прирост за день равен 0,8 мм. Длина перед метаморфозом составляет только 53% от длины взрослой самки.
В естественных условиях развитие длится 45-55 дней при средней температуре воды 21,5°, а при температуре 24,8°-55-60 дней. Размеры метаморфизировавших сеголеток достигают 14-16 мм.
В зоне оптимума половозрелость наступает в среднем при длине в 57 мм. По краям ареала - раньше (42-57 мм), так как рост здесь идет, как правило, более энергично. После наступления половозрелости рост в оптимальных условиях продолжается еще долго, и животное достигает больших размеров. Чем дальше от оптимума, тем резче затормаживается рост, и в итоге образуются популяции из более мелких особей.
В степном и лесостепном Предбайкалье и Забайкалье и на юге Дальнего Востока обитает монгольская жаба (Bufo raddei). Она отличается от зеленой жабы светлой полосой на спине, менее бугристой кожей и присутствием на голени большой железы. Монгольская жаба не распространяется так далеко на север, как зеленая. За пределами нашей страны она обитает в степях и пустынях Центральной Азии. Высоко поднимается в горы. В Гобийском Алтае, например, она найдена на высоте 3800 м.
Всюду в Западной Европе, включая Южную Англию и Южную Швецию, обитает камышовая жаба (Bufo calamita), отсутствующая в Италии и на Балканском полуострове. В нашей стране этот вид встречается только на западе, доходя на восток до Минска и Бобруйска. По внешнему виду камышовая жаба очень похожа на монгольскую. Наиболее часто она встречается на песчаных дюнах и менее других наших жаб чувствительна к солености воды и почвы.
Широко распространена в Советском Союзе серая, или обыкновенная, жаба (В. bufo) - самая крупная из наших жаб (до 200 мм). Сверху она бурого цвета, снизу - грязно-белого или желтоватого.
Распространена обыкновенная жаба в Северо-Западной Африке, в Европе, в пределах СССР - в Крыму, на Кавказе, в Сибири и на Дальнем Востоке, в Корее, Японии и Китае. На север доходит до 65° с. ш., в восточной части ареала северная граница распространения спускается много южнее, достигая берега Охотского моря. Населяет все варианты леса и степную полосу, по южной границе которой примерно проходит и южная граница распространения этого вида. В горы поднимается до 3000 м над уровнем моря.
Обыкновенная жаба в средней полосе не так многочисленна, как живущие с ней бок о бок бурые лягушки. Она гораздо менее заметна, чем зеленая жаба, выдающая свое присутствие звонкой брачной трелью самца. По образу жизни обыкновенная жаба сходна с зеленой, но представляет собой вид, распространенный дальше на север. В связи с этим при повышении температуры ее мышцы раньше теряют возбудимость, чем мускулатура зеленой жабы. Она позднее зеленой жабы залегает в спячку и меньший срок бывает неактивна.
Обыкновенная жаба менее плодовита, чем зеленая. Она откладывает 1200-6840 икринок. Процесс икрометания у нее не растянут, и она задерживается в водоеме всего 6-8 дней. Личинки покидают яйцо на ранних стадиях развития, когда есть лишь хвостовая почка и зачатки неразветвленных наружных жабр, которые исчезают затем через 8 дней. Почки задних конечностей появляются на 22-й день, на 32-й день задние конечности разделяются на суставы, на 42-й появляются передние конечности. Развитие головастиков обыкновенной жабы продолжается несколько медленнее, чем зеленой (на 4- 5 дней). Медленнее они и растут. Средний прирост их за день составляет 0,6 мм, и перед метаморфозом они достигают 40% от размеров взрослых. Половозрелость наступает на 3-4-м году жизни; у самцов раньше, чем у самок. Продолжительность жизни в неволе достигает 36 лет.
Чернорубцовая, или малайская, жаба (Bufo melanostictus), обитающая в Юго-Восточной Азии,- самое распространенное земноводное, идущее и высоко в горы. В Гималаях она поднимается на высоту более 3000 м. По размерам и внешнему виду напоминает обыкновенную жабу. Верхняя сторона туловища этого животного от светлого желто-бурого цвета до черно-бурого. Головные гребни и роговые шипы, сидящие на бородавках, черные. Низ тела светлее, одноцветный или же с более темными пятнами. В период спаривания горло самца окрашено в красивый желтый цвет.
Предпочитает возделанные местности, обочины дорог, лужайки. В лесу встречается реже. Днем скрывается под камнями, пнями и в норах. Выходит оттуда лишь незадолго до заката солнца и остается вне убежищ до рассвета. Питается муравьями, термитами, а также жуками и наземными моллюсками. Собирает корм с поверхности почвы. Размножается в течение всего года, но наибольшей активности размножения достигает в ноябре - декабре, в начале сезона муссонов.
В последнее время проведены интересные исследования по биологии обитающей в Северной Америке дакотской жабы (Bufo hemiophrys). Почти вся популяция этих жаб на северо-западе штата Миннесота зимует в рассеянных по прерии земляных насыпях. Высота насыпей достигает 30-60 см, а ширина - 3-12 м. При рытье нор жабы перемещают за год 2407 м3 почвы. Возможно, их роющая деятельность была существенным фактором в происхождении этих насыпей. Массовый выход с зимовки у этого вида обычно происходит 12-18 мая. Половозрелые особи просыпаются раньше молоди, а самцы и самки одновременно. В период выхода с зимовки была поймана 7651 жаба. Все выловленные животные объединяются только в две возрастные группы - неполовозрелых и взрослых. Происходит это потому, что молодые достигают размеров взрослых уже при выходе со второй зимовки. Быстро растут они и в первый год своей жизни. После метаморфоза молодь имеет длину 31 мм. Число самцов и самок среди взрослых жаб одинаково. Соотношение молодых и взрослых меняется по годам. Так, в 1960 г. на одно взрослое животное приходилось четыре неполовозрелых, в 1961 г.- четырнадцать, а в 1962 г.- тринадцать. Зима 1960 г. была малоснежной, и почва промерзла глубже, чем обычно. Вероятно, неполовозрелые особи закапываются не так глубоко, как взрослые, и в малоснежные холодные зимы гибнут в большем количестве. Отмечены вертикальные перемещения жаб в течение зимы. При помощи мечения животных радиоактивными веществами установлен характер перемещения жаб и в активный период. Жаб с меткой четырехмесячной давности можно найти на поверхности на расстоянии 6 м, а закопавшихся - на глубине 55 см. В воде жабы со свежими метками обнаруживаются на глубине до 42 см на расстоянии свыше 15 м. Отдельные животные несколько дней держатся внутри ограниченного района и затем внезапно перемещаются в другой. Из 276 помеченных экземпляров у 84% ежедневные перемещения составляли не более 30 м, и лишь у 4% они превышали 60 м. Суточная смена мест обитания не наблюдалась.
У другой прибрежной жабы (Bufo valliceps), живущей в Техасе и Луизиане, индивидуальные участки ограничиваются расстоянием в 45 м от центра активности. Животные, перемещенные со своих мест жительства, возвращаются обратно, преодолевая при этом расстояния 36-220 м. Одна самка вернулась "домой" после того, как была перенесена на 300 м вниз по реке.
На примере иосемской жабы (В. canorus), обитающей в горах Сьерра-Невада, установлено, что жабы могут в известной мере поддерживать температуру тела на определенном уровне, греясь на солнце, когда холодно, и уходя в прохладную воду или под землю при высоких температурах окружающей среды. Животные ведут себя так, что температура их тела по возможности приближалась к верхней границе их температурного диапазона. У половины отловленных жаб температура тела была выше 24,5°, у очень немногих-выше 28,5е. Ниже 8° температура тела не была отмечена. Разница между температурой тела жабы и температурой субстрата в среднем составляет 3,2°. В 63% случаев температура тела жабы была выше или равнялась температуре почвы. В еще большей степени температура тела жабы превышает температуру воздуха.
Температура тела при различной влажности у земноводных значительно изменяется.
Так, у лопатоногой жабы (Bufo fowleri) в условиях опыта, когда температура окружающей среды была 20°, температура тела животного при 96-100% относительной влажности была меньше температуры окружающей среды на 0,74°, а при 7% относительной влажности - на 7,33°. У этого же вида изучали реакцию на свет в связи с различной температурой окружающей среды. При температуре 20-25° примерно 80% жаб двигались к свету, причем реакция эта наступала в течение 2 минут. При более высоких и более низких температурах их движения становятся медленными и менее целеустремленными, а реакция на свет замедляется.
Самая известная жаба из Южной и Центральной Америки - это ага (Bufo marinus), животное, достигающее в длину 250 мм и более. Сверху темно-бурая или светло-серая с большими темными пятнами. На более светлой нижней стороне разбросаны частые мелкие красновато-бурые пятна.
От родственных видов ага отличается формой и положением костных выступов на голове, особенно выступом, имеющим полукруглую форму и окаймляющим верхнее веко, большой, ясно заметной барабанной перепонкой и очень крупными околоушными железами.
В местах своего распространения этот вид довольно многочислен. Живет на сухих почвах, но по временам ищет себе мест, изобилующих водою, в особенности перед линькой. В отличие от всех других земноводных встречается более или менее постоянно в предустьевых солоноватых водах по побережью и на островах. Вероятно, это возможно потому, что ага имеет наиболее ороговевшую кожу и соответственно наиболее совершенно развитое легкое среди всех земноводных.
Ведет ночной образ жизни, легко передвигается при помощи прыжков. В неволе охотно поедает различных беспозвоночных, не отвергает лягушек и мышей. Одна жаба крупного размера съела в течение зимы трех крупных саламандр. Размножение начинается в июне и длится до октября. В брачный период самцы громко кричат, иногда даже днем. Личинки аги в раннем возрасте черного цвета и непропорционально малы по сравнению со взрослыми. Даже закончив превращение, молодое животное имеет длину всего 10 мм и заметно отличается по окраске от взрослых. Яд этой жабы сильнодействующий. Например, собака, схватившая жабу-агу, погибает. В Южной Америке яд аги жители употребляли для приготовления отравы для стрел.
Рис. 53. Жаба ага (Bufo marinus)
Среди представителей подсемейства настоящих жаб есть виды, резко отличающиеся по своему облику и образу жизни от других жаб.
Так, лазающая ночная жаба (Nectophryne afra) из Камеруна, как и большинство других видов этого рода, имеет на пальцах диски для прилипания, позволяющие ей легко передвигаться по листьям деревьев и даже по вертикальным стеклянным поверхностям, кроме того, она искусный прыгун. Избегает солнца, но при достаточной влажности может попадаться на глаза и днем.
Водяная жаба (Pseudobufo subasper), живущая на Малайском архипелаге, достигает 150 мм длины, внешне очень напоминает обыкновенных жаб, но у нее мощные плавательные перепонки между пальцами задних лап.
Очень интересна живородящая жаба (Nectophrynoides occidentalis), длиной 20-30 мм. Ее биология была впервые изучена в 1942 г. в Гвинее на хребте Монт Нимба (Восточная Африка). Она живет по горным лугам на высоте 900-1600 м. Жаба выбирает места, где под почвой есть породы, имеющие трещины, дающие животному убежище. Температура воздуха в местах обитания этой жабы не опускается ниже 12°. Однако при этой температуре жабы уже малоактивны и перестают кормиться. Только при 20° живородящая жаба оживленно кормится. Излюбленная ее пища - муравьи и пауки. Очень редко поедает моллюсков и червей. Период засухи с декабря по февраль животное неактивно и проводит забравшись в трещины скал или зарывшись в почву. Первыми покидают убежища взрослые самки, начинающие вести активный образ жизни с конца февраля. Затем в течение марта - апреля появляются неполовозрелые особи и наконец самцы. Это единственный живородящий вид всего отряда бесхвостых. Яйца развиваются в нижнем расширяющемся отделе яйцеводов. На свет появляется вполне сформировавшаяся жаба. Каждая самка несет от 1 до 22 зародышей, чаще их бывает 4-12. Развитие зародышей происходит за счет желтка, запасаемого в крупных яйцах. В дыхании эмбрионов, видимо, принимает участие хвост, богатый кровеносными сосудами. Массовое рождение детенышей приходится на начало июня, но продолжается до конца месяца, а если период дождей затягивается, то и до начала июля. В июле и августе наблюдается максимальная активность жаб. В конце августа часть взрослых самок перестает встречаться. Их исчезновению предшествует спаривание, начинающееся в августе и достигающее максимума в сентябре - октябре. Спаривание происходит днем, может продолжаться и ночью. Оно длится от нескольких часов до целого дня. Самцы держатся при этом неподвижно, а самки неутомимо переступают с лапы на лапу. Оплодотворенные самки сразу же прячутся в убежище, где и проводят весь период засухи. К концу периода дождей, в сентябре - октябре, а в некоторые годы и в ноябре, достигают зрелости трехмесячные самки. Оплодотворяясь, они также прячутся в убежища. Дольше всех остаются активными некоторые не достигшие зрелости самки и самцы, забирающиеся в убежища последними. Не достигшие зрелости самки оплодотворяются в следующий сезон, пятнадцати месяцев от роду. Самки, как правило, размножаются два раза в жизни и оставляют потомство в общей сложности из 18-20 особей.
Рис. 54. Африканская живородящая жаба рода Nectophrynoides и ее эмбрион
Живородящие жабы составляют значительную часть животного населения высокогорных лугов. В мае на высоте от 1400-1650 м выше уровня моря их численность достигала 150-160 особей на 100 м2. В августе и сентябре плотность их увеличивается до 400 экземпляров на 100 м2, причем молодых в это время вдвое больше, чем старых. Численность эта довольно постоянна и на протяжении 5 лет мало изменилась. Исключение составляют очень засушливые годы (например, 1946), когда численность сокращается наполовину или даже на три четверти от первоначальной.
Видимо, от австралийских жаб происходит подсемействоюжноамериканских жаб. Среди них наиболее богат видами род Eleutherodactylus - листовых лягушек, которых насчитывается до 200 видов, населяющих юг Северной Америки, Вест-Индию, Центральную и Южную Америку. В Южной Америке они занимают место настоящих лягушек, там отсутствующих. Они обитают на деревьях, кустах и в траве, а также ведут полуводный образ жизни. Родственные виды в пределах одного и того же ареала могут быть изолированы друг от друга благодаря различиям в биологии и не конкурируют между собой. Так, тропические виды листовых лягушек - Е. cornutus и Е. terraebolivarus приурочены к различным участкам леса. Первый держится в типичных лесах, второй на дорогах по краю леса, в древесных завалах и т. п. Однако и в переходных зонах, где оба вида встречаются вместе, они не конкурируют, так как Е. terraebolivarus охотится на травянистой растительности, а Е. cornutus в подстилке.
Листовые лягушки очень громкоголосы. В листве и на земле часто раздается нежный металлический голос маленькой листовой лягушки Е. abbotti. Как лай собаки, звучит крик крупной Е. inoptatus. Невзрачная и очень пугливая Е. flavescens издает пронзительный крик, напоминающий голос обыкновенной квакши.
На деревьях и в высоких травянистых растениях на Антильских островах живет Е. martinicensis. Это животное, длиною в 40 мм, имеет беловато-серую основную окраску, покрытую разнообразным рисунком из бурых пятен. Пальцы передних и задних конечностей у нее заканчиваются пластинками для прилипания. Антильская лягушка откладывает в пазухи листьев 15-30 круглых прозрачных яиц, диаметром 4-5 мм. Кладка заключена в оболочку. Яйцеклетка беловатая или бледно-соломенного цвета. Возможно, что самка держится вблизи кладки. Все развитие происходит внутри яйца, и примерно через 14 дней после их откладки из них выходят совершенно закончившие развитие молодые животные, достигающие в длину всего 5-7 мм. Развитие зародыша происходит в пузыре, заполненном жидкостью. Большой хвост тесно прилегает своей широкой поверхностью к внутренней стенке пузыря, богат кровеносными сосудами и служит органом дыхания.
Листовая лягушка Е. abbotti кладет свои зеленые яйца также на листья. На суше же откладывает яйца живущая на?
Кубе Е. ricordii, отличающаяся красивой окраской. Основной фон у нее красноватый. По бокам тянутся две светлые полосы, а на спине неравномерно разбросаны пятна. Размеры ее невелики - до 30 мм. Е. karlschmidti - наиболее водный вид из представителей этого рода на острове Пуэрто-Рико. Среди всех островных бесхвостых амфибий она уступает по величине только are, достигая 80 мм длины. Самцы имеют хорошо развитые голосовые мешки и ночью громко кричат. Несмотря на полуводный образ жизни, большую часть пищи этот вид собирает на суше. Среди водных животных 91% составляют формы, живущие на дне водоемов. В питании Е. karlschmidti встречаемость насекомых равна 91%, пауков - 5,3%, клещей - 4,1 % , ракообразных 1,45% , многоножек - 1,03% и моллюсков - 0,83%.
Из представителей других родов широко известна рогатка (Ceratophrys cornuta), имеющая плотное жабообразное тело, чрезвычайно большую и широкую голову и соответствующий ей по величине рот. Язык, как у лягушек, с глубоким вырезом сзади. Свое название рогатки получили за своеобразные заостренные выросты над глазами, представляющие собой вытянутое в высокое острие верхнее веко. Высокие бородавчатые гребни на окостеневшей коже головы как бы повторяют это образование. Рогатка достигает 150-200 мм в длину. Она ярко окрашена. Широкая полоса, проходящая от конца морды через всю спину, у самца оранжево-красная, а у более крупной самки ярко-зеленая. Остальные части тела сверху окрашены в красно-бурые, черно-бурые и зеленоватые тона, располагающиеся в виде пятен и полос. Брюхо посредине желтовато-белое, по бокам желтое и усеяно красно-бурыми пятнами. В окраске наблюдаются значительные вариации. Закопавшееся в землю животное сидит так, что наружу выдается только большая голова, совершенно сливающаяся с окружающим фоном. В таком положении рогатка поджидает добычу, которой ей служат не только беспозвоночные, но и другие животные, например бесхвостые земноводные и мелкие зверьки. Живет в сырых лесах.
Из этого подсемейства два вида могут претендовать на особую славу благодаря тому, что у них самые крупные из всех известных головастики. Это псеудис, или удивительная лягушка (Pseudis paradoxa), живущая в Гвинее и ведущая водный образ жизни. Взрослое животное имеет длину 19-69 мм; верхняя сторона его оливково-зеленая с бронзовыми светло-зелеными и черными пятнами, нижняя сторона тела желтая с бурыми пятнами и такими же полосами на нижней поверхности задних ног. Внутренний палец передней ноги может противопоставляться другим. Личинка ее имеет 250 мм в длину.
Немногим меньше личинка у шлемоголовой лягушки (Calyptocephalus gayi), живущей в водоемах Чили. Она достигает 150 мм длины, тогда как длина взрослого животного равна 200 мм.
Подсемействоносатых жаб, видимо, произошедшее от южноамериканских жаб, заключает в себе всего один вид - носатую жабу (Rhinophrynus dorsalis). Тело носатой жабы почти яйцевидно-округлое, голова сливается с ним и вытянута наподобие небольшого хобота. Конечности у нее пухлые, в области плюсны на подошве выступает роговой имеющий вид лопаты бугор. Основная равномерно-бурая окраска разнообразится проходящей вдоль спины желтой полосой и несколькими боковыми пятнами такого же цвета. Длина тела до 85 мм. Она хорошо роет и питается исключительно термитами, слизывая их своеобразным языком, который сзади прирос и подвижен только на конце.
Также от южноамериканских жаб произошло подсемействосвистунов (Leptodactylinae). Представители его сильно варьируют по величине - от 23 до 200 мм.
Глазчатый свистун (Leptodactylus ocellatus) - стройное животное длиной в 90-110 мм. У него совершенно отсутствуют плавательные перепонки и чрезвычайно толсты передние конечности. Глазчатый свистун очень обыкновенен в тех местах, где он обычно встречается, держится всегда поблизости от воды, чтобы иметь возможность тотчас же укрыться в ней, если его спугнут. Днем глазчатый свистун скрывается в лужах, болотах и стоячей воде, при наступлении вечерней прохлады или в дождливую погоду он покидает свои убежища и быстро и ловко передвигается в траве, делая удивительно большие для своей тяжеловесности прыжки. В это время можно слышать его голос, напоминающий свист, которым подзывают собаку. В период спаривания самцы издают другой, отрывистый звук, похожий на удары топора, когда отесывают бревно. Глазчатый свистун строит в водной растительности гнездо, похожее на взбитые белки. Полость этого гнезда сквозная, и животное сидит в нем глубоко в воде. Размножается он в сентябре - октябре.
Другой вид - чакский свистун (L. chaquensis) живет в засушливой зоне Чако и отличается относительно беспрерывным циклом образования половых продуктов.
Пятипалый свистун (L. pentadactylus) характеризуется большими размерами (200 мм). Он привлекает внимание красивой красной окраской задней стороны бедер. Широко распространен по Южной Америке.
Среди свистунов есть виды, развитие которых хотя бы частично происходит на суше. Так, L. prognatus, обычный в окрестностях Буэнос-Айреса, откладывает икру в небольших ямках, вырытых самцом. Развитие икры и выход головастиков приурочены к периоду дождей. Эмбриональное развитие длится 5 дней. Головастики, обитающие в воде, питаются диатомовыми, сине-зелеными водорослями, корненожками, коловратками и растительными остатками. На 68-й день после откладки икры молодое животное достигает размеров взрослых (30 мм). Неблагоприятное время года этот вид проводит во влажных местах под стволами деревьев.
Усатый свистун (L. mystacinus), наиболее многочисленный из свистунов в Бразилии и Парагвае, никогда не идет в воду. Икру откладывает поблизости от луж в пределах того пространства, которое покрывается водой после сильных ливней. Там он выкапывает под камнями или гниющими стволами деревьев ямки и наполняет их своей икрой, заключенной в пенистую массу наподобие взбитого белка. В центре этой пенистой массы находятся бледно-желтые яйца, которые развиваются здесь в головастиков. Если вода в луже поднимается до гнезда, личинки переходят в нее. Если же вследствие засухи мелкие лужи высыхают, личинки прячутся под древесные стволы, листья и ожидают там возобновления дождей, собравшись в комок.
Некоторые виды другого рода из этого же подсемейства - болотницы (Paludicola) напоминают по внешнему виду наших чесночниц. Их также характеризует сильно развитый пяточный бугор. На бедрах взрослых особей имеется по яйцевидной кожной железе, которая у чилийской болотницы (P. bibroni) украшена черно-белым рисунком. Размеры этих животных невелики - от 15 до 41 мм. Образ жизни полуводный. Другой вид - P. fuscomaculata издает, попеременно надувая то свой горловой мешок, то брюхо, похожий на мяуканье котенка крик, который регулярно можно слышать около мелких болот. Пищей этому животному служат главным образом водяные жуки. Бесцветные мелкие яйца, диаметром в 1 мм, откладываются в январе в пенистую плавающую на воде массу. Некоторые болотницы откладывают яйца на дне высыхающих луж, и головастики развиваются вне воды.
Семейство Квакши (Hylidae)
Семейство квакш - одно из самых обширных семейств, 416 видов которого объединены в 16 родов. Населяет Европу, Юго-Западную и Юго-Восточную Азию, Северную Африку, Австралию и прилежащие острова, Южную и Северную Америку. Громадное большинство земноводных, относящихся к этому семейству, имеет на концах пальцев расширенные диски, способствующие прилипанию животного при передвижении по вертикальным плоскостям. Диски эти богаты лимфатическими пространствами и слизистыми железами. Прикрепление к субстрату тем сильнее, чем меньше воздуха между ним и диском. Особые мышцы дают возможность дискам становиться более плоскими и плотнее прижиматься к поверхности, по которой движется животное. Обыкновенно прикрепление к субстрату идет также за счет кожи брюха и горла.
Род Hyla, самый обширный по числу видов из всех родов в классе земноводных, принадлежит именно к этому семейству. Он объединяет собственно квакш, или древесниц, относящихся к 350 различным видам самых разнообразных размеров - от 17 до 135 мм. Представители его распространены во всех частях света, кроме тропической Азии и Африки. Особенно многочисленны квакши в Южной Америке и Австралии.
Австралийская белая квакша (Hyla coerulea) достигает длины 105 мм. Она отличается широкой головой, короткой, тупой мордой, толстыми, особенно передними, лапами и большими подушечками на пальцах. Верхняя сторона тела ярко-травянисто-зеленая, нижняя - беловатая, за исключением внутренней стороны бедра, которая окрашена в бледно-буровато-красный цвет. Радужная оболочка красивого золотого цвета. У многих животных на верхней, зеленой стороне образуется рисунок из белых полос, идущих от края верхней губы под глазом до барабанной перепонки, более коротких полос по плечам и длинных по наружной поверхности предплечья и по заднему краю голени. На плече и по бокам туловища встречаются ярко-белые пятна, часто с темной каймой. Голос самца слышится с апреля по сентябрь и напоминает отдаленный лай раздраженных собак. Голос самки звонче и пронзительней, чем у самца. В неволе животное очень прожорливо, поедает не только беспозвоночных, но и разнообразных рыб, других земноводных, мелких ящериц, мышей и птиц. С ночной охоты квакша возвращается на одно и то же место, где сидит весь день, неподвижно сложив передние лапы под горлом. Яйца откладываются в пенистую массу.
На Новой Гвинее и прилежащих островах живет еще более крупная квакша - Hyla dolichopsis - длиной 120 мм. От своих австралийских сородичей она отличается еще большими подушечками на пальцах, большими плавательными перепонками и очень длинными задними ногами.
По всей Австралии и Тасмании живет золотистая квакша (Hyla aurea), внешне очень напоминающая водяную лягушку, 80-85 мм длиной. Подушечки на ее пальцах очень малы, и по бокам тела тянутся продольные валики, характерные для лягушек. В неволе она, подобно лягушкам, охотно сидит в воде и не лазает, как все квакши, по вертикальным поверхностям. Сверху у этой квакши по красивому зеленому фону бывают разбросаны блестящие золотые пятна или имеется золотой отлив. Внутренняя сторона бедер и голени имеет яркую зелено-синюю окраску. От ноздри к глазу тянется черная полоса, бока светло-бронзово-блестящие с темным сетчатым рисунком, брюхо белое, радужная оболочка великолепного золотистого цвета. Окраска животного может меняться чрезвычайно быстро. Сидящая в воде квакша бывает совершенно черной. Яйца откладывает в августе и сентябре в воду в виде белых пенистых комков.
На Дальнем Востоке, в Японии, Корее и Северном Китае живет единственный азиатский представитель этого рода - дальневосточная квакша (Н. japonica).
По внешнему виду очень похожая на нее обыкновенная квакша (Н. arborea) распространена от Средиземноморья через Западную Европу, юго-запад Европейской части СССР, Украину, Крым, Кавказ и Малую Азию до Ирана включительно. В пределах нашей страны на восток известна до Дона, на северо-восток до Воронежа, Орла, Тулы; на запад до Белоруссии и Южной Прибалтики. В горы поднимается до 1500 м.
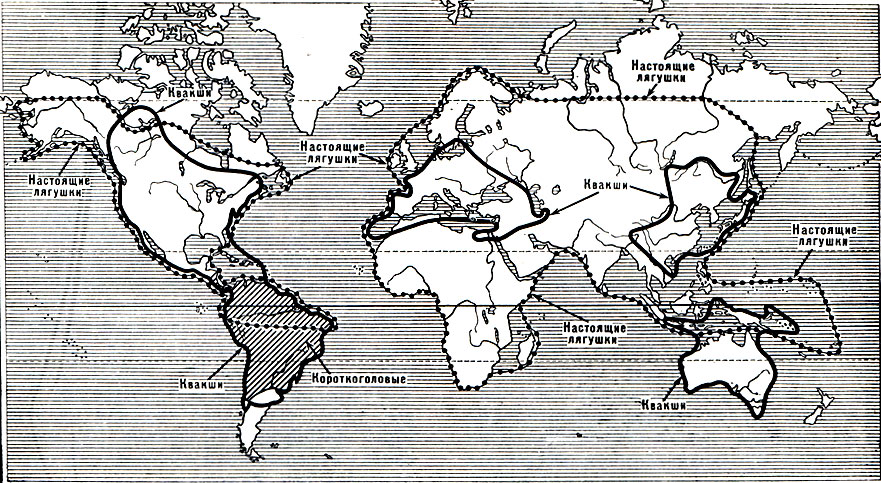
Рис. 57. Области распространения семейств квакш, настоящих лягушек, короткоголовов
Размеры обыкновенной квакши - 35-45 мм. Сверху она окрашена в ярко- зеленый цвет, а внизу в желтовато-белый. Верх отделяется от низа тонкой, расширяющейся сзади черной полосой, имеющей сверху белую каемку.
Окраска квакш очень изменчива. Встречаются молочно-белые, лимонно-желтые, сиреневые, серые, почти черные и, наконец, пятнистые экземпляры. Это изменение окраски, очевидно, связано с действием температуры и влажности. При понижении температуры и увеличении влажности животные темнеют. Однако и в совершенно одинаковых природных условиях можно одновременно встретить квакш совершенно различной окраски.
Большую часть жизни древесницы проводят на суше, забираясь на деревья, кусты и высокие травянистые растения.
Здесь они полностью сливаются с зеленым фоном, на котором держатся, и их бывает нелегко заметить. Однако далеко не всегда квакши окрашены под цвет субстрата, на котором они сидят. Эти животные обладают одинаковой способностью передвигаться как в воде или на суше, так и среди листвы деревьев. В умении плавать они мало уступают водяным лягушкам, а в умении прыгать и лазать далеко превосходят их.
На протяжении почти всего теплого времени года и днем и особенно ночью можно слышать громкие голоса древесниц, напоминающие утиное кряканье. Однако день квакши проводят неподвижно, схватывая лишь пролетающую мимо добычу. На охоту они выходят только в сумерки. При этом, если есть возможность, животные сначала спускаются в воду, купаются, пополняют израсходованный, за день запас влаги в теле и затем начинают кормиться.
В питании квакш 96% составляют наземные корма. Это блошки (Halticini), щелкуны (Elateridae), различные листоеды (Chrysomelidae), гусеницы и муравьи. Значительный процент приходится на летающих насекомых (17,6%). При ловле добычи квакши, подобно лягушкам, выбрасывают длинный липкий язык, бьющий по жертве. При захватывании крупной добычи квакши запихивают ее в рот при помощи передних лап.
На Северном Кавказе квакша уходит на зимовку в конце сентября - в октябре. Зимует она в щелях каменных сооружений, под листьями и мхом у подножия деревьев, в дуплах, норах, под камнями и в иле на дне водоемов. Появляются квакши в начале апреля. Сначала просыпаются самцы, а самки выходят лишь 6-8 дней спустя. С середины апреля начинаются громкоголосые концерты. Квакши предпочитают водоемы с берегами, окаймленными тростником, кустами и деревьями. Спаривание и икрометание начинается в конце апреля и длится весь май. В горах икрометание запаздывает примерно на месяц. Самец обхватывает самку, подобно жабам, под мышками. Брачные мозоли у квакш развиты слабо. Икра откладывается порциями, в виде комков, на протяжении 2-48 часов. Одна самка может отложить 800-1000 икринок. Диаметр яиц - 1-1,5 мм, а вместе с оболочками - 4 мм. Яйца квакши, как и яйца повитухи, могут быть оплодотворены на суше и способны долго противостоять высыханию. Нормально икра развивается на дне водоемов. Как на свободе, так и в искусственных условиях квакши способны использовать для откладки икры небольшие скопления воды в пазухах листьев некоторых растений и в дуплах. Светлые личинки вылупляются из яиц примерно на 10-й день, длина их 5 мм. В небольших скоплениях воды развитие яиц и головастиков замедляется до появления задних ног, а затем сильно ускоряется. Однако общая продолжительность развития остается несколько большей, чем в нормальных условиях. Обычно на 4-й день после вылупления у личинок квакши появляются наружные жабры, которые коротки, не ветвятся и скоро исчезают. Если икра отложена прямо на влажную землю, то личинки вылупляются уже без наружных жабр или с недоразвитыми жабрами. Примерно на 51-й день у нормально развивающихся головастиков вырастают задние ноги. Головастиков квакш легко отличить по хорошо развитому, веслообразному, сильно заостряющемуся к концу хвосту, кожная каемка которого по спине проходит вперед вплоть до глаз. Глаза у них сильно сдвинуты набок. Перед метаморфозом головастики составляют около 125% от размеров взрослых. Метаморфоз наступает примерно на 90-й день. Известны случаи зимовки личинок. Половозрелость наступает на 3-4-й год. В неволе продолжительность жизни до 22 лет.
Велико видовое разнообразие квакш в Новом Свете. Многочисленные североамериканские виды по образу жизни немногим отличаются от обыкновенной квакши. Наиболее известная среди них изменчивая квакша (Hyla versicolor), достигающая 53 мм длины. Этот вид часто встречается в восточных и северных штатах от Канады до Мексики. Изменчивая квакша отличается несколько бородавчатой кожей и ярко-желтыми с грубой темной сеткой бедрами. Обыкновенно светло-серая основная окраска верхней ее стороны может переходить в темно-бурую или от почти белого цвета в красивый зеленый. Ярко выраженная способность менять окраску и отражена в названии животного.
Королевская квакша (Hyla regilla), обитающая на западе Северной Америки, обладает также несколько бугорчатой кожей и необычайно изменчивой окраской; серые, коричневые, зеленые и даже ярко выраженные розово-красные тона могут встречаться у нее во всех оттенках. Между глазами королевской квакши, как правило, помещается V-образное пятно, напоминающее корону.
В восточной части Северной Америки от Канады до южной Каролины обитает маленькая свистящая квакша (Н. crucifer), известная своим голосом, который действительно походит на свист. Калифорнийская квакша (Н. californiae) живет только у воды, редко удаляется от нее дальше чем на несколько метров. Часто по нескольку штук они забираются в щели, углубления в камнях на берегах ручьев и прудов. Много дальше, чем взрослые, до 70 м, уходят от воды молодые. Интересно, что при температурах ниже 25° тело квакши несколько теплее окружающей среды, при более высоких температурах - холоднее. Высыхая, эта квакша может потерять без смертельного исхода до 35% от первоначального веса тела. Калифорнийская квакша активна с середины февраля до начала октября. Питается пауками и насекомыми. Спаривание отмечено с середины апреля до конца июня. Головастики в водоемах наблюдались с конца апреля до конца августа.
В Мексике в период дождей очень часто можно видеть сонорскую квакшу (Hyla eximia). Это маленькая квакша с заостренной головой и широкой спиной, отчего форма ее тела оказывается грушевидной. Сверху зеленая с темными пятнами, расположенными в два, иногда в четыре продольных ряда. Брюхо серовато-желтое, горло темное, часто черное. Она населяет все открытые, поросшие травой места, если там есть хоть маленькие лужицы. Особенно любит сильно заросшие водоемы, в прибрежных частях которых, погрузившись в воду так, что видна только голова, лежат и кричат самцы. Их голоса звучат со второй половины дня до глубокой ночи. Сонорскую квакшу не находили на деревьях. В безветренных уголках она греется на солнце, сидя на листьях агав и кукурузы. В разгаре размножения ее икра встречается в каждой луже и даже в заполненных дождевой водой колеях малопроезжих дорог. Кладка содержит самое большее 25 яиц, которые, как жемчужный венок, окружают стебли растений. Уже на 5-й день после откладки икры вылупляются личинки. Так как в Мексике вода испаряется очень быстро, то много яиц и личинок гибнет.
На Кубе, Гаити и Багамских островах живет доминиканская квакша (Hyla dominicensis). Длина тела самки у этой квакши может превышать 100 мм. У этих квакш неуклюжее тело, невероятно широкая голова, бородавчатая кожа, большие выпученные глаза. Кожа головы прирастает к черепу и неподвижна. Значительно меньший самец имеет два голосовых мешка, расположенных не под горлом, как у других квакш, а по углам рта. На внутренней стороне первого пальца передних конечностей в период размножения развиваются черные брачные мозоли. Окраска самки сверху, за исключением чисто-бурой головы, желтовато-серая с темным расплывчатым мраморным рисунком. На передних и задних ногах отчетливые поперечные полосы. Брюхо без пятен, беловатое. Самец в брачном наряде темно- коричневый с бронзовым отливом и яркими зелеными пятнами. В неволе, кроме насекомых, поедает и мышей. Активна ночью. Когда животное испугано, кожа его покрывается беловатыми выделениями, раздражающими слизистые оболочки.
Поражают разнообразной красивой окраской квакши Южной Америки. Удивительно красива бразильская жемчужная квакша (Hyla albomarginata), достигающая 70 мм длины. Спина и горло у нее голубовато-зеленоватые. Брюхо чисто белое. На спине у одних животных имеются белые, подобные жемчужинам пятна, у других - черные точки. Глазное яблоко ультрамариновое с черным зрачком, окруженным серебристой радужной оболочкой. Пальцы задних ног оранжево-желтые. Эта квакша - обитатель открытых пространств - мечет икру в стоячих водоемах, берега которых порастают травой. Даже в садах из прудов для золотых рыбок звучит ее напоминающий птичий голос.
Многочисленный обитатель южноамериканских болот - двупятнистая квакша (Hyla bipunctata) имеет длину тела всего 25 мм. Спина этой квакши кирпично-красная, брюхо лимонно-желтое, нижняя сторона задних лап яркая, кораллово-красная. Весь день она проводит в убежище, сидя головой вниз. Еще причудливее окраска квакши Н. leucophyllata. Спина ее окрашена в красновато-темно-каштановые тона. На этом фоне вырисовывается своеобразная фигура, имеющая серебристо-белый или золотистый цвет. Она представляет собой два треугольника, один из которых располагается впереди глаз и направлен вершиной к кончику морды, другой, больший, лежит в задней трети спины, и вершина его обращена назад. Оба эти треугольника соединены широкими полосами, идущими по бокам тела животного. Другой вид - Н. crepitans из Венесуэлы - стройная квакша, в два раза большая, чем наша обыкновенная древесница. Днем она имеет сверху почти белую окраску, с рисунком, напоминающим поблекший лист, ночью эта квакша может становиться светло-каштаново-коричневой. Снизу она оранжево-желтая. Самое красивое в ней - большие сине-зеленые глаза. Голос ее напоминает шум, который производят весла при погружении в воду.
Квакша Геслери (Hyla giesleri) отличается сходством с лишайниками, очень бородавчатой кожей и интенсивной оранжевой окраской нижней стороны задних лап.
Днем в сухом воздухе она бежевая. Красная квакша (Hyla rubra), обитающая в Боливии, Парагвае и Южной Бразилии, красивого светло-красно-коричневого цвета, с треугольным пятном между глазами, вершина которого направлена вперед. Бока, пах, а также передняя и задняя сторона бедер серо-желтые или желтоватые с черным мраморным рисунком. Конечности опоясаны поперечными полосами. Брюхо беловатое, горло иногда испещрено коричневыми крапинками. Очень влаголюбива. На стволах деревьев сидит вниз головой. Прекрасный прыгун.
Самая маленькая квакша - Hyla emrichi имеет всего 17-18 мм длины. Живет она на кустарниках, голос ее звучит пронзительно и напоминает звук, получающийся, когда по фарфору проводят ножом. Несмотря на свои маленькие размеры, эта квакша может совершать прыжки до 75 см длиной. Другая, чернопятнистая квакша (Н. nigromaculata), имеющая длину около 30 мм, прыгает на 150 см. Удивительно хорошо сливается с окружающим фоном Н. senicula. Ее бородавчатая верхняя сторона, похожая по окраске на кору деревьев или лишайников, и волнистая складка кожи на предплечье и задних конечностях делают контуры животного совершенно незаметными. Эта квакша издает своеобразный, не лишенный приятности запах. Интересна приспособительная окраска и поведение у очень спокойной географической квакши (Н. geographica). При опасности она прижимается к подстилке с несколько втянутыми глазами и закрывает верхнее веко, которое покрыто светло-коричневыми жилками. Благодаря этому глаз становится неотличимым от тела, окрашенного в глинистый цвет с темным рисунком, напоминающим географическую карту. Все затаившееся животное похоже на засохший лист. Некоторые квакши, когда им грозит опасность, переворачиваются на спину и притворяются мертвыми, другие спасаются при помощи мгновенных длинных прыжков в быстро меняющихся направлениях. Квакши большинства южноамериканских видов в период покоя сидят на нижней стороне листьев, спасаясь здесь от врагов и прямых лучей солнца, другие прячутся в узких щелях между листьями бромелии и во влагалищах ее листьев. Однако некоторые виды, как и наши обыкновенные квакши, держатся открыто на листьях и стеблях.
Среди южноамериканских квакш наблюдаются и очень интересные формы заботы о потомстве. Широко известна в этом отношении квакша-кузнец (Hyla faber), достигающая 90 мм длины. Она встречается в Северной Аргентине и в Бразилии. Кузнецом эта квакша названа за свой громкий крик, напоминающий удары молотка по медному подносу. Днем верхняя сторона тела квакши-кузнеца глинисто-желтая, вдоль середины спины проходит черная полоса. Окраска дополняется отдельными неправильными тонкими черными черточками и точками. На верхней стороне конечностей неясные темные поперечные полосы. Пах охристо-желтый, так же как и грубозернистая кожа брюха. В темноте окраска квакши-кузнеца становится красно-коричневой. Это настоящее ночное животное с большими выпуклыми глазами.
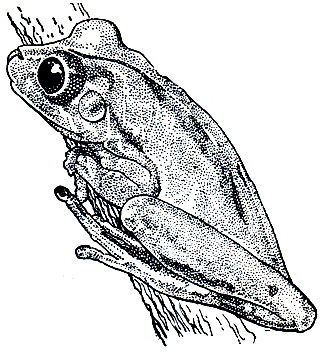
Рис. 58. Квакша-кузнец (Hyla faber)
Спящая днем квакша не закрывает век, а чрезвычайно суживает зрачок, защищая таким образом сетчатку от света. Ее любимая пища - ночные бабочки. В феврале квакша-кузнец строит в водоеме кольцеобразный вал из ила, внутри которого оказывается маленький замкнутый бассейн, куда и откладывается икра. Сооружение имеет 30 см в диаметре и 10 см в высоту. Строит только самка, самец сидит при этом у нее на спине, совершенно безмолвно. Кричащие в это время самцы еще не имеют самок. Когда самка строит кольцевой вал, то на поверхности воды время от времени появляется голова и верхняя часть тела строителя, нагруженного илом. Внутренние стенки вала квакша старательно сглаживает при помощи брюха, горла и передних конечностей, отличающихся сильным развитием дисков на концах пальца. Эти диски прекрасно действуют наподобие лопаточек. Работа происходит исключительно ночью. Вал сооружается в течение двух ночей, и сразу же или на протяжении 4-5 дней после окончания постройки в нее откладываются яйца. Через 4-5 дней вылупляются головастики, развивающиеся затем в маленьких искусственных водоемах. В террариуме наблюдалась постройка гнезд самцом, который делал это с необычайным усердием и сейчас же принимался за новое сооружение, если старое разрушалось. Закончив постройку, он сидел на ней и усиленно квакал всю ночь. Не готовые к икрометанию самки не участвуют в в постройке.
Ярко окрашенная, живущая на высоких деревьях в глубине девственных лесов бразильская квакша (Hyla resinifictrix) вымазывает дупло при помощи передних конечностей смолой. Сделавшись таким образом непроницаемым для воды, дупло скоро наполняется дождевой водой, и в него откладываются яйца. Банановая квакша (Н. nebulosa) укрепляет комки яиц вдоль края и на внутренней поверхности банановых листьев, где даже в жаркую пору сохраняется достаточная прохлада и влажность. В пенистом комке видно, как движутся личинки, которые при перенесении их в воду быстро гибнут.
В Америке не только велико разнообразие видов из рода Hyla, но встречается и еще 15 других родов семействаквакш, представленных в общей сложности 66 видами.
Самка квакши Гёльди (Flectonotus goeldii) носит комки крупных яиц на спине до тех пор, пока из них не выйдут молодые животные с четырьмя конечностями и длинным несуразным хвостом.
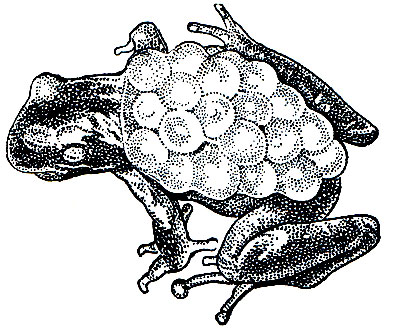
Рис. 59. Самка квакши Гёльди (Flectonotus goeldii) с кладкой яиц на спине
Квакши из рода филломедуз (Phyllomedusa) обитают в Центральной и Южной Америке (22 вида). Сверху они всегда окрашены в зеленый цвет. Те же части тела, которые бывают не видны в сидячем положении, часто яркие: оранжевые, красные и фиолетовые. Филломедузы обычно бывают худы и выглядят изголодавшимися. У всех у них короткий тупой нос и огромные глаза, которым беловатая радужная оболочка придает таинственное выражение. Ночью глаза кажутся совершенно черными из-за широко раскрытых зрачков. Все филломедузы имеют настоящие хватательные лапы, благодаря тому что первый палец передних и задних конечностей может противопоставляться остальным. По ровному месту они бегают, как жабы, шагом, но на вытянутых ногах, так что брюхо приподнято на 1-2 см от земли. В воду по собственной воле они не идут, очень беспомощны в ней и стремятся как можно скорее выбраться. Плавательные перепонки развиты слабо или вовсе отсутствуют. Слабо развиты и подушечки на концах пальцев, способствующие прилипанию. Живут они в кроне высоких деревьев и прекрасно лазают по тоненьким веточкам и листьям. Они способны прыгать, но прыжки для них нехарактерны. По особенностям перемещения филломедузы напоминают хамелеонов. Движения их медлительны, плавны и осторожны. Долго щупает филломедуза передней ногой воздух, пока не найдет ветку, за которую ухватится, затем животное подтягивает противоположную заднюю ногу и вновь тянется вперед другой передней лапой. Оторвать филломедузу от ветки невозможно, не повредив ей ногу. Все представители этого рода ведут ночной или сумеречный образ жизни. Неподвижная филломедуза схватывает добычу быстрым броском длинного липкого языка. Даже для откладки икры эти животные не идут в воду. Она заворачивается в лист или откладывается между двумя или несколькими листами, склеивающимися между собой благодаря липким оболочкам яиц. Так, готовая к откладке яиц самка Ph. hypochondrialis, несущая на своей спине самца, взбирается на лист, свисающий над водой. Затем самец и самка придерживают задними ногами сближенные края листа. В образовавшуюся таким образом трубочку самка откладывает яйца, а самец оплодотворяет их. Затем животные отползают немного дальше, и так продолжается до тех пор, пока весь лист не заполнится икрой. Примерно 100 штук отложенных одной самкой яиц помещаются в двух листьях. Яйца очень крупны и богаты желтком. Развитие совершается быстро. На 3-й день у зародыша появляются наружные жабры, на 5-й они достигают наибольшего развития, а ко времени вылупления головастика атрофируются. Вылупившийся прозрачный, как стекло, головастик, на котором видны только очень большие зеленые глаза с металлическим отливом, падает в воду, где и завершает свое развитие. Спустя шесть недель после откладки икры личинка достигает 80 мм длины; сверху она блестящего зеленого цвета, снизу серебристая, розово-красная. К концу превращения молодое животное достигает уже 2/3 длины своих родителей. Размножаются в январе.
Представители другого рода сумчатых квакш (Gastrotheca, 15 видов) по внешнему виду мало чем отличаются от настоящих квакш (Hyla), но им свойствен крайне интересный способ заботы о потомстве. Самки этого вида имеют на спине особый кожный карман - выводковую сумку, в которой вынашивают яйца.
У карликовой сумчатой квакши (Gastrotheca pygmaeum), достигающей всего 28 мм длины, 4-7 крупных яиц полностью развиваются в сумке. Ее выводковая сумка открывается длинной щелью, проходящей вдоль спины животного. Создается впечатление, что этот мешок возник в результате разрастания продольных кожных складок до соединения их в конце концов по средней линии. Такие кожные складки имеются, например, у некоторых видов рода Hyla, а у квакши Гёльди между ними располагается комок икры, вынашиваемый самкой на спине. Так же помещается икра и у скрытожаберника (Cryptobatrachus evansi), относящегося также к семейству квакш. У других видов сумчатых квакш выводковая сумка сообщается с внешней средой лишь небольшим отверстием в области крестца. Так, у обыкновенной сумчатой квакши (G. marsupiata) кладка содержит около 200 яиц величиной с горошину. В сумке из яиц развиваются головастики, которые последние стадии развития проходят в воде. Головастики покидают сумку, когда наружные жабры у них исчезают, появляются задние конечности, но сохраняется еще большой веслообразный хвост с густой сетью кровеносных сосудов. Вероятно, он служит головастику дополнительным органом дыхания. К моменту выхода личинки в воду весь запас желтка оказывается уже израсходованным, В неволе у этой квакши спаривание наблюдалось 15 апреля и 16 сентября. Спаривание длится 16-36 часов. Во время откладки яиц клоака выворачивается на 2 а и загибается на спину самки к отверстию сумки, У самки, спаривавшейся 16 апреля, головастики стали покидать сумку и выходить в воду 30 июня. Выход их продолжался 72 часа и происходил только ночью. В первую ночь появилось 8-9 головастиков, во вторую - 120-130 и в третью 8-9. Метаморфоз у этих головастиков начался 4 сентября.

Рис. 60. Сумчатая квакша Gastrotheca marsupiata
Другая сумчатая квакша - G. oviferum - откладывает всего 15 яиц, очень богатых желтком и достигающих 10 мм в диаметре. Все развитие целиком происходит в выводковой сумке. У головастиков этого вида, совершающих развитие внутри яйца, развиваются наружные жабры своеобразной формы. Они более всего напоминают цветок вьюнка, укрепляющийся на двух тонких длинных стебельках. У зародыша, развивающегося в яйце, они прижаты к телу. Богатая сеть кровеносных сосудов в этих органах свидетельствует об их значении как органов дыхания. Присутствие в стебельках этих жабр пучков поперечнополосатых мышц свидетельствует о том, что они сформировались тогда, когда животное могло ими произвольно двигать, видимо покидая на определенных стадиях развития яйцо
Таблица 11. Бесхвостые земноводные: 1, 2 - рогатка венесуэльская (Ceratophrys cornuta); 3 - южноафриканский узкорот (Breviceps adspersus); 4, 5 - остромордая лягушка (Rana terrestris), сверху - самец в брачном наряде; 6 - лягушка сибирская (Rana cruenta); 7 - лягушка травяная (Rana temporaria); 8 - лягушка озерная (Rana ridibunda); 9 - лягушка прудовая (Rana esculenta)

Таблица 11. Бесхвостые земноводные: 1, 2 - рогатка венесуэльская (Ceratophrys cornuta); 3 - южноафриканский узкорот (Breviceps adspersus); 4, 5 - остромордая лягушка (Rana terrestris), сверху - самец в брачном наряде; 6 - лягушка сибирская (Rana cruenta); 7 - лягушка травяная (Rana temporaria); 8 - лягушка озерная (Rana ridibunda); 9 - лягушка прудовая (Rana esculenta)

Таблица 12. Бурые лягушки: Остромордая лягушка (Rana terrestris)

Таблица 12а. Бурые лягушки: Травяная лягушка (Rana temporaria) в природной обстановке
Выводковой сумкой на спине обладает еще одна квакша - Amphignathodon guentheri из Эквадора.

Рис. 61. Сумчатая квакша Gastrotheca oviferum
Семейство Короткоголовые (Brachycephalidae)
Короткоголовые (10 родов) распространены на юге Центральной Америки и Больших Антильских островах. Большинство из них мелкие (18-76 мм) ярко окрашенные животные, напоминающие квакш.
Короткоголовые, относящиеся к роду древолазов (Dendrobates), насчитывающему 10 видов, отличаются большой ядовитостью кожных выделений и интересной формой заботы о потомстве. Слизь этих животных употребляется туземцами для изготовления отравленных стрел.
Красящий древолаз (Dendrobates auratus) с Коста-Рики известен особенностью действия своих кожных выделений. Если кожу молодых попугаев слегка смочить слизью из кожи этого древолаза, то вместо зеленых перьев у попугая вырастают желтые или красные. Красящий древолаз зеленый, с крупными, правильно расположенными черными пятнами. Плавательные перепонки у него отсутствуют, на концах пальцев есть диски для прилипания. Задние ноги относительно тонки и коротки. Он ведет дневной образ жизни, любит полутень, влагу и тепло. Животное это целый день в движении. Питается только мелкими мухами. Яйца откладывает на влажную почву, и самец остается рядом с ними. Выведшиеся из икры головастики прикрепляются к его спине и переносятся в воду, где и заканчивают развитие. Самый маленький древолаз - D. pumilio имеет длину всего 18 мм. Сверху это животное блеклого оранжево-красного или красного тона с точечными пятнышками. Другой вид - D. typographicus, длиной в 20 мм, блестит, как лакированный, и напоминает тончайший красный фарфор. Конечности у него темно-сине-серые, на спине тонкие точечные штрихи. Глаза овальные, большие, черные. Активен днем. Кладка яиц с грецкий орех величиной.
У листолаза (Phyllobates trinitatis) из Венесуэлы проявляется такая же забота о потомстве, как и у красящего древолаза. Этот обитатель влажных лесов имеет в длину 25 мм и окрашен в серо-оливковые или коричневатые тона с темными пятнами. Низ у самок лимонно- или золотисто-желтый. Этот листолаз также активен днем и охотится на маленьких насекомых, пауков и червячков.
Род короткоголовое (Brachycephalus) представлен всего одним видом, обитающим в Гайане, Гвиане (Фр.)" Суринаме (Нид.), Бразилии и Чили. Эти напоминающие по внешнему виду жабу крошечные животные 15 мм длины окрашены в ярко- желтый цвет. Живут среди опавшей листвы и на низких бромелиях. Питаются тлями и комарами. Их еще называют седлоносными жабами, так как на спине у них имеется костная пластинка, крепко срастающаяся с кожей и остистыми отростками позвонков.
Самый большой род этого семейства - ателопы (Atelopus) содержит 26 видов. Ателоп Стелъцнера (A. stelzneri) имеет 20 мм длины, сверху по матово-черному фону идут желтые пятна, ладони и ступни красные. Иногда посередине спины тянется красная полоса, брюхо желтое. Этот вид живет на поросших кустарниками песчаных дюнах. Представители этого рода не переносят высоких температур при большой влажности воздуха, что вредит их очень чувствительной бархатистой коже. Активны днем. В период размножения животные эти очень быстры и подвижны, бегают, плавают, лазают, но не прыгают. В период спаривания оба пола издают крик, состоящий из двух звонких тонов вроде колокольчика и следующей за ними понижающейся трели. Яйца откладываются в мелкие. дождевые лужи, и через 24 часа из них уже выводятся личинки. Пестрый ателоп (A. varius) сверху покрыт черными, желтыми и красными пятнами, брюхо желтое. Этот вид также не прыгает. В неволе самец, спариваясь на протяжении недели, погиб от истощения, так как за то время ничего не ел. Третий вид - A. moreirae достигает 29 мм длины, сверху он темно-коричневый, живот карминно-красный, на горле и груди черные пятна, нижняя поверхность конечностей красная. Он обитает на высоте 1800-2000 м над уровнем моря в богатых водой болотистых местах и, как отметил Дарвин, активен днем, на солнце. У личинки лоскутообразные губы с поперечными складками посредине, окаймленные сосочками. Все три вида - наземные животные и неохотно идут в воду.
К роду Rhinoderma принадлежит единственный представитель - ринодерма Дарвина (R. darwini). Это очень своеобразное животное длиной несколько более 30 мм. Живет она в горах Чили. Ринодерма - настоящий водный обитатель, не переносящий высоких температур. Она охотно сидит в воде, погрузившись в нее по глаза и опираясь на концы пальцев передних лап. При этом длинный мягкий заостренный отросток, имеющийся на конце морды, высовывается вверх. Со звонким писком бросается это пестро окрашенное животное на добычу. Окраска ее чрезвычайно изменчива. Различают три основные вариации окраски верха. Первая характеризуется глинистым цветом спины и резко отличающимися от нее каштаново-бурыми боками и головой. На конечностях поперечные темные полосы. Во втором случае спина окрашена в зеленые тона, в третьем варианте - в серо-бурые, серо-желтые или серо-красные. Две последние расы имеют по основному фону более темный, меняющийся в очертаниях рисунок. Нижняя сторона, имеющая две вариации, разрисована очень изящно и пестро. В первом случае горло и передняя часть груди светлее остальной брюшной стороны и представляют собой все переходы от соломенно-желтого цвета до буровато-оранжевого, причем эти части могут быть покрыты темными пятнами и точками; во втором случае вся нижняя часть окрашена почти одинаково в черно-бурый или оливково-черный цвет. Горло и грудь имеют металлический блеск. На брюхе располагаются большие чисто белые зубчатые пятна, а также желтые и иногда красные. Кроме того, на всех темных частях брюшной стороны имеются мелкие белые, синеватые или кирпично-красные бородавочки. Созревание половых продуктов начинается с конца декабря и продолжается в январе. Брачный период тянется с конца декабря до конца февраля, запаздывая по сравнению с другими земноводными, живущими в этой же местности. Яйца откладываются самкой несколько раз поодиночке или попарно. Самец, после оплодотворения, берет их в рот и запихивает в горловой мешок. Горловой мешок самца сначала так короток и узок, что в него входит не более двух яиц. В стадии полного развития зародышей размеры горлового мешка зависят от количества находящихся в нем детенышей, и он простирается до заднего конца туловища. Присутствие зародышей в горловом мешке не мешает самцу нормально питаться. Яйца у ринодермы Дарвина очень велики и содержат большое количество желтка. В течение первой половины развития до появления передних конечностей детеныши не связаны со стенками мешка и лежат в нем беспорядочно. Когда кончается запас желтка, эмбрионы располагаются двумя слоями так, что их спины обращены к стенкам мешка, а друг к другу они повернуты брюхом. Со стенкой горлового мешка сначала срастается хвостовой плавник, а затем и спинная часть головастика. Дальнейшее питание происходит через стенку этого мешка, богатую кровеносными сосудами. Кожа головастиков лишена самого верхнего слоя эпидермиса, который затруднял бы всасывание. Головастики остаются в мешке до окончания своего превращения, и их хвостовой плавник совершенно исчезает еще до рождения. Детеныши покидают мешок поодиночке, так как развиваются они неодинаково быстро. Вынашивающий потомство самец худеет, а затем восстанавливает силы, видимо, в весенние месяцы, изобилующие насекомыми (сентябрь - декабрь). В связи с этим размножается ринодерма Дарвина позднее, чем другие земноводные, обитающие рядом.
Семейство Настоящие лягушки (Ranidae)
Одно из самых больших семейств отряда бесхвостых земноводных, объединяющее более 400 видов, входящих в 32 рода. Чрезвычайно разнообразные амфибии этого семейства характеризуются присутствием зубов на верхней челюсти, цилиндрическими, нерасширенными (или слабо расширенными) поперечными отростками крестцового позвонка и отсутствием вставочных хрящей между фалангами пальцев. Вероятным центром возникновения амфибий этого семейства следует считать восточное полушарие, причем Африка стала местом их наибольшей дифференциации. Ныне распространены по всему свету, за исключением арктических районов, Австралии и крайнего юга Южной Америки.
Самый обширный род - настоящих лягушек (Rana) объединяет более 200 видов. Сюда относятся как очень мелкие виды с максимальной длиной тела до 30 мм, так и самая крупная из бесхвостых земноводных - лягушка-голиаф, достигающая 326 мм.
Озерная лягушка (Rana ridibunda) - самый крупный вид среди земноводных нашей фауны. Наибольший ее размер - 170 мм. Самки всегда крупнее самцов. Однако в разных местах обитания размеры животных заметно изменяются. Максимальных размеров достигают озерные лягушки, обитающие между 45-50° с. ш. и 30-50° в. д. Иными словами, самые крупные особи живут в центре ареала, отличающемся, видимо, самыми благоприятными условиями существования для вида. По мере продвижения к границам ареала размеры озерной лягушки уменьшаются. Так, в дельте Волги самые большие самки достигают 149 мм, а самцы 128 мм. Севернее, в Воронежской области, самые большие самки имеют длину тела 117 мм, а самцы 112 мм. В Туркмении, территория которой лежит в зоне пустынь, по южной границе распространения вида, наибольшая из пойманных озерных лягушек достигала 88 мм. Размеры животных изменяются не только в разных частях ареала, но и в разных местах обитания, незначительно удаленных друг от друга. Например, озерные лягушки, населяющие окрестности Астрахани, оказались крупнее лягушек того же возраста, живущих от них примерно на расстоянии 80 км - в нижней зоне дельты Волги. Разница в длине тела у молодых самок составила 20-25 мм, а у самцов 30 мм. По-видимому, меньшие по размерам лягушки находились в худших условиях питания.
Неподвижную озерную лягушку нелегко заметить среди водной или прибрежной растительности, благодаря тому что она окрашена сверху в зеленый, оливковый или темно-коричневый цвет с большим или меньшим количеством черных или темно-зеленых пятен. Иногда вдоль спины у нее тянется светлая полоса. Снизу она грязно-белого или желтоватого цвета, обычно с темными пятнами. У самцов в брачное время на первом пальце передней ноги развиваются утолщения серого цвета - брачные мозоли. У квакающих самцов по углам рта бывают видны серые резонаторы.
Озерная лягушка распространена по всей Европе и в пределах нашей страны, проникая в Азию, доходит на восток до озера Балхаш. Северная граница ее распространения почти совпадает с южной границей тайги. У нас она обитает в Казахстане, Средней Азии, на Кавказе, в Крыму; за пределами нашей страны этот вид встречается в Иране, Малой Азии, Иордании, ОАР и Алжире, находя здесь южную границу распространения. Озерная лягушка характерна как для широколиственных лесов, так и для степей. На юге она проникает и в зону пустынь, а на севере краем ареала заходит в тайгу. Поднимается в горы до 2500 м.
Всю жизнь эта лягушка проводит в воде или неподалеку от нее, населяя самые разнообразные типы водоемов, в том числе и большие, глубокие, быстротекущие реки. При большой влажности воздуха и высоких температурах, например в Южном Дагестане, она охотится дальше от воды, чем в средней полосе. В окрестностях Еревана озерные лягушки отходят от водоема на 2-3 м, иногда до 15-20 м, а молодые экземпляры - на 4-5 м.
Тесная связь с водоемами позволяет озерной лягушке осваивать такие малодоступные для земноводных ландшафты, как пустыни.
Озерная лягушка относится к многочисленным видам. В дельте Волги, в некоторых ильменях, используемых для рыборазведения, живет до 60 тыс. озерных лягушек. В Колхидской, Алазано-Авторанской и Ленкоранской низменностях численность этих животных достигает нескольких десятков на 100 кв. м. В Туркмении на километровом маршруте по берегу реки Карасу (район Багира) насчитывали до 141 особи этого вида. Средняя плотность населения лягушек в окрестностях Алма-Аты от 1000 до 2000, а в окрестностях Илийска от 450 до 1000 особей на 1 га. Однако получение точных данных о численности озерной лягушки в разных частях ее ареала - это задача будущих исследований.
Особенности суточной активности озерной лягушки подробно наблюдались в Южном Дагестане летом в мелководной старице реки Самура, неподалеку от места впадения этой реки в море. В любое время суток общее число озерных лягушек, плавающих на поверхности воды и прыгающих в зарослях прибрежной растительности по берегам старицы, остается примерно одинаковым. Однако дважды в сутки они совершают перемещение на сушу и обратно. На берегу их много с 21 до 7 часов и с 11 до 17 часов. Самое большее число лягушек на суше наблюдается в час ночи и в час дня. Количество лягушек в воде уменьшается соответственно тому, как оно возрастает на суше. Во время пребывания лягушек на берегу желудки их наполнены максимально. Прибрежные заросли оказываются для них главным местом охоты. В воде животные спокойно лежат на поверхности или лениво передвигаются. В это время происходит переваривание пищи и опустошение желудка. Водоем - это место отдыха с наиболее благоприятными условиями температуры и влажности, дающее в то же время и надежное укрытие от врагов. Появляясь на суше и ночью и днем, озерные лягушки оказываются животными с круглосуточной активностью. Во время дневной активности лягушки все время ненадолго уходят в водоем пополнить запас влаги в теле, благодаря этому днем какое-то количество лягушек встречается не только на суше, но и в воде. Ночью же в часы наибольшей активности все лягушки на суше и не идут в водоем, так как при более низких температурах они не подвергаются опасности высыхания.
Суточный ритм поведения озерных лягушек не одинаков в разных частях их ареала. Так, в Туркмении летом на берегах водоема озерные лягушки встречаются чаще всего в ранние утренние часы, по вечерам и ночью. В жаркие дневные часы основная масса животных находится в воде. Перестают охотиться и те, которые находятся на суше, оставаясь в тени и на увлажненных участках среди прибрежной растительности. Желудки у большинства особей в это время не содержат пищи. В начале марта, когда по утрам еще свежо, лягушки вылезают на берег обычно не раньше 9 часов, и уже к 10 часам количество греющихся на солнце особей заметно возрастает. Между 10 и 16 часами животные интенсивно кормятся, и их в это время раза в два-три больше на суше, чем в водоеме. К вечеру, наоборот, количество лягушек больше в воде, чем на берегу. Однако уже во второй половине марта ночи бывают теплыми и лягушки становятся активными на протяжении всех суток.
Следовательно, характер суточной активности меняется и по сезонам. Эти изменения касаются не только времени, когда животные особенно энергично охотятся, но и интенсивности охоты. В дельте Волги неполовозрелые озерные лягушки в апреле мало кормятся, и их желудки очень незначительно наполнены. Постепенно они начинают все чаще и чаще кормиться, и до начала августа интенсивность питания непрерывно растет, а затем резко идет на убыль. Та же картина наблюдается и у самцов. Они отличается от молоди лишь тем, что до конца мая активность, связанная с кормежкой, у них возрастает очень незначительно. В это время деятельность, связанная с процессами размножения, преобладает у самцов над всеми другими. Если они и не соблюдают в полной мере так называемого брачного поста, то питаются значительно меньше, чем в другое время года. Своеобразна активность самок. Весной они начинают кормиться позже молодых и самцов, но наибольшая степень наполнения желудков у них отмечена во второй половине мая. С этого времени активность их начинает снижаться и к концу августа мало отличается от активности самцов. В среднем самой большой активностью добывания корма в течение лета отличаются молодые, неполовозрелые лягушки, лишь немногим, примерно на 1/5, меньше она у самок, у самцов же активность питания почти вдвое меньше, чем у самок.
По мере понижения температуры окружающей среды снижается активность озерных лягушек, и они уходят в зимнюю спячку. В южной части Армении спячка начинается при средней температуре воздуха 11,5° и средней температуре воды 8°. Зимуют озерные лягушки на дне водоемов, мигрируя осенью либо в более глубокие из них, либо к родникам. При осенних перемещениях на места зимовок лягушки могут проходить значительные расстояния. Зимующие лягушки часто скапливаются под нависающими берегами или скрываются в подводной растительности. В разных климатических зонах озерные лягушки уходят на зимовку неодновременно. В горах спячка начинается раньше, чем на равнине. Так, в Южной Армении озерные лягушки уходят на зимовку во второй половине октября, а в окрестностях Махачкалы задерживаются до середины ноября. Также раньше впадают в спячку и популяции, обитающие севернее. Под Курском озерные лягушки перестают встречаться на суше в сентябре - октябре. В Туркмении резкое сокращение их активности замечается к концу ноября. Однако здесь о настоящей спячке у озерной лягушки говорить трудно. Часть их остается активной. В незамерзающем родниковом истоке в Багире бодрствующие лягушки не были редкостью даже при отрицательной температуре воздуха (-4°) в течение всего года. Большинство впадает в неглубокий сон; они хотя и вялы, но не лишены способности плавать и прыгать. Потревоженные животные без особого труда перемещаются и укрываются в другом месте. Возле артезианских колодцев и родников озерные лягушки не впадают в спячку и в Южной Армении.
Различно и время выхода озерных лягушек с зимовки. В Туркмении это конец февраля - начало марта. Также в начале марта просыпаются озерные лягушки под Одессой и в окрестностях Махачкалы, а во второй половине марта - под Ереваном. В это время средняя температура воздуха около 10°. Под Курском этот вид появляется в апреле, под Москвой - в мае. На сроки пробуждения лягушек значительно влияет высота над уровнем моря. Так, в Боржомо-Бакурианском районе, на высоте 1143 м над уровнем моря, они просыпаются в первых числах мая, а на высоте 1655 м над уровнем моря - в первых числах июня. Молодые уходят на зимовку позднее. Под Ереваном они задерживаются до конца ноября, тогда как основная масса взрослых впадает в спячку до первой половины ноября. Весной они просыпаются от зимнего сна несколько раньше взрослых. В результате продолжительность зимовки в низменных районах Кавказа равна 60-90 дням, в Туркмении - 90-95, под Киевом - 150180, под Москвой - 210-230.
От времени первого появления лягушек до начала икрометания проходит от одной недели до месяца. У южных популяций этот интервал, по-видимому, меньше, чем у северных. В период размножения самцы держатся на поверхности воды, образуя большие скопления. Они очень подвижны и громкоголосы. Их "брачные песни" привлекают самок. Откладке икры предшествует спаривание. Манера обхвата самки при этом у всех лягушек, в том числе и у озерной, своеобразна. Самец схватывает ее позади передних лап так, что его лапы сходятся на груди у самки. Это своеобразное спаривание имеет большое биологическое значение. Оно стимулирует одновременное выметывание в воду икры и сперматозоидов, увеличивая процент оплодотворенных яиц при наружном оплодотворении. Икра откладывается в виде комка, образовавшегося благодаря склеиванию слизистых яйцевых оболочек.
Диаметр яйца озерной лягушки - 1,5-2,0 мм, а всей икринки - 7-8 мм. Верхняя половина яйца темно-бурая, а нижняя белая.
Количество яиц, откладываемое одной самкой, до определенных размеров возрастает с увеличением длины ее тела. Так, в дельте Волги лягушки длиной 91-95 мм откладывают в среднем 3916-3989 яиц, у лягушек размером 106-109 мм число яиц возрастает до 4540-5195.
Среднее число яиц при длине тела самки 110-115 мм достигает 5408-6818 штук, при 116-119 мм - 7969-9360 штук. У лягушек величиной 120-126 ми в некоторые годы плодовитость резко снижается, достигая уровня, характерного для молодых самок (3614 яиц), только что вступающих в размножение. В другие годы плодовитость этой размерной группы продолжает расти (11 237 яиц), но тогда она снижается у более крупных самок, достигающих 128 мм. Число икринок у такой самки равнялось 2935 штук. Как показывают приведенные цифры, плодовитость зависит не только от величины животного, так как число яиц, откладываемых самками одинаковой величины, изменяется по годам. Видимо, в этих случаях сказываются условия жизни вида, не остающиеся постоянными из года в год. Плодовитость озерных лягушек из других частей ареала не выходит за пределы, полученные для нее в дельте Волги.
Озерные лягушки выметывают икру одним комком или отдельными кучками от 3 до 10. Период икрометания растянут. Особенно продолжителен он у южных популяций. Эта растянутость может определяться откладкой икры порциями или неодновременным созреванием ее у разных особей. В Туркмении, возможно, имеют место две кладки в году.
Икрометание начинается тогда, когда средняя температура воды достигает 15,6-18,6°. Это свидетельствует о значительной теплолюбивости озерной лягушки. В соответствии с теплолюбивостью этих животных и сперматозоиды их обладают высокой теплоустойчивостью. Они без вреда могут выносить нагревание до 41,4°. Теплоустойчивость сперматозоидов не меняется у лягушек, обитающих в разных участках ареала.
Скорость развития икры находится в теснейшей зависимости от температуры окружающей среды. В Армении в июне при средней температуре воды 20,4° и воздуха 21,9° развитие яиц длится 7-8 дней. В мае при средней температуре воды 16,4° и воздуха 11,2° развитие продолжается 9-10 дней. В среднем по этим двум наблюдениям для развития яиц озерной лягушки требуется 154,4 градусодня. На юго-востоке Казахстана нормальное развитие икры происходит при температуре 18-24°. В одной и той же местности в водоемах, по-разному прогреваемых, срок развития икры не одинаков.
Длина только что появившегося из яйца головастика озерной лягушки в Армении - 7-8 мм, в "Туркмении - 4,8-5 мм. У них уже довольно длинный хвост окружен хорошо развитым плавником. Наружные жабры разделены на ряд лопастей. Эти особенности строения показывают, что головастики озерной лягушки покидают яйцо на более поздних стадиях развития, чем у некоторых других бесхвостых земноводных, например у остромордой лягушки. В это время окраска тела головастиков светло-желтая или коричневая. Достигая примерно 30 мм длины, головастики заметно зеленеют.
Личинки озерных лягушек первое время остаются на тех местах, где они появились на свет, и держатся кучкой, но затем очень скоро расплываются по всему водоему. Их можно встретить в толще воды как на мелких местах, так и на более глубоких, как в зарослях растений, так и в чистой воде. В глубоких и больших водоемах головастики обычно держатся у берегов, где вода теплее и где им, вероятно, легче добывать корм. Они ведут дневной образ жизни и наиболее интенсивно питаются в 10-12 часов. На ночь головастики опускаются на дно и прячутся под камнями и растительностью.
Прорыв рта и переход к активному питанию в Туркмении происходит тогда, когда личинка достигнет 16 мм длины. В дельте Волги головастики к этому времени имеют длину 16,8 мм. В разных водоемах их размеры оказываются неодинаковыми. На полоях они достигают 22,2 мм, в ильменях - 16,7 мм, а в авандельте - 11,3 мм.
Питание головастиков озерной лягушки изучено с применением точных методов анализа содержимого желудков так, как ни у одного другого вида наших земноводных. Они питаются не высшими растениями, как это было принято считать, а главным образом водорослями. Преобладающую группу их кормов составляют диатомовые и зеленые водоросли, в большинстве представляющие собой мельчайшие растительные организмы, часто одноклеточные. Самые крупные из них - нитчатые зеленые водоросли, очень тонкие и нежные, но имеют значительную протяженность. Диатомовые и зеленые водоросли были встречены у головастиков во всех исследованных желудках, причем составляли примерно 60% веса этого содержимого. Ко второстепенным кормам относятся простейшие, коловратки, сине-зеленые водоросли и жгутиковые. Случайные корма включают плодовые тела низших грибов (плесени), эпидермис (кожицу) высших растений, мельчайших представителей круглых и кольчатых червей, моллюсков, ракообразных, мшанок и насекомых. Все они не играют сколько-нибудь заметной роли в рационе головастиков.
Организмы, идущие в корм головастикам, относятся к группе обрастателей, живущих на подводных растениях, или ведут придонный образ жизни на мелководье. Обитателей толщи воды поедают головастики мало. Вероятнее всего, они попадают к ним в пищу тогда, когда, отмирая, падают на дно или оседают на подводных растениях. Своеобразное строение ротового аппарата головастиков отлично приспособлено к соскребанию пищи с растений или со дна. Их маленький рот окружен выдающимися вперед бахромчатыми губами, которые образуют небольшой конический хоботок. Верхняя губа меньше и менее подвижна, чем нижняя. Нижняя длиннее и шире, значительно мягче и подвижнее. По свободному краю ее идут в несколько рядов небольшие мясистые сосочки, имеющие, по-видимому, осязательную функцию. Они скопляются преимущественно в углах рта. Ротовое отверстие ограничено двумя крепкими роговыми "челюстями", напоминающими клюв. Внутренняя поверхность обеих губ между свободным краем их и клювом образует поперечные складки, на гребнях которых, так же как и по свободному краю губ, появляются небольшие черные роговые зубчики. Каждый из зубчиков головастика - это одна видоизмененная эпителиальная клетка. Он быстро изнашивается, и на смену ему немедленно возникает точно такой же.
Вес поедаемой головастиками пищи растет с увеличением веса их тела, но не пропорционально. В то время как вес тела увеличивается в 40 раз, количество потребляемой пищи возрастает только в 15 раз. Происходит это потому, что с возрастом прожорливость этих животных уменьшается. В дельте Волги у головастиков длиной в 17 мм вес пищи составляет в среднем 5,9% от веса их тела, при длине в 35 мм -5,4%, а при длине 63 мм - 2,3%, т. е. потребление пищи в процессе развития уменьшается примерно в 3 раза.
Личиночный период развития у озерной лягушки один из самых длинных среди наших бесхвостых земноводных. Несмотря на то что наружные жабры исчезают у них раньше, чем у других лягушек, на 7 дней, почки задних конечностей появляются поздно - на 32-й день. Задние конечности разделяются на отделы к 59-му дню и приобретают подвижность к 74-му дню. Передние конечности появляются на 82-й день, и на 84-й день начинается рассасывание хвоста. В общем личиночный период развития занимает 80-90 дней, а может быть и значительно большим.
Зато растут головастики озерной лягушки быстрее, чем у многих других видов. Их средний прирост за день с момента вылупления до метаморфоза в искусственных условиях составляет 1,0 мм. Перед метаморфозом длина головастика в средней полосе составляет 70-90 мм, в дельте Волги - 55-69 мм, в Армении и Туркмении - 50-52 мм. Они лишь на 15-25% меньше молодых половозрелых лягушек. Во время интенсивного формирования органов темп роста головастиков замедляется, составляя 0,5 мм за день (прибавление веса не больше 4,9 мг). В период интенсивного роста увеличение длины головастика за день составляет 0,7-1,5 мм (прирост веса 7,6-11,2 мг). В глубоких водоемах рост происходит несколько медленнее, чем в мелких.
Существует ряд данных, говорящих о зависимости сроков развития личинок от температуры. Так, у озерной лягушки в Московской области личиночный период длится 80-85 дней, в районе Киева - 70-75 дней, на Кавказе (низменность) - 55-60 дней. В холодных горных водоемах головастики не успевают метаморфизировать и зимуют на этой стадии развития. Иногда в глубоких водоемах это наблюдается и под Москвой - у северной границы распространения вида.
Наилучшая температура воды для существования головастиков озерных лягушек 18-28°. Максимальная температура воды, при которой они могут существовать, 43°. При температуре воды 5-6° развитие головастиков прекращается, а при 1-2° они гибнут.
Скорость наступления метаморфоза связана также с характером питания личинок. В условиях опыта удалось задержать наступление метаморфоза у животноядных головастиков, выкармливая их водорослями. Вероятно, в связи с растительноядностью головастиков озерной лягушки им свойствен длительный период развития.
Имеет значение и количество поедаемой пищи, определяющееся, видимо, не только потребностями организма, но и кормностью водоемов. Так, головастики, развивающиеся в авандельте Волги, к периоду метаморфоза оказываются крупнее развивающихся в ильменях и полоях. При этом на протяжении всего времени после прорыва рта и перехода к активному способу питания они, в расчете на грамм веса тела, поедают пищи больше, чем головастики в других местах обитания.
С первых дней своего существования головастики находятся в состоянии метаморфоза, с каждым днем приобретая все новые и новые особенности, характерные для взрослого животного, ведущего наземный образ жизни, причем на каждом предыдущем этапе развиваются системы органов, которые начинают функционировать на последующем. Однако метаморфозом обычно именуют те изменения, которые происходят в непосредственной связи с переменой среды обитания и приводят к полной потере личиночных органов. Метаморфоз у головастиков озерной лягушки, как и у всех других, начинается с изменений кишечника, так как организм перестает питаться, затем освобождаются, прорывая жаберные крышки, передние конечности. Далее порядок изменений в строении головастиков у лягушек, подобно озерной всю жизнь живущих в водоеме, и у тех, которые приходят в него только на период размножения, не одинаков. Различия связаны, видимо, с тем, что у первых при превращении личинок во взрослую форму смена среды обитания выражена очень незначительно, так как сеголетки остаются в водоеме и выходят на сушу только кормиться. У личинок озерной лягушки сначала начинает исчезать хвост, изменяется строение глаз, перестраивается ротовой аппарат и только после этого, когда личинка приобретет вид взрослого животного, исчезают органы водного дыхания - жабры. У личинок наземных бесхвостых земноводных исчезновение жабр происходит гораздо раньше, вслед за появлением передних конечностей. Наконец, изменяется строение кожи, и прежний головастик становится лягушкой, отличающейся от взрослых только размерами и недоразвитием половых органов. Метаморфоз у озерной лягушки продолжается около 5 дней.
Только что метаморфизировавшие сеголетки по размерам обычно значительно меньше головастиков. В Армении они имеют длину 14-15 мм. В дельте Волги средний их размер сразу после метаморфоза в июле - 26 мм. На зимовку они уходят, достигнув 30-39 мм длины, а отдельные экземпляры и 55 мм. В Воронежской области средние размеры сеголеток, уходящих на зимовку,- 20-30 мм, а наиболее крупных - 32-34 мм. Во время спячки лягушки почти совсем не растут.
На следующий год в конце мая перезимовавшие сеголетки в дельте Волги имеют длину тела 40-49 мм, в конце июня - 50-59 мм, а в конце июля - 70-79 мм. До конца августа самки и часть самцов остаются такого же размера. Некоторые самцы дорастают до 80-89 мм. В течение следующего лета двухгодовалые самцы достигают длины более 90 мм, а самки - 90-99 мм. К 5 годам самки имеют 130-139 мм длины. С возрастом рост замедляется, хотя и не прекращается полностью всю жизнь.
Половозрелости озерные лягушки достигают на третьем году жизни, когда длина тела самцов равна 80-89 мм, а самок - 90-99 мм. В Воронежской области половозрелость наступает у лягушек длиной в 70-80 мм, а под Казанью - 60-70 мм. Продолжительность жизни озерной лягушки в природе 6-7 лет. Особенно велика смертность сеголеток. Летом после метаморфоза они представляют основную часть популяции. В Туркмении в водоемах у Багира подсчитано, что в июне сеголетки составляли 62,5% от всей популяции. Однако в марте следующего года их доля равнялась всего лишь 14,5%.
Численность озерных лягушек в одних и тех же местах в разные годы может изменяться, однако вопрос этот очень мало изучен. Особо важную роль в их жизни должно играть пересыхание водоемов во время засухи, что мешает нормальному росту и развитию головастиков и лягушат. Однако для тех озерных лягушек, которые обитают в достаточно глубоких, постоянных водоемах, этот фактор имеет очень малое значение. В некоторых случаях сеголетки, которые уходят на зимовку позднее взрослых, в массе гибнут, застигнутые неожиданными морозами. Возможна гибель и на зимовках от недостатка кислорода в водоеме.
В особенностях питания озерной лягушки проявляются все характерные черты, свойственные лягушкам вообще. Это животноядные организмы. Список поедаемых ими кормов очень велик, большинство из них - беспозвоночные, главным образом насекомые. Среди них первое место почти везде, где бы ни изучалось питание этого вида, занимают жуки. Только в окрестностях города Либехов (Чехословакия) на первом месте оказались перепончатокрылые. На втором месте здесь, так же как и под Казанью, стоят двукрылые. Возле Махачкалы и в Туркмении второе место занимают перепончатокрылые, а в Армении - прямокрылые. Как бы то ни было, но преобладающими кормами озерной лягушки везде оказываются наиболее массовые животные.
Существуя главным образом за счет насекомых, озерная лягушка, в отличие от других бесхвостых нашей фауны, нападает и на позвоночных. В некоторых случаях добычей этого вида становятся мелкие млекопитающие, например землеройки или молодые полевки. Многочисленны указания на схватывание лягушкой, сидящей у воды, мелких птиц; описано нападение ее на птенцов поганки, гнездящейся на воде. Однажды была найдена мертвая лягушка с торчащим изо рта пуховым птенцом чибиса. Встречаются в желудках озерной лягушки и ужата, и некрупные земноводные (квакши, остромордые лягушки), и головастики, и лягушата, в том числе и свои собственные. Однако млекопитающие, птицы и пресмыкающиеся - редкие объекты питания озерной лягушки. Головастики, лягушата и мальки рыб - наоборот. В некоторых случаях они могут составлять определенную долю в питании этого земноводного. Так, собственные головастики становятся основным кормом во время сильного паводка в дельте Волги, когда другие корма сносятся водой или становятся недоступными. Значительное число мальков может быть уничтожено озерной лягушкой в рыбхозах и на рисовых полях, где разводят рыб. Одним словом, при больших, искусственно создаваемых концентрациях молоди. Однако, по наблюдениям в Армении, при наличии массы мальков рыб и головастиков озерные лягушки продолжали все-таки поедать главным образом насекомых.
Несмотря на то что озерная лягушка всю жизнь теснейшим образом связана с водоемами, значение наземных организмов в ее питании гораздо больше, чем водных. В средней полосе наземные корма составляют 68% от числа всех кормов, встречающихся в желудках, в Предкавказье - 86%, в окрестностях Махачкалы - 73-95% и в Туркмении - 95%. Это свидетельствует о том, что озерная лягушка охотится главным образом на суше. По мере продвижения на юг в более оптимальные условия температуры роль наземных форм в питании увеличивается. Это явление хорошо согласуется с тем, что при высоких температурах озерная лягушка больше времени проводит на суше, а при большой влажности окружающей среды и дальше отходит от водоемов.
В питании озерной лягушки велик также процент летающих форм (24%). В этом отношении среди земноводных средней полосы она уступает только прудовой лягушке (27%). В Туркмении значение летающих животных в питании озерной лягушки резко повышается. Они составляют более 60% от числа всех встреченных форм. Возможность схватить летающее животное связана у лягушек со способностью совершать большие прыжки, а также и со своеобразной манерой охоты. Они могут молниеносно выбрасывать далеко вперед вытягивающийся клейкий язык, прикрепляющийся во рту не своим основанием, а лишь передним концом. Прилипшая к языку добыча подтягивается ко рту и схватывается челюстями, снабженными мелкими, заметными только на ощупь зубами. У озерных лягушек доля летающих кормов увеличивается также и благодаря их дневной активности, так как активность летающих форм днем тоже наибольшая.
Набор кормов, употребляемых озерными лягушками, несколько меняется не только в разных географических пунктах, но часто и в близлежащих местах. Например, в окрестностях Махачкалы у лягушек из водоема, лежащего в полупустынном ландшафте, водные животные составляли 27%. Преобладающими кормами здесь были насекомые, встречающиеся в 78% вскрытых желудков (из них 67% представляли жуки, 39% - двукрылые) и 33%-позвоночные. В другом водоеме на склоне горы водные животные в питании лягушек встречались редко (5%). Во всех вскрытых желудках были найдены насекомые, из них жуки, двукрылые и перепончатокрылые были обнаружены в 60% желудков. Позвоночными эти лягушки не питались. Интересно, что и средняя степень наполнения желудков, вычисленная в процентах от веса тела, у лягушек из первого водоема оказалась примерно в два раза большей, чем во втором водоеме. Наблюдения проводились между 12 и 13 часами в одном водоеме 25, а в другом 27 мая.
Разнообразие состава кормов может значительно изменяться на одном и том же месте в разные годы. Список кормов озерной лягушки в дельте Волги в 1956 г. включал 67 животных, в 1957 - 36, в 1958 - 44, а в 1959 - 21.
Меняются особенности питания и в разные месяцы. Например, в дельте Волги в начале мая и в конце августа насекомые встречаются в 30% желудков, а в остальное время - в 70-74%, рыбы в начале мая найдены в 14% желудков, а в другие месяцы - в 1-3%. Встречаемость земноводных в желудках озерной лягушки особенно велика в июне - начале июля (28%), тогда как в другие сроки они обнаружены в 16-20% желудков. В течение лета происходит также увеличение в питании доли водных обитателей.
Различные возрастные группы озерной лягушки различаются между собой и по размерам животных, употребляемых в пищу. У всех у них господствующими кормами будут жуки, но молодые лягушки, и особенно сеголетки, питаются более мелкими формами. Сеголетки в больших количествах поедают цикадок, имеющих в длину 3-4 мм. В пище более старших лягушек эти животные отсутствуют. В определенных условиях озерная лягушка в больших количествах уничтожает медведок, причем неполовозрелые лягушки и сеголетки употребляют в пищу главным образом личинок этого насекомого. В питании молодых лягушек, больше чем у взрослых, встречаются муравьи и пауки.
Сеголетки кормятся почти исключительно на суше. Водные организмы составляют в их пище всего 6% от общего числа встреченных кормов. У неполовозрелых лягушек более старших возрастов они составляют 26%, а у взрослых 38%.
У сеголеток в среднем разнообразие кормов наименьшее - 30 форм, против 34-55 у старших возрастов. Доля преобладающих кормов у них несколько выше (90%), чем у лягушек других возрастов (82-88%). Причины этих возрастных особенностей питания, видимо, в том, что молодь ограничивается лишь более мелкими формами, тогда как старшие могут употреблять в пищу и мелких и крупных животных. Кроме того, неполовозрелые лягушки охотятся главным образом на суше и только увеличиваясь в размерах начинают в больших количествах добывать пищу и в воде. Возможно, молодые отличаются большей избирательной способностью в отношении корма и меньшей ловкостью на охоте. Однако эти предположения еще не имеют подтверждений.
Среди всех наших земноводных озерные лягушки привлекают наибольшее внимание с точки зрения оценки их значения в производственной деятельности человека. Это объясняется тем, что они наносят вред, поедая мальков рыб. Необходимо оценить размеры наносимого ими вреда. Выяснилось, что озерные лягушки в естественных условиях поедают очень незначительное количество рыбы. Их склонность к этому корму значительно возрастает там, где плотность населения мальков оказывается повышенной. Это происходит в водоемах искусственного рыборазведения и на рисовых полях, где выращивают мальков рыб, причем и здесь сколько-нибудь значительное количество мальков поедается только в определенных местах их концентрации, например у шлюзов. Следовательно, воздействие озерных лягушек на продуктивность рыбхозов очень незначительно.
Предполагалось, что головастики озерной лягушки могут конкурировать из-за пищи с молодью рыб. Но изучение этого вопроса показало, что эти предположения не имеют никаких оснований.
Головастики озерной лягушки, образующие громадные скопления, играют заметную роль в круговороте веществ в природе. Представление о численности головастиков в отдельных водоемах дают следующие числа: в ильменях дельты Волги на 1 м3 водоема приходится в среднем 9000 головастиков. Средняя за сезон биомасса головастиков в этих водоемах равна 400 г/м3. Биомасса головастиков в одном ильмене может достигать веса в 11,5 т, а во всех ильменях Дамчикского участка Астраханского заповедника достигает приблизительно 2282,5 т.
Вся эта масса головастиков живет за счет диатомовых и зеленых водорослей, малодоступных другим позвоночным животным. Головастики, в свою очередь, поедаются некоторыми хищными рыбами, а из наземных обитателей ужами и разнообразными птицами: цаплями, чайками, крачками, утками, некоторыми куликами, зимородками и даже такими птицами, которые не связаны в питании с водоемами. Например, головастиков озерной лягушки охотно едят сизоворонки, сороки, дрозды.
Головастики озерной лягушки могут играть существенную роль в выкармливании домашней птицы.
Многие животные поедают и взрослых лягушек. К ним, например, относятся сомы, судаки, щуки, османы, ужи, аисты, цапли, чайки, крачки, поганки, пастушки, коршуны, болотные луни, змееяды, сарычи, курганники, домовые сычи, филины, вороны, грачи, сизоворонки, сорокопуты, жуланы, лисицы, шакалы, барсуки, выдры и даже домашние кошки.
Озерные лягушки, питающиеся главным образом наземными кормами и в свою очередь поедающиеся рыбами, повышают кормность водоемов именно за счет этих наземных беспозвоночных, играя роль промежуточного звена. Становясь пищей пушных зверей и промысловых рыб, озерная лягушка оказывается полезным животным и с точки зрения хозяйственной деятельности человека.
Если же к этому прибавить и уничтожение озерными лягушками вредных насекомых, то этот вид в целом окажется гораздо более полезным, чем вредным, для человека.
Прудовую лягушку (Rana esculenta) от озерной хорошо отличает высокий внутренний пяточный бугор, более или менее сжатый с боков. Обычно она ярко-зеленая со светлой полосой вдоль спины и с большим или меньшим количеством черных пятен. Встречаемость продольной спинной полосы увеличивается по направлению на север и на восток. В отличие от озерных лягушек среди прудовых иногда попадаются особи с темным височным пятном (9%). Снизу прудовая лягушка белого или желтоватого цвета с темными пятнами или без них.
У самцов в брачное время на первом пальце передней ноги темный бугор - брачные мозоли; в углах рта наружные белые или желтоватые резонаторы. Весной плавательные перепонки на задних ногах у самцов разрастаются значительно меньше (на 35%), чем у бурых лягушек, а у самок несколько больше - на 13% вместо 2-8%.
Прудовая, или, как ее часто называют, съедобная, лягушка значительно уступает по размерам озерной. Ее максимальная длина 100 мм. По направлению к северу и на восток размеры прудовой лягушки уменьшаются.
Населяет Европу, за исключением Пиренейского полуострова, Южной Франции, Греции и Балканского полуострова. В пределах нашей страны имеет ареал в виде клина, суживающегося к востоку и едва переходящего Волгу в среднем ее течении.
Обитает в водоемах главным образом широколиственных и смешанных лесов. В некоторых местах, например в Беловежской пуще, встречается по влажным лесам и вдали от воды. В степях живет только по водоемам среди речных урем. В тайгу почти не проникает, населяя в ее южных районах только водоемы открытых ландшафтов. В горы поднимается до 1100 м.
Ночью в средней полосе вне периода размножения лишь отдельные особи прудовой лягушки изредка появляются на поверхности воды. Основная масса животных находится на дне водоема, где температурные условия в это время наиболее благоприятны. Они всплывают в массе на поверхность к 8 часам утра и исчезают к 10 часам вечера. Наибольшее число особей активно между 12 и 16 часами - в самое теплое время суток. В эти часы, как показали наблюдения, лягушки большую часть времени кормятся. В 6-8 часов вес содержимого желудка не превышает 1,1% веса тела; максимум приходится на 12-16 часов, когда содержимое желудка составляет 14% веса тела. Начиная с 20 часов вес съеденной пищи резко падает и к 22 часам не превышает 2% веса тела. Активность прудовой лягушки, постоянно находящейся в условиях оптимальной влажности, зависит от температуры окружающей среды, и в теплые ночи она может и не прекращаться.
Весной прудовая лягушка многочисленна на поверхности водоемов с 10 до 22 часов. Кривая активности этого вида имеет двувершинный характер. Первый пик приходится на 14-16 часов, второй - на 20-22 часа. В течение почти всего активного времени большинство лягушек находится в воде, и только в самое теплое время суток они перекочевывают на берег или плавающие в воде предметы. Здесь они заняты охотой, о чем свидетельствует наполнение желудков, составляющее почти 20% от веса тела. Этот первый пик активности связан с питанием. Во время второго пика активности, в 20-22 часа, наблюдается наибольшее число спаривающихся и поющих особей, а вес корма в желудке не превышает 4% от веса тела. Следовательно, оживление лягушек в это время связано с процессами размножения.
Разница между весенней и летней активностью прудовой лягушки заключается в том, что летом продолжительность кормежки больше, чем весной. Этому благоприятствует более высокая температура воздуха и воды.
В активный период большую часть пищи прудовая лягушка добывает на суше. Водные корма имеют в ее рационе меньшее значение, чем у озерной, но во много раз большее, чем у бурых лягушек. Кроме жуков и двукрылых, в питании. этих земноводных играют заметную роль стрекозы и муравьи. Эти преобладающие корма составляют 66% от всех встречающихся. Примерно у 9% молодых прудовых лягушек встречаются комары, в уничтожении которых этот вид имеет большее значение, чем другие лягушки. Корма, общие с озерной лягушкой, составляют 43% от всех животных, встречающихся в желудках прудовой. Интересно, что у озерной лягушки они составляют 69%. Эта разница, видимо, объясняется не только большей или меньшей приверженностью видов к водоемам, но и разными типами водоемов, в которых обитают прудовые и озерные лягушки. Например, веснянки и поденки, входящие в состав кормов озерной лягушки, отсутствуют у прудовой. Это, должно быть, связано с тем, что указанные насекомые откладывают яйца в быстро текущие водоемы, занимаемые озерной лягушкой, но избегаемые прудовой. Имеют также значение и размеры добычи. Более крупные лягушки поедают и более крупных животных. Прудовая лягушка среди других наших земноводных добывает максимальное количество летающих насекомых; более 26% животных, найденных в желудках этого вида, принадлежит к ним.
Зимняя спячка у прудовой лягушки в среднем 100 дней, на 15-25 дней больше, чем у бурой, но несколько короче, чем у озерной. Это наиболее теплолюбивый вид среди наших лягушек.
Проснувшись, прудовые лягушки, как и все зеленые лягушки, не сразу приступают к размножению. Обычно они мечут икру во второй половине мая, позднее озерных, спустя 15-20 дней после пробуждения. Одна самка откладывает 2000-3000 икринок диаметром 1,5-2 мм. Размножение растянуто, так как икра откладывается несколькими порциями. Температура воды, в которой развивается икра прудовой лягушки, как правило, не падает ниже 16° и не поднимается выше 31°. Развитие ее идет значительно быстрее, чем у бурых лягушек, нерестящихся ранней весной. Однако скорость развития яиц в эксперименте при одних и тех же условиях у травяной лягушки несколько большая, чем у прудовой. Яйца прудовой лягушки отличаются большей устойчивостью к высоким температурам, чем яйца травяной. Личинки выклевываются из яйца на сравнительно поздних стадиях развития, хвост у них окружен хорошо развитым плавником и имеет вытянутую форму, наружные жабры разделены на ряд лопастей. На 6-й день, раньше, чем у всех других видов лягушек, головастики прудовой теряют наружные жабры. На 30-й день у них появляются зачатки конечностей, на 50-й задние конечности разделяются на суставы, на 62-й они приобретают подвижность, на 69-й становятся видимыми передние конечности, и на 71-й начинается резорбция хвоста. Развитие может затягиваться до 133 дней. Известны случаи зимовки головастиков. Рост их отличается значительной интенсивностью (в среднем 0,9 мм в день). К моменту метаморфоза длина головастика достигает длины тела половозрелой самки. Средние размеры сеголеток 30-32 мм, вес 3,4 г.
В популяциях прудовых лягушек выделяются три возрастные группы. Соотношение полов таково: самцов 31,4%, самок 68,6%. Половозрелость наступает на 3-й год.
Колебания численности этого вида изучены мало. В отличие от бурых лягушек они меньше страдают от засухи. Однако в Дарвиновском заповеднике за 1947-1949 гг. численность прудовых лягушек изменилась значительно, сократившись в 4 раза. В известной мере это объясняется местной спецификой условий существования. Численность прудовой лягушки падает здесь в годы с низким уровнем водохранилища, когда спад воды в июне влечет за собой пересыхание мелких водоемов и как следствие гибель головастиков. По некоторым данным, при пересыхании водоемов прудовые лягушки зарываются на дне их и, покрытые подсохшей грязью, как бы впадают в спячку.
Икру прудовой лягушки поедают кряквы, головастики отмечены среди кормов обыкновенной чайки, взрослых уничтожают белокрылые крачки, выпи, сарычи, филины, камышницы.
К группе зеленых лягушек относится также чернопятнистая лягушка (Rana nigromaculata). Внутренний пяточный бугор у нее высокий, сжатый с боков, между спинно-боковыми складками большое число продольных кожных ребрышек. Сверху она серовато-оливкового цвета с большим числом иногда сливающихся черных пятен. Вдоль середины спины проходит продольная светлая полоса. Низ тела белого цвета. Иногда попадаются особи с темным височным пятном (около 4%). В углах рта у самца наружные резонаторы серого или почти белого цвета. Максимальная длина тела 95 мм. Она уменьшается по мере продвижения вида на север и запад. Чернопятнистая лягушка живет в Китае, Восточной Монголии, Корее, Японии и в пределах нашей страны на Дальнем Востокег на север до 55° с. ш. Интересно, что у этого восточного вида крупные экземпляры населяют восточную часть ареала, тогда как у западного вида - прудовой лягушки - они обитают на западе.
Чернопятнистая лягушка держится в водоемах, часто на рисовых полях. Просыпается в конце марта - начале апреля. Икрометание в марте - апреле, обычно по утрам. Самка откладывает около 5000 яиц диаметром 1,7 мм. Головастик перед метаморфозом составляет около 71 % от длины взрослых. На зимовку уходит в октябре. По своему образу жизни близка к озерной лягушке.
Остромордая лягушка (Rana terrestris) - многочисленный вид в нашей фауне, относящийся к группе бурых лягушек. Внутренний пяточный бугор у нее высокий, сжатый с боков, морда заостренная. Сверху она коричневая или сероватая с темными пятнами и точками. Это делает ее малозаметной среди травы, гниющих листьев, хвои, палочек и сучков в тех местах, где она обычно живет. От глаза через барабанную перепонку почти до плеча у нее тянется темное постепенно суживающееся височное пятно. Это пятно хорошо маскирует глаз лягушки, который легче всего делается заметным у затаившегося животного и выдает его присутствие. Горло у остромордой лягушки беловатое, большей частью с мраморным рисунком. Брюхо белое или желтоватое, в подавляющем большинстве случаев без пятен. Общий тон окраски остромордой лягушки может значительно меняться в зависимости от температуры и влажности среды. В сухую и солнечную погоду наблюдается заметное его посветление. На севере Горьковской области в народе существует примета, что лягушки светлеют к хорошей погоде. Весной у самцов развивается яркая серебристо-голубая окраска и все тело становится раздувшимся, отекшим. Среди бесхвостых земноводных средней полосы остромордая лягушка единственная имеет такой ярко выраженный брачный наряд. На первых пальцах передних лап у самца имеются темные шероховатые брачные мозоли, не расчлененные на части. Плавательная перепонка на задних лапах развита у них в период размножения лучше, чем после него, когда лягушки уходят из водоемов. Относительная площадь ступни (площадь ступни, деленная на длину тела и умноженная на 50) в период размножения у них увеличивается на 80%. У самок разрастание перепонки выражено значительно меньше. Площадь ее ступни изменяется всего лишь на 8%.
Максимальный размер, которого может достигнуть остромордая лягушка,-78 мм. Однако обычная длина половозрелых особей - от 51 до 70 мм. Географические закономерности изменения длины тела этого вида не установлены. Однако пропорции тела лягушек из разных мест обитания не одинаковы. Например, относительная длина задних ног у самцов растет с юга на север, у самок таких изменений не происходит. Лягушки из лесотундры и тундры не подчиняются этой закономерности. Они отличаются короткими задними конечностями. Пропорции тела животных изменяются не только в зависимости от их мест обитания или пола, но и от возраста. Так, у самцов с возрастом относительная длина ног становится больше. Однако у самых старых самцов в ряде случаев наблюдаются обратные изменения, относительная длина конечностей становится меньше. Интересно, что животные, появившиеся на свет в разные годы, могут различаться между собой пропорциями тела больше, чем обитающие в разных географических пунктах. Иными словами, существуют изменения в строении тела по годам. Все это характеризует сложные взаимоотношения организмов с окружающей средой и важно для изучения эволюции вида.
Остромордая лягушка распространена на запад до Северо-Восточной Франции, обитает в Бельгии, Голландии, Дании, далее западная граница ее ареала постепенно переходит в северную, проходя через Южную Швецию, Финляндию, Карелию, и выходит к берегам Белого моря, проходит через низовья Печоры, юг полуострова Ямал, переходит в нижнем течении Енисей и спускается на юг до Тувы. Южная граница идет по Алтаю, через Северный Казахстан, пересекает реку Урал у Уральска, нижнее течение Волги, Дона, Днепра, проходит через Румынию, Венгрию, верховья Дуная и Рейна. В Крыму и на Кавказе отсутствует.
Остромордая лягушка населяет лесную, лесостепную и степную зоны. В Северном Казахстане заходит в полупустыню, встречается и в тундре. В горы поднимается до 700 м. Большая часть ареала остромордой лягушки совпадает с ареалом травяной, однако границы ее распространения раздвинуты к югу.
По сравнению с травяной лягушкой отличается несколько меньшими требованиями к влажности. Посаженные в террариум на сухой песок, травяные лягушки гибнут на второй-третий день, а остромордые живут больше недели. В местах, где влажность 81-90%, травяная лягушка встречается редко (23% встреч), а остромордая значительно чаще (40,9% встреч). Видимо, этим в известной мере объясняется более широкое проникновение остромордой лягушки в зону степей.
В тундре остромордая лягушка распространена в значительно меньшей степени, чем травяная. Не поднимается она и в горы на Полярном Урале. Видимо, устойчивость ее к низким температурам меньше, чем у травяной.
В лесной зоне оба эти вида должны быть отнесены к многочисленным. На 100-метровом маршруте в хвойных лесах в среднем можно встретить двух остромордых лягушек, а в лиственных - четырех. На севере остромордая лягушка встречается реже травяной, а на юге она над ней преобладает.
Оба вида в известной мере делят между собой территорию. Это показывают одновременные подсчеты численности остромордой и травяной лягушек на одних и тех же маршрутах, проходящих по разным биотопам. Наблюдения эти были проведены в Костромской, Владимирской и Горьковской областях. В различных типах боров и по степным склонам попадались только остромордые лягушки, травяные здесь не встречались. В елово-пихтовом лесу, на ржаном поле среди елово- пихтового леса, в дубовом лесу и по оврагам с мелким кустарником, напротив, отсутствовали остромордые лягушки, а встречались травяные. И в Ярославской области, а также в Дарвиновском заповеднике остромордые лягушки более многочисленны в бору-зеленомошнике, чем в ельнике-зеленомошнике.
Значительно большая связь остромордой лягушки с сосной, а не с елью вновь подтверждает ее меньшую требовательность к влажности. Сосна обычно растет на песках, влагоемкость которых близка к 2%, тогда как у суглинистых и глинистых почв, характерных для еловых и смешанных лесов, влагоемкость достигает 15%.
Там, где травяная лягушка преобладает над остромордой, последняя занимает более сухие места обитания. Там, где остромордая лягушка многочисленнее травяной, она занимает и ее биотопы, в первую очередь различные типы лиственных лесов.
В пределах своего распространения она встречается в очень разнообразных типах лиственных лесов, в осиновых, липово-дубовых, дубовых, буковых лесах и ольшаниках. Обитает в пойменных лесах и березовых колках. Придерживается опушек и полян. В Волжско-Камском заповеднике самая большая численность остромордой лягушки наблюдалась в осиннике. Здесь за 10 суток в ловчую канавку ловилось до 165 лягушек, в дубово-липовом лесу с примесью березы, клена, вяза, ели и с обильным травостоем - 86, а в березняке - 32. В сосново-еловом лесу не ловилось более 15 лягушек.
В открытых биотопах лесной зоны, в суходольных лугах и на степных склонах остромордая лягушка встречается реже, чем в лесах. Здесь на 100 м маршрута встречается менее одной лягушки. Однако в пойменных лугах численность этого вида значительна - до 4 лягушек на 100 м учетной линии. Довольно часто остромордая лягушка живет на болотах, особенно по их окраинам, предпочитает осоковые, но не избегает и сфагновых. На сфагновых болотах численность этого вида примерно такая же, как на суходольных лугах.
Остромордая лягушка относится к группе наземных лягушек и не только проводит на суше большую часть активного периода, но и, как правило, зимует. Однако на пределах своего распространения в степях и в тундре она не порывает связи с водоемами и после периода размножения.
Выходит на охоту вечером и активно кормится между 20-22 часами, несмотря на то что температура воздуха в это время понижается. Это связано с тем, что ночью влажность выше. После полуночи активность начинает медленно снижаться. С 4 часов и до 18 она держится на низком уровне. Однако остромордую лягушку чаще, чем других наземных позвоночных, можно видеть деятельной и днем.
Весной, в период размножения, когда лягушки держатся в водоемах или по их берегам, характер их поведения меняется. Период пониженной активности сокращается и длится примерно с 4 до 10 часов. Лягушки оказываются малодеятельными лишь в самое холодное время суток, а днем и в первую половину ночи они активны. Максимум их деятельности, так же как и летом, развивается между 20-24 часами. В это время наблюдается максимальное число спаривающихся особей, чаще звучит брачная песнь и больше откладывается икры. Весной активность, связанная с процессами размножения, подавляет все другие виды деятельности. Лягушки мало кормятся, у них наблюдается "брачный пост".
В неактивное время суток лягушки весной скрываются на дне водоемов, где колебания температуры менее резки, чем в воздухе, а летом прячутся в более влажных местах, под упавшими деревьями, в пнях и т. п.
В степи и в тундре, где озерные лягушки не отрываются от водоемов и после периода размножения, весенний характер активности сохраняется у них и летом.
Основу питания остромордой лягушки составляют жуки. Другие корма у лягушек из разных географических пунктов имеют разное значение. В одних случаях значительную долю в питании, кроме жуков, составляют пауки, кобылки, клопы и гусеницы; в других к этим кормам присоединяются комары, но падает значение клопов или исчезают и комары и клопы, но появляются муравьи. Иногда же все другие корма, кроме жуков, отличаясь значительным разнообразием, встречаются в желудках не часто, в небольших количествах, и трудно какому-нибудь из них отдать предпочтение.
Состав кормов может меняться не только в разных географических пунктах, но и в соседних биотопах. Под Казанью в лесах к доминирующим кормам можно отнести жуков (48,9%), пауков (29,2%), кобылок (27,7%), гусениц (15,4%) и клопов (14,9%). В тех же местах в пойме набор преобладающих кормов сокращается. К ним относятся жуки (72,0%), пауки (44,0%) и гусеницы (16,0%). Остальные корма встречаются лишь в 4% исследованных желудков.
В питании остромордых лягушек очень велико значение наземных животных. Они составляют в средней полосе 91,2% от всех встреченных в желудках кормов. Интересно, что и в зоне степей, где лягушка постоянно держится у водоемов, она питается исключительно наземными организмами. В тундре значение водных кормов в питании этого вида возрастает.
Различие в кормах остромордой и травяной лягушек определяется тем, что первая обитает в более сухих местах, чем вторая. Например, травяная лягушка больше поедает наземных моллюсков, тоже придерживающихся более влажных мест.
Применением мечения остромордых лягушек удалось установить, что кормовой участок отдельной особи занимает площадь до 0,2-0,3 га. Обычно лягушки не уходят далее 25-30 м от того места, где были помечены. В пределах этого участка животное постоянно передвигается в поисках пищи. Кормовые участки разных лягушек, обитающих по соседству, перекрываются. Размеры кормового участка и приверженность к нему лягушек определяются его кормностью. Если запасы пищи становятся недостаточными или изменяются условия влажности, ограничивающие активность животных, лягушки предпринимают миграции, перемещаясь в другие места. Передвижения совершаются постепенно со скоростью от 3 до 20 м в сутки, при этом могут смениться не только кормовые участки, но и биотопы. Такие перемещения могут свершаться и в течение нескольких недель, и на протяжении двух и более активных сезонов.
Интенсивность питания остромордых лягушек, как и у других земноводных, зависит от степени их активности, которую ограничивают условия температуры и влажности. Понижение температуры осенью приводит к тому, что лягушки все чаще и чаще попадаются со слабо наполненными или совсем пустыми желудками. Постепенное ослабление активности приводит в конце концов к зимней спячке.
Таблица 13. Черепахи: 1 - болотная (Emys orbicularis); 2 - каспийская (Clemmys caspica); 3 - звездчатая (Testudo elegans); 4 - средиземноморская (Testudo graeca); 5 - расписная украшенная (Chrysemys picta); 6 - красноухая (Pseudemys scripta); 7 - бугорчатая (Maiaclemys terrapin); 8 - матамата (Chelus fimbriata)

Таблица 13. Черепахи: 1 - болотная (Emys orbicularis); 2 - каспийская (Clemmys caspica); 3 - звездчатая (Testudo elegans); 4 - средиземноморская (Testudo graeca); 5 - расписная украшенная (Chrysemys picta); 6 - красноухая (Pseudemys scripta); 7 - бугорчатая (Maiaclemys terrapin); 8 - матамата (Chelus fimbriata)
Кормовые летние миграции неизменно переходят осенью в миграции к местам зимовок, которые у этого вида выражены не ярко.
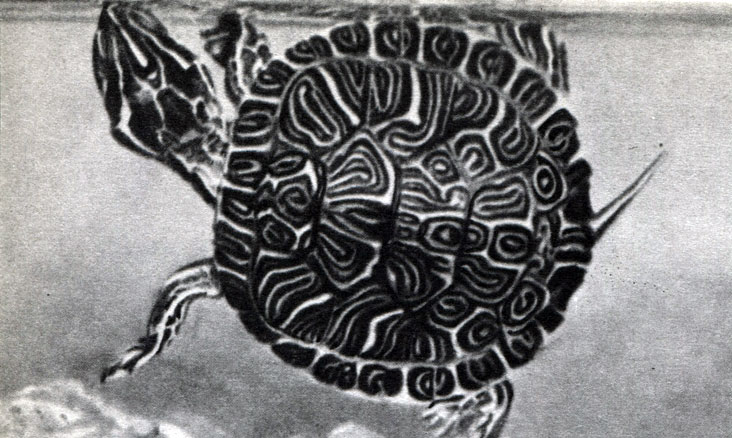
Таблица 14. Флоридская украшенная черепаха (Pseudemys floridana) со спинной стороны
Таблица 14а. Украшенная черепаха (Pseudemys dorbifjni) с брюшной стороны

Таблица 14б. Европейская болотная черепаха (Emys orbicularis)
Большинство остромордых лягушек зимует на суше: в ямах, засыпанных листьями, в кучах листвы и хвои, под кучами хвороста, в норах грызунов и т. п. Меньшее число зимовок встречается в незамерзающих ручьях, лесных речках, богатых родниками, и в торфяных болотах.
На зимовку уходят в северных районах в начале сентября, южнее - в конце октября, примерно на две недели раньше, чем травяные. Срок зимней спячки в среднем 165-170 дней, на 10-15 дней больше, чем у травяной. Это, видимо, связано с меньшей устойчивостью остромордой лягушки к низким температурам. Молодые уходят на зимовку позже взрослых.
Просыпаются остромордые лягушки под Киевом в середине марта, под Москвой в середине - конце апреля. В холодные весны выход с зимовки может задерживаться до начала мая. В тундре активность начинается значительно позднее, к размножению остромордые лягушки приступают лишь в середине июня. Неполовозрелые появляются позднее взрослых. Период активности этого вида в течение года в Подмосковье занимает 135 дней, а в Северной Буковине - 210.
Половозрелые особи переходят из мест зимовок в водоемы. Эти передвижения проходят в массе очень быстро - за 3-4 дня. Собираясь в нерестовые водоемы, лягушки проходят значительные расстояния - до 800 м. За сутки они могут пройти до 300 м.
Пришедшие в водоем лягушки без промедления приступают к размножению. Минимальная длина участвующих в размножении самок - 42,5 мм, самцов - 43,4 мм. Половозрелость наступает на 3-й год. Отмечено, что лягушки, живущие во влажном и теплом климате, начинают размножаться, достигнув меньших размеров по сравнению с лягушками, живущими в другом климате. Более или менее длительное время в водоемах проводят самцы, подстерегающие приходящих в водоем еще не отметавших икру самок. Отдельные самцы могут оставаться в водоеме до 20-25 дней. Самки же не только приходят в водоем позднее самцов, но и, отметав икру, сейчас же покидают его. Этим объясняется то, что в водоемах обычно встречается большее число одиночных самцов и только спаривающиеся, не отложившие икру самки. В самый разгар размножения на суше можно встретить или отдельных, еще не отложивших икру самок, идущих к водоему, или, напротив, самок, уже удаляющихся от него. Самцы в это время на суше не встречаются.
Уходящие из водоемов лягушки снова проходят большие расстояния, но так как они в это время интенсивно кормятся, восстанавливая затраченную в период размножения энергию, скорость движения их невелика - до 16 м в сутки.
У степных лягушек, в течение всего активного периода размножения не уходящих от водоемов, сроки икрометания растягиваются до месяца, тогда как у лесостепных и лесных популяций на это требуется 10-15 дней.
Собравшиеся для размножения в водоеме самцы образуют большие скопления. Иногда на мелководье на 1 м2 можно насчитать их до 25 штук. Булькающий крик этих животных создает иллюзию бегущего весеннего ручья или кажется похожим на отдаленный лай собак. В некоторых случаях самки, отметавшие икру, имеют травмы после крепких брачных объятий. Площадь кожи, сорванной лапками самца на груди самки, доходит до 4 см2.
Кладка в .виде одного, реже двух или трех комков откладывается недалеко от берега на мелких, незатененных, хорошо прогреваемых местах. Она обычно около суток лежит на дне, а затем всплывает. В одном месте часто скапливается большое количество икры, отложенной многими самками.
Водоемы, в которых мечут икру остромордые лягушки, разнообразны, чаще лесные, с поросшим травой дном. Нередко нерестовыми водоемами служат торфяные болота.
Одна самка откладывает 504-2750 яиц. Количество их зависит от возраста животного. С увеличением его возрастает число откладываемых яиц. Однако это происходит лишь до определенного предела. У самок, достигающих размеров 69-70 мм, плодовитость вновь падает.
Диаметр икринки 6-8 мм, диаметр яйца 1,5-2,0 мм, но может быть и менее - до 1,0 мм.
Температура воды, при которой начинается откладка икры, равна 12,0-14,8°. Выход личинок из икры наблюдали на Украине через 3 дня после ее откладки. В Татарии это происходит через 5-10 дней. При изменениях температуры воды от 4 до 23° головастики выходят из икры через 8-10 дней. Колебания температуры воды, в которой развивается икра остромордой лягушки, бывают очень большими. Случается, что она лежит в воде, сверху покрывшейся корочкой льда. В таких случаях развитие икры задерживается, но она не гибнет. Это объясняется большой устойчивостью яиц к воздействию низкой температуры. Важное значение имеет также и тот факт, что температура в комке икры в среднем в течение суток на 3° выше, чем температура окружающей среды. Минимальная разница, приходящаяся на самое холодное время суток, 1,5°. Полностью набухшая оболочка яйца содержит только около 1 % сухого вещества, остальное составляет вода, среди всех других веществ отличающаяся самой большой теплоемкостью. Обладая большой теплоемкостью и концентрируя на себе, подобно собирательным линзам, световые и тепловые лучи, прозрачные слизистые оболочки яиц аккумулируют большое количество тепла. Тепловая инерция в комке икры объясняется также и малой теплопроводностью оболочек. Икра нагревается сильнее и быстрее воды, а остывает дольше. Усиленному поглощению тепловых лучей способствует также скопление темного пигмента на одном полюсе яйца, обращенном к свету. В то же время пигмент служит ширмой, ограждающей яйцо от вредного действия ультрафиолетовых лучей.
Выклев головастика из яйца происходит благодаря растворяющему оболочки яиц ферменту, который выделяют одноклеточные железы зародыша.
На протяжении всей своей жизни головастики остромордой лягушки образуют скопления и не расплываются по водоему, придерживаясь мелководий.
О питании их известно очень мало. Вероятно, они предпочитают животную пищу. Ротовая воронка у них менее глубокая, бахрома по ее краям мелкая, роговые челюсти значительно уже, чем у растительноядных головастиков озерной лягушки. Зубчики на губах обычно мелкие.
У только что вылупившейся личинки части тела едва обозначены. Голова отделена от туловища легким перехватом, а задний конец зародыша вытянут в коротенький хвост (рис. 15). Хвост окружен широким плавником, идущим и вдоль спины личинки. Головастики окрашены в черный цвет и достигают 5,5-7,5 мм длины.
Вскоре после вылупления развиваются наружные жабры, отличающиеся значительной длиной. Они сильно ветвятся и сохраняются дольше, чем у других наших лягушек. Все это, видимо, связано с тем, что, живя большими скоплениями, головастики испытывают недостаток кислорода.
В первую половину личиночного развития, до появления зачатков конечностей, когда усиленно идут процессы формирования различных органов, головастики остромордой лягушки увеличиваются за день на 0,4 мм. Наибольшей интенсивности рост животного достигает в промежуток времени от появления зачатков конечностей до разделения задних конечностей на отделы, т. е. как раз в то время, когда процессы формообразования приближаются к концу и ослабевают. В это время личинки увеличиваются приблизительно на 0,7 мм в день. Затем интенсивность роста вновь падает, и перед метаморфозом головастики вырастают в день на 0,4 мм.
Среди других наших лягушек остромордая характеризуется наименее интенсивным ростом.
Перед метаморфозом длина тела головастиков (35-45 мм) составляет около 67% от длины тела взрослой самки. Их сравнительно небольшие размеры находятся в соответствии с непродолжительным личиночным развитием. Все личиночное развитие занимает в среднем 60-65 дней, но в исключительных случаях может растягиваться и до 120 дней. Продолжительность метаморфоза 4 дня. Тундровые популяции остромордых лягушек отличаются очень быстрым развитием. Максимальная продолжительность его 45-55 дней, несмотря на то что температура воды, в которой живут головастики, далека от оптимальной.
Только что метаморфизировавшие сеголетки имеют длину тела 13-20 мм. Северные формы в этом отношении, видимо, не отличаются от южных. Пол в разных зонах дифференцируется на разных этапах развития. В популяциях, обитающих в степном Зауралье, он различим уже у сеголеток длиной 19-20 мм. В лесостепи, в лесу, в лесотундре Зауралья и в горах Южного Урала пол можно различить только на второе лето их жизни, а в тундре еще позднее. Видимо, на формирование пола оказывает влияние температура окружающей среды. Чем она ниже, тем позднее дифференцируется пол животного.
С момента метаморфоза до ухода на зимовку лягушата в Волжско-Камском заповеднике вырастают в среднем на 3,4 мм, а за 6 зимних месяцев - всего на 1,1 мм. Их рост зимой происходит в 8 раз медленнее, чем летом. На Южном Урале лягушата в первое лето жизни вырастают с 13 до 24-25 мм. Только что окончившие метаморфоз лягушки с Полярного Урала имеют размеры тела тоже около 13 мм, но не успевают за первое лето достичь размеров своих южных сородичей. И в дальнейшем они, видимо, растут медленнее их. На это указывают максимальные размеры остромордых лягушек, отмеченные у тундровых: лягушек (55,4 мм) и южных (60,2 мм).
Расселение молодых из водоемов, как правило, начинается в конце июня - в июле и происходит со сравнительно большой скоростью. Эти крошечные животные преодолевают за сутки 25-60 м.
В популяции остромордой лягушки в весенний период отчетливо выделяются по размерам три возрастные группы: годовалые лягушки длиной 25 мм, двухлетние - длиной до 42 мм и старшего возраста, имеющие размер более 42 мм, Соотношение численности этих возрастных групп, видимо, не одинаково в разные годы. В Дарвиновском и Волжско-Камском заповедниках в 1947 и 1950 гг. преобладающей группой оказалась вторая. Однако в 1936 г. под Звенигородом наиболее многочисленной была третья группа. Соотношение этих возрастных групп изменяется и в течение одного сезона. Все эти изменения объясняются разной интенсивностью гибели лягушек, происходящей под воздействием разнообразных причин.
Общая численность остромордых лягушек вследствие их гибели может измениться в 4-9 раз, если сравнивать между собой разные годы. Сокращение численности происходит главным образом от засухи, когда погибает масса икры и головастиков, а также в суровые малоснежные зимы на зимовках. Однако у них имеются и враги и паразиты. На головастиков; нападают плавунцы и другие водные хищники из беспозвоночных. Из позвоночных животных их уничтожают зеленые лягушки и ужи, особенно молодых.
Взрослых лягушек поедают озерные лягушки, ужи, гадюки, аисты, каравайки, малые выпи, речные чайки, малые подорлики, сарычи, вороны и даже глухари. У барсуков эти лягушки встречаются в 56% исследованных желудковг нападают на лягушек и выдры, норки,, хори, ласки, лисицы, ежи и даже обыкновенные бурозубки и кроты. Однако у этих животных они встречаются всего лишь в 0,6-19% желудков.
У остромордой лягушки обнаружено 25 видов внутренних паразитов: 16 видов плоских червей, 8 - круглых и 1 вид скребней. К головастикам присасываются большая и малая ложноконские и улитковые пиявки. У взрослых лягушек наружные паразиты не найдены. Этим они,, видимо, обязаны ядовитым выделениям кожных желез. В оболочках икринок остромордых лягушек содержится особое вещество - ранидин, губительно действующее на простейших. Оно вдвое сильнее карболовой кислоты.
В народной медицине сушеная икра лягушек используется при лечении рожистого воспаления лица.
Травяная лягушка (Rana temporaria) по внешнему виду очень напоминает остромордую, но отличается от нее более крупными размерами (до 100 мм), темным мрамороподобным рисунком на брюхе, тупой мордой и низким внутренним пяточным бугром. В брачное время у самца голубеет горло, а на первом пальце передних ног становятся ясно заметны четырехраздельные черные шершавые бугры.
Населяет всю Европу, исключая Пиренейский полуостров, на север доходит до пределов континента, южные границы ее распространения - юг Франции и Италии. В Крыму, на Кавказе и в низовьях Волги отсутствует. На восток едва переходит за Урал. В горы поднимается до 3000 м. Типичная лесная форма, в Европе травяная лягушка встречается и в лесостепи, заходя в степи лишь по поймам рек. Этот многочисленный вид все лето проводит на суше, удаляясь от водоемов на значительные расстояния, но населяя лишь влажные биотопы.
Расселение травяных лягушек по суше определяется их зависимостью от влажности. В этом отношении они занимают промежуточное положение между зелеными лягушками и жабами. Они способны без вреда для себя терять большее количество воды, чем прудовые лягушки, но значительно меньшее, чем жабы, особенно зеленые. Проницаемость их кожи для воды также меньшая, чем у жаб, но большая, чем у прудовой лягушки. Степень проницаемости кожи для воды регулирует ее отдачу телом в окружающую среду. Меньше всего воды пропускает кожа у животных, побывавших на солнце, когда сверху на теле образуется тоненькая пленка подсохшей слизи. Проницаемость кожи отличается и географической изменчивостью. Травяные лягушки из областей с большей влажностью обладают кожей, более проницаемой для воды.
У северных пределов своего обитания в связи с низкими температурами воздуха и на юге, где велика сухость, травяная лягушка держится у воды.
Как и другие земноводные, она избегает соленых водоемов и не может прожить более суток в воде, соленость которой достигает 0,07%.
О распределении травяной лягушки по биотопам было сказано при характеристике образа жизни остромордой. Добавим лишь, что остромордая лягушка оттесняет травяную в общем направлении с юго-востока на северо-запад. Основными причинами отступания травяной лягушки, видимо, оказывается некоторое потепление климата и воздействие антропогенных факторов, особенно вырубка лесов. Вследствие этого температура и влажность изменяются в сторону более благоприятную для остромордой лягушки. В этом смысле остромордую лягушку можно считать следующей за культурным ландшафтом, который для травяной лягушки менее благоприятен.
Травяные лягушки редко попадаются на глаза днем. В это время они сидят затаившись в густых кустах, под камнями, в пнях, в густой траве - словом, там, где больше влажность. Часто, приподнимая упавшие деревья, можно увидеть под ними одну или несколько лягушек. Они сидят, тесно прижавшись к земле, и находятся в состоянии легкого оцепенения. Проходит некоторое время, прежде чем потревоженные животные обращаются в бегство. На день в поисках более влажных убежищ лягушки могут перемещаться и из одного биотопа в другой. Так, во влажном осиннике днем наблюдалось больше травяных лягушек, чем на соседнем сухом водораздельном лугу. Ночью же большинство из них выходило на охоту на луг.
Активная деятельность у травяных лягушек начинается с наступлением сумерек, максимума она достигает с 23 до 2 часов, затем число деятельных животных падает, достигая минимума к 11 часам. Бодрствующие ночью лягушки усиленно кормятся. Их желудки оказываются наиболее наполненными в 4-8 часов, т. е. непосредственно после ночного периода активности.
По некоторым наблюдениям, кривая активности травяных лягушек, так же как и остромордых, имеет двувершинный характер. Первый пик наблюдается в 21-22 часа, затем активность резко снижается и вновь достигает вершины в 3 часа, после чего она постепенно падает, чтобы к утру дойти до минимума. Перерыв ночью приходится на самое темное время, а максимум активности соответствует вечерним и утренним сумеркам. Такой характер активности, видимо, связан с продолжи-тельными ночами и, следовательно, зависит от времени года и географического положения местности. В те дни, когда температура воздуха выше, охотящихся лягушек встречается больше. Наибольшая их активность наблюдается в июльские и августовские ночи, отличающиеся самой высокой температурой воздуха.
Несмотря на то что травяные лягушки предпочитают высокую температуру, активность их приходится не на самый теплый период суток. Происходит это оттого, что для бурых лягушек, не связанных вне периода размножения с водоемом, влажность окружающей среды имеет решающее значение. Во время их активности влажность наибольшая из наблюдающейся в течение суток. Хорошим подтверждением этому служит тот общеизвестный факт, что после дождя и обильной росы лягушки проявляют значительно более оживленную деятельность и выходят на охоту и днем. В Заполярье травяную лягушку с одинаковой степенью вероятности можно встретить активной днем и ночью. Сеголетки, как правило, активны днем.
Главную роль в питании травяной лягушки (73%) играют жуки и двукрылые, затем наземные моллюски и прямокрылые. Подавляющую массу корма эти животные добывают на суше (94,2%). Следовательно, хотя список кормов травяной лягушки и велик (87 форм), но основу питания составляет несравненно меньшее число массовых организмов.
Летающих животных в пище травяной лягушки около 16%, т. е. несколько меньше, чем у остромордой. Это, видимо, связано с тем, что остромордая лягушка чаще, чем травяная, охотится днем, когда больше активных летающих насекомых. У северной границы ареала вида травяные лягушки, теснее связанные с водоемом, употребляют в пищу большее количество водных организмов. Интенсивность питания в разное время года неодинакова. Весной, в период размножения, у них наблюдается "брачный пост". Однако это явление еще не изучено и не выяснено, отсутствует ли питание полностью, как ведут себя в этом отношении самцы, самки и неполовозрелые особи.
С наступлением осени число лягушек, имеющих в желудке пищу, также постепенно сокращается. У взрослых это происходит быстрее, чем у молодых. Травяные лягушки перестают быть активными с наступлением регулярных заморозков, когда средняя суточная температура воздуха становится ниже 6°, а температура воды превышает температуру воздуха и колеблется в пределах от 6 до 10°.
Молодые уходят на зимовку на одну-две недели позднее взрослых. Они встречаются еще и в середине ноября при дневной температуре 0°. Различное поведение взрослых и сеголеток объясняется различной устойчивостью их к низким температурам. В то время как взрослые в условиях опыта не выносят переохлаждения тела ниже минус 0,4-0,8°, сеголетки оказываются устойчивыми к охлаждению до минус 1-1,1°, а может быть, и ниже. При более низких температурах у сеголеток число дыхательных движений в минуту значительно больше, чем у взрослых, но с повышением температуры эти различия сглаживаются.
Среди наших земноводных травяные- лягушки отличаются непродолжительным сроком зимней спячки. В среднем она длится 155 дней. Меньше спят только обыкновенные тритоны и жерлянки. Срок зимней спячки находится в связи с отношением животного к температуре. Температура тела у травяных лягушек в природе колеблется от 6,0 до 24,5°, у остромордых - от 10,5 до 27,5°. Размах колебаний температуры тела у первой составляет 18,7°, у второй - 17°. Травяная лягушка спит зимой меньше остромордой, видимо, потому, что она живет в интервале более низких температур и может переносить более широкий размах ее колебаний.
Продолжительность спячки меняется в зависимости от географического положения местности. Под Киевом она равна 130-150 дням, под Москвой - 180-200, под Архангельском - 210-230.
Осенью, когда средняя суточная температура воздуха колеблется в пределах от 8 до 12°, а минимальная падает до минус 5°, травяные лягушки группируются в местах, близких к их будущим зимовкам: в заболоченных участках, прилегающих к водоемам, в придорожных канавах, в зарослях осоки по берегам речек и т. п. К местам зимовок они двигаются по канавам, ручейкам или сильно увлажненным местам, избегая сухих и открытых пространств. По ручейкам и канавкам животные движутся как по течению, так и против него, причем совершают миграции преимущественно днем. На пути своего движения лягушки часто останавливаются. Скорость движения их по суше в среднем 3-4 м в минуту. Расстояние, преодолеваемое за все время миграции, по имеющимся наблюдениям, не превышает 1,5 км. Путь этот лягушки проходят за один день. Весь процесс скопления лягушек у мест зимовок обычно длится не более 2-3 дней. Места осеннего скопления обычно связаны с зимовками водным путем и лежат от них не далее чем в 100-150 м.
В годы высокой численности травяной лягушки перемещения к местам зимовок могут принимать форму заметных миграций.
Осенние передвижения лягушек, очевидно, вызваны не только падением температуры воздуха ниже температуры водоемов, но также сезонными изменениями кормовой базы. К этому времени начинают исчезать наземные насекомые и в питании лягушек усиливается роль водных беспозвоночных.
Почти любой не промерзающий до дна водоем может служить местом для зимовки травяной лягушки. Однако она предпочитает прежде всего не сильно каменистые, быстро текущие, незамерзающие речки, затем торфяные канавы и болота с жирным илом. Меньше всего зимовок можно встретить в больших реках, если в них нет тихих заводей. Сильный весенний разлив очень затрудняет выход лягушек из таких рек на сушу. Наконец, в больших реках больше хищных рыб, истребляющих зимой значительное количество лягушек. Немногочисленны также зимовки в озерах и прудах, особенно в сильно загрязненных, мелких, непроточных прудиках, где животные гибнут от недостатка кислорода и большого количества выделяющихся вредных газов.
Обычно описываемое зарывание лягушек в ил относится только к прудовой лягушке. Травяные располагаются либо просто на дне водоема, либо под нависающими ?берегами, либо в зарослях растительности, а в проточных водах и под камнями.
На зимовке травяная лягушка сидит в очень типичной позе, поджав задние лапы, а передними как бы закрывает голову, вывернув их ладонями наружу. При этом на ладонях очень хорошо заметна густая сеть кровеносных сосудов, вследствие чего ладони всегда ярко-розовые.
Зимовки в стоячих водоемах, как правило, располагаются около незамерзающих стоков или ключей. Здесь лучше условия аэрации и меньше сероводорода. В некоторых случаях, когда условия зимовки оказываются неблагоприятными, местоположение ее в течение зимы меняется. Перемещение может происходить на расстоянии до 120 м.
В проточных водах незамерзающий сток - не столь обязательное условие зимовки. Однако и там обычно наблюдается скопление лягушек у места впадения в реку ручья или притока.
Такое расположение зимовок предохраняет земноводных от заморов - главной опасности, грозящей их благополучию в водоемах. Заморы возникают в результате резкого снижения содержания кислорода в воде, расходующегося в процессах гниения органических остатков. Зимующие в воде лягушки могут гибнуть и при промерзании водоемов до дна.
Однако если водоем не промерзает, то условия зимовки в нем оптимальны для земноводных. Угрозы высыхания здесь не существует вообще, а температура никогда не падает ниже нуля, и колебания ее незначительны. В торфяных канавах и ямах температура не падает ниже 3°, а в некоторых ключах, где зимуют лягушки, температура всю зиму держится в пределах 6-8°.
Число зимующих в одном месте травяных лягушек различно. В ряде случаев это бывают одиночные особи, в некоторых случаях число их может достигать нескольких сотен. Чаще всего встречаются зимовки, состоящие из двух-трех десятков экземпляров. Самцы, самки и молодь зимуют вместе.
Зимующие лягушки вялы, но не лишены способности к движению. Желудок у них не всегда бывает пуст. В некоторых случаях до 10% исследованных животных имели в желудках различных водных беспозвоночных, обрывки элодеи, спирогиры и других водорослей, а также семена и собственную кожу, сброшенную во время линьки. Предположение, что зимнее содержание желудков травяных лягушек представляет собой остатки пищи, проглоченной осенью, маловероятно, так как скорость переваривания у них при температуре 0,5-2° равна 72-120 часам, а ниже температура тела на зимовках не падает. Несмотря на то что рост травяных лягушек зимой резко замедляется, он, так же как и развитие половых продуктов, все же не прекращается совсем. Следовательно, процессы жизнедеятельности у лягушек во время зимнего сна не прекращаются, а только крайне замедляются. При понижении температуры тела почти в 2 раза уменьшается число дыхательных движений. Потребление кислорода при одной и той же температуре (20°) во время спячки в 2 раза меньше, чем в период активности. При 0° выделение углекислоты в 20 раз меньше, чем при 25,5°.
Однако одно замедление жизнедеятельности не может обеспечить выживаемость на зимовках под водой. В летний период лягушка, содержащаяся при 2°, погибает без легочного дыхания через 8 суток, несмотря на то что животное находится в депрессии и жизнедеятельность его сильно понижена. Во время же спячки лягушки живут только за счет кожного дыхания пять месяцев и более. Это возможно благодаря ряду изменений в организме. В летние месяцы понижением температуры до 0° вызвать у лягушек зимнюю спячку не удается именно благодаря существованию отличий в строении и физиологии у "зимних" и "летних" животных. Так, в печени у них еще с осени откладывается запасное питательное вещество - гликоген. Зимой увеличивается в коже число капилляров и у гемоглобина почти вдвое повышается сродство к кислороду, понижается проводимость и возбудимость нервных путей, положительный гелиотропизм сменяется отрицательным и т. д. Известно, что при температуре около 0° у лягушек увеличивается содержание воды в тканях, так как прекращается выделение ее почками, но продолжается поступление через кожу. Видимо, благодаря этому вес земноводных за период спячки не уменьшается, а в некоторых случаях даже увеличивается.
Все эти факты показывают, что явление зимней спячки не простая реакция на понижение температуры или влажности, а сложная цепь взаимосвязанных изменений в организме, выработавшихся исторически как приспособление.
Условия зимовки для земноводных в водоеме более благоприятны, чем на суше, однако и здесь значительная часть зимующих животных гибнет, не пережив трудное время года. Например, в 1938 г. около 20% находившихся под наблюдением в окрестностях Москвы зимовок травяной лягушки погибло полностью.
Просыпающиеся раньше других травяные лягушки первыми откладывают икру. В среднем откладка икры под Москвой у них начинается 22 апреля. За одиннадцать лет наблюдений самые первые кладки наблюдались 7 апреля, самые поздние - 3 мая. Икрометание начинается очень скоро после пробуждения, через 3-5 дней. Спаривание у травяной лягушки начинается еще по пути к нерестовым водоемам. В это время у самок вся икра уже овулировала и находится в последнем тонкостенном, растянувшемся отделе яйцеводов, готовая к откладке. У всех половозрелых особей вида созревание и откладка яиц происходят более или менее одновременно. Выметав икру, лягушки долго не задерживаются в водоемах, расходясь по летним местам обитания. В период размножения у самцов более чем в полтора раза увеличиваются размеры плавательных перепонок на задних лапах. У самок, так же как и у других лягушек, перепонки увеличиваются мало.
Кладка травяной лягушки имеет типичную для всех лягушек форму комка, образовавшегося благодаря склеиванию слизистых яйцевых оболочек и содержащего от 670 до 1400 икринок. Свежеотложенную икру в водоеме легко узнать, так как она представляет собой небольшой комочек тесно прилегающих друг к другу яиц. Постепенно, по мере разбухания слизистых оболочек, расстояние между отдельными яйцеклетками увеличивается и весь комок приобретает значительно больший объем. Яйца склеиваются только там, где они соприкасаются, в других местах между ними остаются каналы, так что комок икры по своей структуре похож на гроздь винограда. Промежутки между икринками способствуют свободному проникновению кислорода к каждому из развивающихся зародышей. Эти каналы сохраняются только в том случае, когда комок находится в воде во взвешенном состоянии. В комках, опустившихся на дно, по крайней мере часть каналов нарушается, и нормальное развитие икринок в этих участках оказывается невозможным. Это важно учитывать при развитии икры в лабораторных условиях. Воды в сосудах должно быть такое количество, которое необходимо для того, чтобы комок свободно плавал в ней. Кислородный режим развивающихся яиц улучшается и тем, что на слизистых оболочках их поселяются водоросли, выделяющие в процессе фотосинтеза кислород.
Раннее размножение травяных лягушек приводит к тому, что кладки их иногда можно наблюдать в водоемах, еще полностью не освободившихся ото льда. Установлено, что яйца этого вида выдерживают переохлаждение до минус 6° и не теряют способности к развитию. Несмотря на это, развитие икры травяных лягушек ранней весной возможно только благодаря тем приспособлениям, которые общи у нее с остромордой лягушкой.
Обладая способностью развиваться при низкой температуре, яйца травяной лягушки не могут без вреда для себя долго выдерживать температуру около 24-25°. Некоторые исследователи полагают, что южная граница распространения травяной лягушки определяется именно этим обстоятельством. Так, изучение ее в Пиренеях, где живут одиночные представители этого вида, привело к заключению, что южная граница распространения травяной лягушки совпадает с июльской изотермой 21°. В Англии известны случаи, когда наступление жаркой погоды во время нереста задерживало икрометание. В результате в поздно отложенных перезревших комках икры наблюдался большой процент погибших яиц. Предполагается, что при значительном повышении температуры окружающей среды зародыши гибнут от недостатка кислорода, так как потребность в нем с повышением обмена веществ возрастает, а форма кладки в виде большого комка препятствует аэрации каждого яйца. Однако, если учитывать описанную выше структуру икряного комка, это маловероятно. Яйца травяных лягушек, обитающих на юге ареала, отличаются большей устойчивостью к высоким температурам, чем у северных популяций.
Скорость развития находится в прямой зависимости от температуры. Чем выше она, тем быстрее идет развитие. В среднем головастики травяной лягушки выклевываются из яиц через 8-10 дней после их откладки. В глубоких, затененных водоемах икра развивается примерно в четыре раза медленнее, чем в хорошо прогреваемых водоемах. Однако при одних и тех же температурных условиях в эксперименте скорость развития яиц травяной лягушки по сравнению с другими нашими лягушками оказывается наибольшей.
Развитие головастиков у травяной лягушки занимает 50-90 дней. При более высоких температурах оно происходит быстрее. Температурный оптимум лежит в пределах 21-26°. Однако, подобно остромордой лягушке, развитие травяной на Полярном Урале идет очень быстро , 43-50 дней. Температура воды, в которой живут головастики, колеблется здесь, от 0 до 22° и чаще всего равна 10-15°, т. е. далека от оптимальной. Быстрый темп развития у северных популяций - это приспособление к существованию' в местах, где лето очень коротко.
В эксперименте при одинаковых условиях развития рост головастиков травяных лягушек, так же как и у других видов, достигает максимума в период ослабления процессов дифференцировки органов, что приходится главным образом на время от появления почек конечностей до полного расчленения задних конечностей на отделы. В среднем темп роста головастиков травяной лягушки лишь немногим больше, чем у остромордой (0,6 мм в день). Размеры головастиков этого вида перед метаморфозом малы. Длина их составляет лишь 55% от длины половозрелых самок.
Более или менее подробно жизнь головастиков травяной лягушки в естественных условиях изучалась в Англии в разных участках различных водоемов на протяжении трех лет. Наблюдения проводились еженедельно.
Большую часть своей жизни головастики живут колониями, образуя большие скопления. Плотность их внутри колонии может достигать 100 штук на 100 мм2.
Примерно через месяц-полтора после вылупления они рассеиваются по водоему, начиная вести одиночный образ жизни. В некоторых случаях, главным образом при небольшой численности головастиков, скопления их исчезают значительно раньше. Кормятся они на мелководьях, среди водорослей, на растительной пленке, покрывающей водоемы, и на дне их. Колонии перемещаются в поисках более кормных, не занятых другими участков. Так, на местах икрометания головастики то исчезают, то появляются снова. Возможно, что более крупные особи занимают лучшие места обитания, оттесняя из них более мелких. На протяжении трех лет оказывалось, что головастики, держащиеся среди водорослей, имели больший вес, чем те, которые были собраны из других участков этого же пруда. Выпадение осадков, способствующее заливанию мелководий, таких, где обыкновенно происходит икрометание, также сказывается на увеличении веса личинок, здесь кормящихся. Обычно же головастики на местах икрометания весят меньше переместившихся в другие участки.
С развитием растительности в прудах темп роста головастиков ускоряется. В начале своей жизни они растут медленнее и кормятся, видимо, главным образом на дне.
Численность головастиков в разные годы в одном и том же пруду при более или менее одинаковом количестве отложенной икры может существенно изменяться. Так, головастики, населяющие пруд в 1948 г., составляли лишь 1% от популяции 1947 г. Темп их роста даже в одном и том же водоеме также неодинаков в разные годы. Несмотря на то что в 1948 г. головастики вылупились раньше, чем в 1947 г., их максимальный вес к 10 мая был в 2 раза меньше, чем вес головастиков в это же самое время в 1947 г. Наибольший вес головастиков перед метаморфозом, по наблюдениям, во всех прудах за все годы изменяется от 500 мг до 1 г. В отдельных случаях головастики весят и более 1 г.
Как правило, рост продолжается до конца июня, а затем кривая нарастания веса головастиков резко идет вниз. В это время основная масса личинок метаморфизирует и остаются в личиночном состоянии, видимо, отставшие в росте и развитии. Падение веса в период метаморфоза - явление, характерное для земноводных. К концу июля головастики перестают встречаться в водоемах.
Однако этот наиболее типичный ход развития может значительно нарушаться. В 1947 г. в одном из прудов головастики уже к 20 мая достигли значительных размеров, 400-500 мг. В начале июня стали попадаться первые сеголетки. Тем не менее в массе метаморфоз не наступал, и вес головастиков то понижался, то- повышался, оставаясь примерно на одном и том же уровне с 21 мая по 29 июня. Затем он значительно возрос (до 700 мг), начался массовый метаморфозой к 1 августа личинок в водоеме уже не было. В этом же году в другом пруду развитие головастиков затянулось еще более, приняв катастрофический характер. За исключением небольшого числа метаморфизировавших в конце июня, большая часть популяции до октября оставалась на личиночной стадии, постепенно истощаясь и теряя в весе. В другие годы в этом пруду численность головастиков была сильно разреженной, но они быстро росли и развивались. Однако число метаморфизировавших сеголеток в этом водоеме в течение всех лет оказывалось малым. Малая продуктивность этого водоема, видимо, объясняется плохими кормовыми условиями в нем для головастиков. В некоторых прудах метаморфоз задерживался, несмотря на незамедляющийся темп роста.
Причины, определяющие все разнообразие хода развития головастиков, еще очень мало известны.
После метаморфоза рост травяных лягушек продолжается до трех лет и более. Половозрелость наступает на третий год. Известны случаи, когда в неволе травяные лягушки доживали до 18-летнего возраста. Однако продолжительность их жизни в природе значительно меньшая - 4-5 лет.
Особенно велика смертность в период развития. Смертность яиц и головастиков в сумме составляет 80,4-96,8%.
Численность травяных лягушек в разные годы значительно изменяется. Так, с 1939 по 1942 г. в средней полосе Европейской части СССР она возросла более чем в 45 раз; напротив, с 1936 по 1939 г.- неуклонно падала. Изменение численности может происходить одновременно на значительной территории. Сопоставление границ территории, на которой происходило снижение численности травяной лягушки в 1936-1939 гг., с границей засухи, бывшей в эти годы, показало, что засуха оказалась основной причиной гибели животных. Иссохшими черными трупиками головастиков были усеяны потрескавшиеся ложа высохших водоемов. Пересыхание болот, ранний листопад и сухость лесной подстилки привели к гибели большого количества сеголеток и взрослых лягушек.
Другой причиной, определяющей падение численности травяной лягушки, были сильные морозы при малоснежной зиме 1938/39 г. В этом году нередко встречались зимовки, промерзшие до дна. Гибель амфибий зимой может достигать, по-видимому, значительных размеров. Есть указания на большой процент гибели их в зиму 1928/29 г. Наконец, известно, что суровая зима 1828/29 г. привела к резкому снижению численности земноводных почти во всей Европе и полному исчезновению их в Исландии.
Травяная лягушка менее чувствительна к суровым зимам, чем остромордая. Это объясняется тем, что она зимует в водоемах. Однако остромордая лягушка, как более сухолюбивая форма, оказалась несколько больше устойчивой к засухе.
Так как засуха и морозы по-разному сказываются на численности этих близких видов, то изменение численности их не всегда совпадает по времени и масштабам.
Гибнут травяные лягушки и от хищников. Икру лягушек поедают некоторые птицы: серая утка, свиязь, кряква, камышница, большой веретенник, черная крачка. Головастики отмечены в пище сизоворонок, сороки, дрозда-рябинника и белобровика; взрослые входят в состав кормов обыкновенной чайки, черного аиста, ястреба-тетеревятника, сарыча, осоеда, подорлика, болотного луня, филина, мохноногого сыча, ворона, серого сорокопута, жулана.
Из бурых лягушек, отличающихся хорошо развитым темным височным пятном, идущим от глаза через барабанную перепонку, у нас в стране обитает еще 5 видов: сибирская (Rana cruenta, или R, chensinensis), закавказская (R. camerani), малоазиатская (R. macrocnemis), прыткая (R. dalmatina) и дальневосточная (R. semiplicata). Биология этих видов изучена мало. У всех у них внутренний пяточный бугор низкий, не сжатый с боков. Окраска тела сверху светло-бурая с большим или меньшим количеством темных пятен. У сибирской и закавказской лягушек часто бывает светлая полоса вдоль спины. У сибирской лягушки на брюхе кроваво- красные пятна, откуда она и получила свое латинское название (cruenta в переводе значит "обрызганная кровью"). У других видов брюхо однотонное, красное или розовое. Самая маленькая из них сибирская, максимальная длина которой 66 мм, несколько крупнее дальневосточная - 79 мм, еще крупнее малоазиатская и прыткая, достигающие 80 мм длины, и самая большая - закавказская, имеющая длину до 90 мм. Малоазиатская и прыткая лягушки отличаются также значительной длиной задних ног.
Сибирская лягушка населяет Сибирь, Северо-Восточный Казахстан, Северную Киргизию, на Дальнем Востоке встречается в Приморье, Приамурье, на Сахалине и Шантарских островах. На западе граница ее распространения проходит между 70 и 80° в. д. На юг спускается до Центрального Китая, на север доходит до тундры. На восток от Урала по лесной и лесостепной полосе она как бы заменяет травяную и остромордую лягушек. Подобно последней, встречается и в степях и в полупустынях. В южных частях ареала держится только вблизи водоемов. Имеющиеся сведения по биологии этого вида собраны главным образом в Казахстане. Численность сибирской лягушки в окрестностях Алма-Аты от 500 до 800 особей на 1 га. Основной корм - насекомые. Ранней весной часто встречаются водные насекомые, в другое время года, как правило, только наземные. Вредные насекомые составляют 50-70%.
Сибирская лягушка уходит на зимовку во второй половине октября - в начале ноября. Зимует в зарослях заболоченных водоемов, в колодцах-копанцах и на суше недалеко от воды: в ямах с гниющей растительностью, в расщелинах почвы, в норах грызунов и т. п. Весной появляется в марте - начале апреля. Жизнедеятельна 7-8 месяцев в году. Вскоре после пробуждения, не более чем через 10 дней, приступает к икрометанию. Брачный период длится от двух недель до месяца. Самцы редко издают негромкие звуки. ?Спаривание происходит под водой. Откладывает 1000-1600 икринок. Яйцо окрашено в темно-бурый цвет. Диаметр яйца 1,7-2,3 мм, а икринок - 5-7 мм. Местом икрометания служат водоемы, расположенные в поймах рек, неглубокие, слабозаболоченные, медленно текущие источники, колодцы и арыки. Икра обычно откладывается при температуре воды 18°. Головастики вылупляются через 6-10 дней, в это время они достигают длины 7-12 мм. Головастики, ведущие уже подвижной образ жизни, сверху темно-серые с мелкими пятнышками и крапинками бурой окраски; с нижней стороны одноцветные, серые и тело их очень прозрачно. К концу развития длина головастиков колеблется в пределах от 37 до 60 мм. Питаются они фито- и зоопланктоном и детритом. Корма растительного происхождения составляют 20-25%. Длина только что метаморфизировавших сеголеток составляет 13-17 мм. Выход лягушат на сушу происходит в последних числах мая. Развитие занимает от 25 до 40 дней. В течение месяца размеры сеголеток увеличиваются на 7-10 мм и к концу лета длина их достигает 33 мм.
На юго-востоке Казахстана лет 20-30 назад сибирская лягушка была многочисленной, а сейчас заметно сократилась в числе. Возможно, это связано с тем, что на протяжении последних 50 лет в бассейн Балхаша проникла озерная лягушка, вытесняющая сибирскую лягушку.
Закавказская и малоазиатская лягушки очень похожи друг на друга, и не раз возникал вопрос о том, можно ли считать их разными видами. Однако подробный анализ особенностей их строения подтверждает принадлежность их к разным видам. Об этом же свидетельствуют и физиологические их особенности. Мышечная ткань этих двух видов теряет возбудимость при разной температуре, и закавказская лягушка оказывается более устойчивой по отношению к воздействию высоких температур. Споры вокруг этих двух видов привели к тому, что и до сих пор не всегда можно решить, к какому из них относятся те или иные биологические особенности, приводимые в литературе.
Закавказская лягушка распространена от Южного Дагестана через Восточное Закавказье, включая Талыш, до плоскогорий Армении. В горы поднимается до высоты 3210 м над уровнем моря. Приспособленная к сухопутному образу жизни, закавказская лягушка может жить при потере воды до 29,5% общего веса тела. Она держится вдали от водоемов, собираясь возле них только в период икрометания и осенью перед уходом на зимовку.
Основные корма закавказской лягушки принадлежат к наземным формам, 70-80% приходится на долю жесткокрылых. Около 10% от всех встреченных экземпляров составляют гусеницы. Более 50% поедаемых животных - вредители.
Зимуют закавказские лягушки в водоемах обычно по нескольку экземпляров, зарывшись в ил на глубину 30-40 см. Концентрация возле водоемов осенью происходит тогда, когда средняя температура воздуха достигает 6-7°, Уходить в водоемы закавказские лягушки начинают при температуре 4-5° и исчезают полностью при 3-4° мороза. В горах на высоте 1760-2000 м над уровнем моря это случается в начале ноября; на высоте 1300 м во второй половине ноября. Молодые лягушки уходят на зимовку позднее, встречаясь до начала декабря при средней температуре воздуха минус 1-2°.
Весной закавказские лягушки появляются в начале, а в высокогорных районах в конце марта. Продолжительность спячки у них в разных географических точках изменяется от 100 до 140 дней.
Подобно другим бурым лягушкам, закавказские проявляют большую устойчивость к низким температурам, чем зеленые лягушки. Это проявляется в том, что они позже озерных лягушек впадают в спячку и раньше просыпаются, а также выше идут в горы. В соответствии с этим они более чувствительны к высоким температурам. Их мышечная ткань скорее теряет возбудимость при повышении температуры, чем у озерных лягушек. Однако если сравнить по этому показателю закавказских и травяных лягушек, то первые оказываются более устойчивыми по отношению к высоким температурам, что связано с их более южным распространением.
В размножении закавказские лягушки участвуют, достигнув 50-55 мм длины. Самцы составляют 60% популяции. У этого вида очень своеобразно выражен половой диморфизм, развивающийся в период икрометания и выражающийся в том, что брачная окраска самки ярче, чем у самца. Верх тела самки становится розоватым, а брюшко - ярко-оранжево- красным. Самцы в это время серые или буроватые. Розовые у них только нижние части бедра и каемка на брюшке. Первый палец передних ног у самцов черный. Самки крупнее самцов.
Днем в водоемах эта лягушки незаметны. Икра выметывается ночью, одной или реже двумя порциями. Одна самка откладывает от 3500 до 5000 икринок. У самок длиной 85 мм диаметр яйца равен 2 мм, у меньших - от 1,5 до 1,8 мм. Температура воды во время икрометания бывает от 4 до 14°. На высоте 980 м над уровнем моря откладка икры начинается со второй половины марта, а на высоте 1940 м - в конце апреля.
Развитие зародыша при температуре воды 5-8° продолжается около 10 дней. Покинувшие оболочку яиц темно-коричневые головастики имеют длину 9-10 мм. На 2-3-й день после вылупления появляются наружные жабры, затем прорывается рот и начинают функционировать внутренние жабры. Примерно на 20-25-й день развития, когда головастики станут длиной 23-25 мм, появляются зачатки конечностей. На 50-55-й день левая передняя конечность выходит наружу через жаберное отверстие, а правая - прорывая жаберную крышку. Хвост рассасывается на протяжении 6-7 дней. При колебании температуры воды от 5 до 23° развитие у закавказской лягушки длится 60-70 дней. Длина лягушонка после метаморфоза равна 14-15 мм, а перед уходом на зимовку - 30-35 мм. В низменных местах сеголетки появляются в середине мая, в высокогорных районах - во второй половине июня.
Малоазиатская лягушка встречается в Малой Азии, на Черноморском побережье Кавказа и в Предкавказье. Изредка попадается на высотах до 3500-4000 м. В Боржомо-Бакурианском районе она наиболее многочисленна на высоте от 1500 до 1700 м. В Азербайджане эта лягушка обычно держится на высоте 700-1200 м. Они населяют горные леса и лесостепи, встречаются в садах.
В водоемы малоазиатские лягушки собираются чаще всего только в период икрометания и на зимовку, однако, видимо, не избегают их и в другое время. Так, в окрестностях Ставрополя днем их можно видеть только в холодной воде лесных ручьев. В Боржомо-Бакурианском районе они выходят из водоема в 9-10 часов, удаляются от берега на 40-60 м и охотятся на суше до 17-18 часов, затем вновь собираются на берегу водоема. Молодые лягушки возвращаются к воде на час-полтора позже.
Малоазиатские лягушки в большей степени, чем другие бурые лягушки, употребляют в пищу водных животных (23,9%), оказываясь по этому показателю в одном ряду с зелеными лягушками. Преобладающие корма у них представлены жуками, личинками двукрылых и водными ракообразными. Среди жуков первое место в питании этой лягушки занимает род Bimbidion, представители которого всегда держатся у воды.
Суммарный процент преобладающих кормов у них велик (72,3%), так же как и у других бурых лягушек. Летающие насекомые в пищу употребляются мало. Однако все это может быть характерно только для молодых лягушек, питание которых проанализировано в большей степени, чем у взрослых.
В период размножения наблюдается "брачный пост".
На зимовку в окрестностях Ставрополя и в Боржомо-Бакурианском районе малоазиатские лягушки уходят в конце сентября, в Азербайджане - в октябре, а изредка и в начале ноября. Зимуют большими скоплениями в тихих родниках ущелий. Однако есть указания о возможности зимовок этой лягушки и на суше.
На низменностях появляются в середине марта, в горах - в конце апреля. Для этого вида описаны такие же изменения окраски в брачный период, как и для закавказской. Самцы во время размножения довольно сильно урчат, напоминая по голосу травяных лягушек.
Прыткая лягушка отличается стройным телом, узкой головой и необычайно длинными задними ногами. Если ее заднюю ногу вытянуть вперед, то голеностопное сочленение идет далеко за конец морды. Глаза у прыткой лягушки большие, выпуклые, барабанная перепонка расположена очень близко к глазу и по размерам едва ему уступает. Сверху прыткая лягушка розовато-бежевого или светло-бурого цвета с темными пятнами. На задних лапах темные пятна располагаются довольно четкими полосами. Брюхо всегда белое, у живых с заметно розовым оттенком. Через некоторое время после поимки оно становится ярко-розовым. В период размножения у самцов на первом пальце появляются серые брачные мозоли. Резонаторов не имеют. Голос слабый.
Прыткие лягушки исключительно подвижны. Они делают прыжки 1-1,5 м в длину и до 1 м в высоту. Уходя от преследования, способны совершать скачки до 3 м.
Прыткая лягушка населяет западную, среднюю и юго-восточную части Европы от северо-востока Испании и Франции на восток до Малой Азии. Северные пределы распространения - Дания, острова Рюген, Борнхольм и крайний юг Швеции; южные пределы - остров Сицилия, Апеннинский полуостров и Пелопоннес. В СССР прыткая лягушка встречается только в районах, прилегающих к Восточным Карпатам. В горы идет до 1500 м над уровнем моря, но чаще встречается на равнине. На всем протяжении своего ареала прыткая лягушка немногочисленна. Ведет сухопутный образ жизни. Самки удаляются от воды дальше, чем самцы. Излюбленные места обитания - луга с густой и высокой травой, лесные поляны в буковых и смешанных лесах, заросли кустарников в долинах и реже сады. Сухих участков не избегает, но предпочитает места с влажностью от 65 до 80%. Активна в сумерки, а во влажных местах и днем.
В питании этого вида преобладают жуки. пауки, двукрылые, равнокрылые и перепончатокрылые. Корма добывает почти исключительно на суше. Водные формы представлены личинками двукрылых и ветвистоусыми рачками. Вредные насекомые составляют 41,5% от числа поедаемых особей.
На зимовку уходят в середине или конце октября. Зимуют зарывшись в ил на дне водоемов.
В Закарпатье весной появляются во второй половине марта, немного позже, чем травяная и остромордая, что свидетельствует о большой теплолюбивости этого вида. Икра не переносит низких температур. Прыткие лягушки приступают к спариванию только тогда, когда температура воды поднимается до 4-5°. Одна самка откладывает от 600 до 1400 икринок. Диаметр яйца 2-3 мм, а всей икринки - 9-12 мм. Верхняя половинка яйца бурая или черноватая, нижняя желтоватая или грязно-белая.
Сперматозоиды прыткой лягушки сходны со сперматозоидами травяной лягушки и сильно отличаются от сперматозоидов остромордой, на которую по внешним признакам и анатомическому строению прыткая лягушка похожа больше.
Развитие головастика продолжается 2-3 месяца. Наибольшая длина головастика 55-60 мм. Метаморфоз заканчивается в августе. Только что закончившие превращение лягушата имеют длину тела 13-20 мм.
Как и другие представители рода, водный образ жизни ведет и бугорчатая лягушка (Rana rugosa), обитающая в Японии, Корее и в южной части Приморского края. Она достигает 56 мм длины, кожа ее сверху бугорчатая. Верхняя сторона тела окрашена в тусклый серо-бурый цвет, в задней части тела переходящий в зеленый. Брюхо грязно-белое с черными мраморными разводами. Голос этой лягушки - негромкое ворчание, которое раздается и ночью и днем, как в период икрометания, так и после него.
Другая водная лягушка тропическая прибрежная лягушка (Rana limnocharis) широко распространена в Юго-Восточной Азии. В горы поднимается до 2000 м. Длина ее редко превышает 50 мм. Верхняя сторона тела оливково-зеленая или оливково-бурая; пятнистый рисунок травянисто-зеленый или темно-бурый; полоса, проходящая по средней линии спины, бывает то узкая желтая или травянисто-зеленая, то широкая оранжевая; иногда она вовсе отсутствует. Нижняя сторона белая, губы в темно-бурых пятнах. В тропической части провинции Юньнань это самый многочисленный вид лягушек открытых ландшафтов. По окраинам рисовых полей их встречается в 4 раза больше, чем в прилежащем лесу. Активны ночью, когда больше активных насекомых. Популяция этих лягушек представлена двумя возрастными группами: сеголетками (18-32 мм) и взрослыми (более 34 мм). Этот вид характеризуется быстрым ростом и ранним наступлением половой зрелости - в годовалом возрасте. Вероятно, основная часть популяции обновляется через год, поскольку лягушки старше одного года составляют менее 2% популяции. Размножение приурочено к периоду дождей - с мая по август. Отличается очень высокой плодовитостью. Икринки величиной с просяное зерно образуют овальные комки. Головастики вылупляются через 48 часов.
Индийская тигровая лягушка (Rana tigrina), которая может достигать величины 150 мм, по окраске и внешнему виду очень похожа на предыдущий вид, но отличается от него лучше развитыми продольными складками на спине, которые часто выступают острым углом. Употребляется в пищу. Под Кантоном есть ферма по разведению этой лягушки в искусственных прудах.
Одна из самых красивых лягушек - красноухая лягушка (R. erythraea) обитает* на Малайском полуострове и прилежащих островах. Она отличается стройным тело- сложением, имеет явственные пластинки для прилипания на пальцах обеих пар конечностей. Сверху она зеленая с металлическим блеском, с боков темно-коричневая. Продольные складки спины: у этой лягушки серебристо-белые, барабанная перепонка красная; верхняя половина радужной оболочки золотисто-желтая, нижняя огненно-красная. Этот многочисленный вид селится в водоемах болотах и на рисовых полях. Сезонность в размножении не выражена. У самцов изменения в интенсивности сперматогенеза и в развитии брачных мозолей в течение года незначительны. У самок также на протяжении всего года обнаружены яйца на разных стадиях созревания. Однако процент самок и самцов, готовых к размножению, изменяется в различные месяцы от 10 до 50.
У коротконогой лягушки (Rana curtipes), распространенной в лесах Западной Индии, развитие головастиков продолжается с июля по март. Питаются они исключительно растительной пищей. К октябрю длина их тела достигает 5 см, а длина кишечника - 20 см. В октябре - январе морфологической дифференцировки у головастиков не происходит, но они энергично растут и в январе достигают 11 см (длина кишечника 28 см). К концу января появляются зачатки задних конечностей и начинается редукция кишечника. В феврале заканчивается формирование конечностей и начинается рассасывание хвоста.
По всей Южной и Тропической Африке, а также на Мадагаскаре и в Северо- Восточной Африке распространена ведущая водный образ жизни нильская лягушка (R. mascareniensis), достигающая 40-48 мм длины. Верхняя сторона тела у нее оливково-зеленая, бурая или серо-зеленая в более темных пятнах, нижняя - белая; задняя сторона бедер белая в мраморных разводах. Вдоль спины может быть светлая полоса. Нильская лягушка играла большую роль в египетской мифологии. Божество Ка, имевшее голову лягушки, представляло собой одно из видоизменений бога правды Пта. Кроме того, была еще богиня Хека с лягушачьей головой, которая вместе со своим супругом, богом Кхнумом, олицетворяла воду. Лягушка была символом воскресения. Головастик обозначал в иероглифическом письме число сто тысяч. В древних Фивах найдены были даже бальзамированные нильские лягушки.
В Африке же обитает самый крупный изо всех известных лягушек вид - лягушка-голиаф (Rana goliaph), имеющая в длину 250 мм и больше и достигающая веса 3,25 кг. Она имеет очень ограниченное распространение, населяя территорию шириной примерно в 100 км вдоль побережья республики Камерун и Рио-Муни.
Лучше других изучены лягушки, населяющие водоемы Северной Америки.
Самая крупная из них - лягушка-бык (R. catesbeiana) имеет длину 200 мм. Сила голоса лягушки-быка находится примерно в таком отношении к силе голоса наших зеленых лягушек, в каком находятся размеры этих амфибий. Лягушка-бык отличается большими размерами барабанной перепонки, которая не уступает по величине глазу, а у самцов даже превосходит ее. Оливково-бурая или оливково-зеленая верхняя поверхность тела покрыта большими темно-бурыми и черными пятнами; нижняя часть тела желтовато-белая, одноцветная или с мраморным рисунком. Радужная оболочка красноватая с желтым ободком. У этого вида хорошо развиты плавательные перепонки и задние ноги достигают в длину 25 см. Продольные спинные складки отсутствуют. Лягушка-бык распространена в восточной части Северной Америки, на юге более многочисленна, чем на севере. Нигде не образует таких больших скоплений, как наши зеленые лягушки. Предпочитает густо поросшие кустарником берега рек, отличающихся чистой водой. От опасности спасается прыгая в воду. Главную пищу составляют насекомые, пауки, а также моллюски. В зависимости от пола и по месяцам состав пищи не изменяется, По мере роста лягушек количество насекомых в их пище снижается, доля растительных остатков увеличивается. В водоемах в открытой местности средний вес содержимого желудков больше, чем в водоемах, расположенных в лесу. Вероятно, в первом случае добыча более доступна для лягушек.

Рис. 64. Лягушка-бык (Rana catesbeiana)
Благодаря своим размерам лягушка- бык бывает и настоящим хищником, поедая всех других животных, которых в состоянии осилить: рыб, других земноводных, птенцов и т. п.
В Канаде развитие головастиков длится 2 года. После метаморфоза в разных популяциях скорость роста, время полового созревания и соответственно максимальные размеры тела неодинаковы.
Похожа на лягушку-быка значительно меньшая по размерам крикливая лягушка (Rana clamitans). Верхняя сторона тела этой лягушки окрашена спереди в серый цвет, сзади в оливково-зеленый; горло лимонно-желтое, брюхо белое; спина и передние ноги в бурых пятнах, а задние ноги в перевязях того же цвета. Кожа грубая, шероховатая. В отличие от лягушки-быка имеются спиннобоковые складки. Средние размеры крикливой лягушки достигают 47 мм. Максимальная скорость роста наблюдается в первый год жизни, и дальнейшее увеличение ее размеров незначительно. Эти обитатели водоемов не встречаются дальше чем на 18 м от воды. Метаморфоз наблюдается в первой половине июля.
Леопардовая лягушка (R. pipiens), окрашенная в разные оттенки зеленого цвета и, подобно другим зеленым лягушкам,, не имеющая темного височного пятна, идущего через барабанную перепонку, ведет наземный образ жизни, предпочитая влажные места. В этом отношении она похожа на бурых лягушек. Размеры ее достигают 75-90 мм. Она имеет обширный ареал, занимающий большую часть Северной и Центральной Америки. Благодаря высокой подвижности , выживаемости и относительно меньшей требовательности к влажности леопардовая лягушка, видимо, расселяется на север, северо-восток и северо-запад по долинам рек, сухим долинам и междуречьям, вытесняя сходный с ней по образу жизни вид - пятнистую лягушку (Rana pretiosa). Последняя более требовательна к влажности, но более вынослива по отношению к низким температурам. Проникновение леопардовой лягушки на север тормозится ее приверженностью к более высоким температурам. Вместе два эти сходных вида долго жить не могут и представляют собой типичный случай видов-двойников.
В желудках леопардовых лягушек 15% объема пищи составляют личинки чешуекрылых, 9% - улитки, 4% - мокрицы. Известны случаи нахождения в ее желудке летучих мышей. Летом в хорошую погоду леопардовые лягушки обычно 95% времени суток остаются в своих убежищах, некоторые держатся там более 24 часов и даже до 5 суток. Передвижение их по индивидуальным участкам обычно не превышает 5-10 м. Такие передвижения происходят в любое время суток, но почти 2/3 общего расстояния проходят в темноте. Движение лягушек внутри индивидуального участка образует сложную сеть пересечений, петель и сдваиваний маршрутов. При ночных дождях лягушки иногда предпринимают значительные передвижения, проходя при этом 100-160 м. С рассветом миграции прекращаются, но могут продолжаться следующей ночью. Одна лягушка за две ночи прошла 240 м. При сильных продолжительных дождях почти вся популяция лягушек мигрирует. Из 30 лягушек, отловленных и помеченных во время дождей или после них вне своих индивидуальных участков, 25 позднее были найдены вновь на своих прежних местах или на пути к ним. Наибольшая зарегистрированная скорость миграции - 46,5 м в час. Пройденное расстояние и скорость передвижения сильно зависят от температуры. Метаморфоз в первой половине июля. R. pipiens и R. pretiosa отличаются незначительной скоростью роста.
Придерживаясь влажных мест, ведет наземный образ жизни и маленькая бурая лягушка - лесная лягушка (R. silvatica), проникающая к северу в Америке дальше всех других видов земноводных. Зимует на суше. В умеренном климате она приступает к размножению раньше всех других видов лягушек. На Аляске ее размножение в течение 12 лет начиналось между 24 апреля и 18 мая. В течение трех дней, непосредственно предшествовавших началу размножения, средняя суточная температура составляет 6,1°. У истоков реки Миссисипи она размножается в прудах на возвышениях, а затем расселяется в низинные болота. Сюда же приходят и метаморфизировавшие сеголетки. Размеры индивидуальных участков, на которых они остаются все лето, в среднем составляют 69,5 - 72,3 м2. Многие лягушки, повторно отловленные через год, находились вблизи мест прошлогодней поимки: на расстоянии 14-29 м. На елово-лиственничном, торфяном болоте лесная лягушка ведет дневной образ жизни. Максимумы ее деятельности наблюдаются между 8 и 10 и между 16 и 18 часами. Степень и длительность ее активности прямо пропорциональны влажности воздуха. Молодые лягушки предпочитают более влажные места, чем старые лягушки. Рост этих животных особенно интенсивен в молодом возрасте и почти прекращается ко времени полового созревания. В период размножения рост останавливается. Темп его также замедляется при снижении температуры и недостатке пищи. Самки, как и у большинства других земноводных, растут несколько быстрее и к половозрелости достигают больших размеров.
Рис. 65. Лесная лягушка (Rana sylvatica)
Семейство Веслоногие лягушки (Rhaeophoridae, или Polypedatidae)
Это семейство включает около 400 видов, объединяемых в 10 родов, из которых около 150 видов, главным образом родов Polypedates и Rhacophorus, обитает в Южной и Юго-Восточной Азии, Малайском архипелаге, 250 видов - в Центральной и Южной Африке и на Мадагаскаре (род Hyperolius - около 190 видов, род Chiromantis - 12 видов, Hylombates - около 15 видов, Megalixalus - 30 видов, Kassina - 4 вида и др.).
Большинство видов веслоногих лягушек ведет древесный образ жизни, замещая квакш, от которых они отличаются рядом особенностей строения скелета.
Наиболее примитивен род Rhacophorus, некоторые виды которого известны своей способностью совершать планирующие прыжки, за что они получили название летающих лягушек. Между пальцами передних и задних конечностей у них находятся очень сильно развитые плавательные перепонки. Когда животное прыгает, оно растопыривает пальцы, раздувает тело и легко планирует вниз. На концах пальцев у них имеются расширенные диски. У некоторых видов наблюдали прыжки в 10-12 м.
Таблица 16. Черепахи: 1 - бисса (Eretmochelys imbricata); 2 - суповая зеленая черепаха (Chelonia mydas); 3 - логгерхед (Caretta caretta); 4 - кожистая черепаха (Dermochelys coriacea)

Таблица 16. Черепахи: 1 - бисса (Eretmochelys imbricata); 2 - суповая зеленая черепаха (Chelonia mydas); 3 - логгерхед (Caretta caretta); 4 - кожистая черепаха (Dermochelys coriacea)
Нередкая в горных лесах Явы и Суматры яванская веслоногая лягушка (Rh. reinwardti) достигает 75 мм длины. Сверху она окрашена в интенсивно-зеленый цвет, снизу ярко-желтая. У нее имеется кожный вырост на пятке, кожная оторочка, тянущаяся по предплечью и по краю пятого пальца задних конечностей; заднепроходное отверстие также покрыто кожной лопастью. У молодых большие интенсивно-синие пятна помещаются между пальцами на плавательных перепонках обеих конечностей, а также в подмышечных впадинах. У взрослых лягушек большинство этих пятен исчезает. Самец гораздо меньше самки. Его окраска меняется быстрее, чем у самки. Днем яванская веслоногая лягушка впадает в особое состояние, напоминающее сон. Этому состоянию предшествуют быстрые и сильные дыхательные движения, а вслед за этим лягушки как будто съеживаются и дыхательные движения становятся медленными и редкими.
Рис. 67. Яванская веслоногая лягушка (Rhacophorus reinwardti)
Период размножения у нее с января до августа, разгар его - в марте. Икра откладывается в пенистых комках на листья, висящие над водой. Пена из студенистой оболочки яиц взбивается задними ногами животного. В кладке содержится 60-70 непигментированных яиц, 3 мм в диаметре.
Японская веслоногая лягушка (Rh.buergeri) известна своим голосом, ее кваканье напоминает пение птиц. В Японии этих певцов продают на рынках, и лучшие из них дорого стоят. Живут они в быстро текущих горных потоках и своими присосками на пальцах удерживаются на камнях, прыгая с одного на другой.
Другой вид - Rh. schlegelii, обитающий также в Японии, размножается с середины апреля до середины мая. Неся на своей спине гораздо более мелкого самца, самка закапывается под вечер в землю на отвесном берегу около водоема и делает здесь шаровидную нору, имеющую 6-9 см в диаметре. Здесь во взбитую ногами лягушки пену откладываются яйца размером около 6 мм. Мало-помалу пенистая масса становится жидкой и вместе с головастиками вытекает из норы по тому самому ходу, через который вышли родители. Этот ход направляется наклонно вниз в воду, и личинки легко скатываются по нему.
Сетчатая веслоногая лягушка (Rh. reticulata), живущая на Цейлоне, носит свои двадцать яиц прикрепленными к брюху. Яйца эти, величиной с конопляное семя, крепко примыкают одно к другому, образуя собой круглую пластинку. Головастики веслоногой лягушки завершают развитие в воде.
Рис. 66. Костноголовая веслоногая лягушка (Rhacophorus otilophus)
У крестоносной лягушки (Rh. cruciger) на спине, окрашенной в светлый красновато-серый цвет, имеется темный рисунок в форме песочных часов, который начинается в промежутке между глазами и оканчивается несколько кзади от плеч. Этот рисунок может по временам становиться более узким и более резким, местами сходить в конце концов совсем на нет и затем появляться снова. В очень сухую пору эта лягушка имеет цвет белой слоновой кости. С увеличением влажности животное сильно темнеет.
Еще один вид - Rh. dennysi - отличается очень крупными размерами, до 110 мм. По зеленой спине у нее безо всякого порядка разбросаны черно-бурые пятна с узкими желтыми ободками, так что лягушка напоминает лист, пораженный грибной болезнью.
Представители самого большого рода из этого семейства - Hyperolius - обитают в Африке и на Мадагаскаре. Это маленькие лягушки, красивой окраски, ведущие древесный образ жизни. Замечательна пестрая Н. melanoleucus, открытая в 1950 г. в горах Конго, У Н. concolor окраска молодых отлична от взрослых. Спина у них светло-коричневая с многочисленными темно-коричневыми и светло-серыми окаймленными пятнами. Пятью месяцами позже животное становится одноцветным оливково-зеленым, через 16 месяцев - зеленым, как лист, с лимонным брюхом и кораллово-красной внутренней стороной ног. У вида Н. cinctiventris внутренняя сторона бедер и пальцы кроваво-красные, особенно днем. Брюхо розовое или землянично-красное. Самец крупнее самки, однотонно коричневый, имеет точечный, а иногда и сетчатый рисунок на спине.
Лягушки из рода Hylambates населяют Тропическую и Южную Африку. Подобно предыдущим, они по внешнему виду очень напоминают квакш. Н. rufus - лягушки плотного строения, с грубозернистой кожей, с большими выпученными глазами. Самцы у этого вида не крупнее 55 мм, тогда как самки достигают 85 мм длины. Самки встречаются в природе гораздо реже самцов. Самцы иногда серые, иногда серо-бурые с зеленым рисунком и великолепными темно-желтыми суставными сгибами. Самки светло- или темно-серые или красно-коричневые с большим темным ромбом на спине и поперечными полосами на ногах. У самок в яичнике найдены яйца 3 мм в диаметре. Активны ночью. Н. vermiculatus тоньше, меньше, чем предыдущий вид, и имеют более гладкую кожу. Бока молочно-белые с черными пятнами. Скрытые части задних ног бирюзово-голубые. Н. brevirostris отличаются своеобразной заботой о потомстве, встречающейся еще только у рыб. Самка этого вида носит крупные яйца (около 4 мм в диаметре) во рту.
Очень красива банановая лягушка (Меgolixalus laevis) - обитатель Камеруна, 30-40 мм длиной, днем шоколадной окраски, по бокам у нее широкие полосы с металлическим блеском, которые иногда сзади, объединяясь, образуют подкову. Ночью эти животные красно-коричневые с перламутровыми пятнами. Активность ночная. На почву не спускаются. Выделения кожи очень ядовиты. В неволе с наступлением сумерек самцы собираются на листьях, висящих над водой, и выводят свои мелодичные трели. При этом они толкаются, приплясывают вокруг друг друга и ударяют себя по морде задними ногами. Готовые к откладке икры самки выбирают самца, спариваются с ним и, подобно филломедузам, откладывают 40 яиц в свернутый при помощи задних ног лист. В некоторых случаях кладка продолжается на другом листе. В течение лета наблюдается 4-5 кладок. На 12-й день выводятся личинки и падают в воду. Через 2-3 месяца из воды выходят молодые лягушки 18-20 мм длиной и на следующий год становятся половозрелыми. Также на листья откладывает яйца хватающая лягушка (Chiromantis rufescens) из Камеруна.
Рис. 68. Африканская банановая лягушка (Меgolixalus laevis)
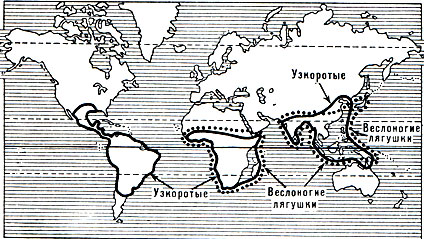
Рис. 69. Области распространения семейств узкоротых и веслоногих лягушек
Семейство Узкоротые (Brevicipitidae)
Объединяет около 175 видов, как древесных, так и наземных и роющих. Узкоротые чрезвычайно разнообразны по внешнему виду и особенностям строения, почему их группируют в 56 родов и 14 подсемейств. Большинство родов включает всего по 1-2 вида.
Самый большой род узкоротых квакш - Microchyla (29 видов). Представители его широко распространены и встречаются в Юго-Восточной Азии, Малайском архипелаге, Южной Америке и на юге Северной Америки.
Каролинка (М. carolinensis) - обитатель Северной Америки, имеет в длину 30-35 мм. Это проворное животное отличается яйцеобразной формой тела, ведет сумеречный образ жизни и нуждается во влажности и почве, обильно поросшей растительностью. День проводит в норах или под гниющими деревьями. Спаривание с конца апреля до начала сентября. Крик самца напоминает голос ламы. Самка, которая несколько меньше самца, откладывает 40 яиц.
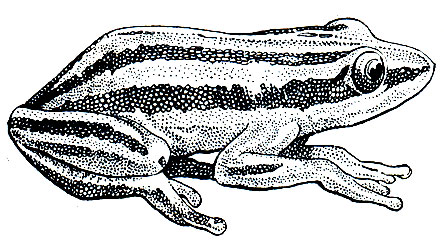
Рис. 70. Конголезская пятилинейная лягушка (Hyperolius quinquevittatus)
В Юго-Восточном Китае широко распространена красивая узкоротая квакша (М. pulchra) длиной в 28-35 мм. Спина у нее красно-коричневая или шоколадная с отчетливым темным треугольным рисунком и темными пятнами. Передняя сторона бедер от желто- до оранжево-красной. Обитает в холмистой местности, поросшей невысокой травой и отдельными кустарниками. Может прыгать на 3 м. Мечут икру с марта до мая в маленьких скоплениях воды. В воде, бедной кислородом, при 23-28° развиваются плоские, почти прозрачные головастики, имеющие явную плавательную перепонку на задних* ногах, тогда как у молодых лягушек ее нет.

Рис. 71. Красивая узкоротая квакша (Microhyla pulchra)
Род Kaloula содержит 11 видов, специализированных на питании муравьями и термитами, обитающих в Южной и Восточной Азии и на Малайском архипелаге.
Маленькая голова украшенной лягушки (К. pulchra), окрашенная в охристо-желтый цвет, представляет своеобразный контраст большому телу, имеющему красивую коричневую окраску. Широкая охристо-желтая полоса тянется и по бокам тела лягушки от века до задних конечностей. Длина тела животного 75 мм. Задние ноги имеют сильно развитый внутренний пяточный бугор, которым животное хорошо роет. Передвигается маленькими торопливыми прыжками. При помощи расширенных концов пальцев может хорошо лазать. В Сингапуре кричат в период размножения так, что близко от них не слышно своего голоса. У кричащего самца раздувается не только резонатор, но и все тело. Испугавшись, животное надувается, и его ширина становится почти равной удвоенной длине. Успокоившись, лягушка выпускает из себя воздух с отчетливо слышным шипением. Червеобразный язык ее может выбрасываться на 4 см. Кормится почти исключительно муравьями.
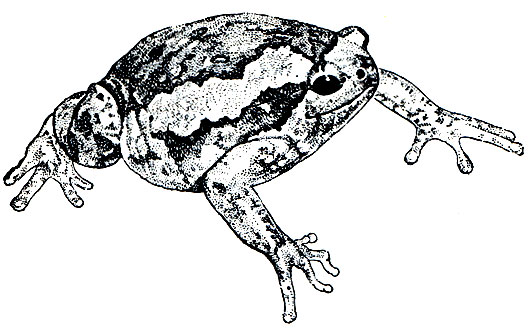
Рис. 72. Украшенная лягушка (Kaloula pulchra)
Крупная К. baleata похожа на предыдущий вид образом жизни и окраской. Однако она часто имеет кирпично-красные пятна на плечах и в основании задних конечностей. Хорошо закапывается, встречается и под камнями и на деревьях. Основная пища - термиты.
Лягушки рода Stereocyclops, живущие в Южной Америке, имеют своеобразное приспособление к питанию термитами. Тонкая кожа спины этого животного густо усеяна громадными железами, выделяющими жидкость, которая затвердевает на воздухе, образуя своего рода панцирь - средство защиты от термитов. В дополнение костное кольцо окружает роговую оболочку глаза.
В Африке живут представители рода узкоротых (Breviceps). Всего их известно 6 видов. Эти земноводные в спокойном состоянии похожи на резиновый мяч.

Рис. 73. Капский узкорот (Breviceps gibbosus)
Восточно африканский узкорот (B. mosambicus) отличается чрезвычайно плотным сложением, короткой головой, почти плоской мордой, конец которой едва выдается, маленьким ртом и направленными вперед глазами. Конечности его очень коротки и до локтевого и коленного суставов скрыты в коже туловища; тем не менее они сильно развиты. На пятке находится большая лопатообразная мозоль с заостренным краем, служащая для рытья. На спине он имеет грязную красно- бурую окраску, на боках - желто-бурую. Может быть одноцветным или покрытым различного рода черными пятнами. Всегда имеется косая черная полоска, которая тянется от глаза вниз и назад. Нижняя сторона грязно-белая с большим черным пятном на горле. Длина тела 50 мм. В больших количествах появляется после дождя. Неблагоприятное время года проводит зарывшись. Вероятно, питается термитами.
В Южной и Тропической Африке живут представители рода лягушек-поросят (Hemisus). Мраморная лягушка (Н. marmoratus) - небольшая лягушка, которая может надуваться так же, как и короткоголов, сверху серо-бурая с темным мраморным рисунком; горло, бока и задняя сторона бедра зеленоватожелтые; низ белый. Это животное интересно в том отношении, что самка, зарывшись в землю, прикрывает собственным телом яйца, из которых детеныши выходят в виде хорошо развитых головастиков.

Рис. 74. Мраморная лягушка (Hemisus marmoratus) при виде опасности сильно раздувается
Очень редкая лесная лягушка из Южной Африки малый узкорот (Anhydrophryne rattrayi) откладывает яйца в специально вырытую ямку. Яйца почти белые, заключены в желатиновую капсулу. В кладке 11-19 яиц. Через 11 дней из них появляются головастики, остающиеся сидеть все вместе. Через 15 дней после вылупления у них исчезает хвост и лягушата выходят наружу.
У лягушки Mantophryne robusta, из Новой Гвинеи, немногочисленные крупные яйца соединены четковидно. Самец прикрывает их своим телом и охватывает передними лапами. Все развитие проходит в яйце, причем органом дыхания головастика служит хвост.
Самки восточноафриканских узкоротых жаб рода Hoplophryne кладут яйца между листьями бананов или в старые бамбуковые стволы. Представитель другого рода африканских узкоротых жаб - Phrynomantis microps отличается наиболее пестрой окраской тела. Вся верхняя сторона у нее яркого кирпичного цвета с золотисто-зеленым отблеском посередине спины; бока головы и туловища резко выделяются густым черным цветом, и черная же окраска внедряется в окраску спины в виде остроугольного, направленного от бедер вперед треугольника. Брюшная сторона синевато-серая с различными мелкими светлыми пятнами. Подобные же светлые пятна, только большие по размерам и более редко расположенные, находятся на верхней стороне конечностей. Все животное достигает размеров средней величины квакши. Лягушка лишена способности прыгать, ползает она довольно медленно, но производит впечатление гибкого животного. Она умеет очень ловко пользоваться самыми ничтожными углублениями, чтобы спрятаться, и узкими щелями и расселинами для прохода. Питается термитами.
Другие роды из семейства настоящих лягушек немногочисленны. Большинство из них объединяют 1-5 видов.
Род Arthroleptis представлен 35 видами, обитающими в Южной и Тропической Африке. Они характеризуются отсутствием нёбных зубов, горизонтальным зрачком, сердцевидным языком, малоразвитыми плавательными перепонками и крепко связанными между собой наружными костями плюсны. Все они принадлежат к наземным или даже роющим формам.
Длиннопалая лягушка (Arthroleptis stenodactylus) весь день проводит зарывшись и появляется на поверхности земли лишь ночью. У самца этого вида, как и у других родственных видов, третий палец передних конечностей сильно удлинен. Длина этого маленького животного не превышает 40 мм.
Род Phrynobatrachus включает 14 видов крошечных водяных лягушек, живущих в Африке.
Наталийская лягушка (Phrynobatrachus natalensis), представляющая чрезвычайную изменчивость в цвете и рисунке и редко имеющая более 30 мм в длину, распространена от Судана до Южной Африки и Анголы и местами очень многочисленна. В Африке же обитает в высшей степени своеобразная волосатая лягушка (Astylosternus robustus). В брачное время у самцов этой лягушки на боках тела и бедрах развиваются длинные волосовидные сосочки кожи, служащие дополнительными органами дыхания. Появление их связано со слабым развитием легких.
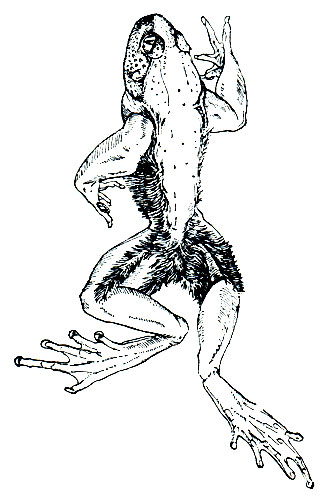
Рис. 75. Волосатая лягушка (AstylosternuS robustus)
Весьма причудливую форму тела имеет рогатая лягушка (Ceratobatrachus guen- theri) - обычный обитатель Соломоновых островов. Ее треугольная приплюснутая голова очень невелика и вытянута на морде в остроконечный кожный вырост. По одному такому же острию имеется на верхних веках, над заднепроходным отверстием и пяточным сочленением. Поперек головы и вдоль спины проходят более или менее многочисленные тонкие кожные складки. Узкая, часто зубчатая кожная лопасть окаймляет наружную сторону предплечья и плюсны. Вся нижняя сторона тела в мелких зернистых бородавочках. Окраска и рисунок на теле лягушки очень изменчивы и имеют приспособительное значение, помогая ей сливаться с окружающим фоном. Размеры тела 75-85 мм. Развитие лягушки проходит в яйце.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'