Класс эхиуриды (Echiuroidea)
К классу эхиуровых червей относится не менее 100 видов свободноживущих типично морских раздельнополых червей, стоящих близко к морским кольчецам-полихетам, хотя по многим признакам они резко от них отличаются.
Ряд других признаков дает полное основание сближать их с кольчецами и включать в тип аннелид в качестве четвертого класса.
Размеры тела эхиурид колеблются от нескольких миллиметров до 20-30 см. Геологически это очень древняя группа роющихся в грунте червей, получивших в связи с таким образом жизни ряд характерных особенностей.
Мешковидновальковатое тело эхиуриды (рис. 304) спрятано в грунте. На переднем конце его, за редким исключением, помещается хобот, способный сильно вытягиваться и сжиматься. В нормальном положении хобот поднимается над грунтом и у некоторых форм имеет в длину свыше 1 м. Других придатков тела у эхиурид, кроме щетинок, нет. У отверстия нефридиев у части эхиурид находятся две так называемые половые щетинки, а у некоторых эхиурид вокруг анального отверстия располагается кольцо щетинок в один или два ряда.
У взрослых форм совершенно отсутствуют сегментация, параподии и параподиальные щетинки. Кожа тонкая и прозрачная, особенно у форм, живущих на глубинах; у мелководных видов - иногда толстая и покрытая папиллами.
Кишечник у эхиурид сквозной, часто образующий петли (рис. 304); ротовое и анальное отверстия расположены на противоположных концах тела, ротовое - у основания хобота. Хобот обычно имеет желобчатый характер и покрыт мерцанием. Передний конец хобота иногда расширяется в большую лопасть, у некоторых видов цельную, у других - разделенную на два лепестка. Внутри тела эхиурид помещается обширная полость. Сам червь, особенно взрослый, почти неподвижен, хобот же его обладает большой подвижностью. Внутри его и вдоль тянутся три канала, сообщающихся с общей полостью тела. Сокращение мышц кожно-мускульного мешка создает внутренний тургор: под некоторым давлением полостная жидкость нагнетается в каналы и полости хобота, и он приобретает твердость (гидравлический скелет). При ослаблении внутреннего давления хобот спадается и избегает таким образом опасности быть съеденным другими животными. По переднему краю хобота иногда располагаются особые органы чувств, возможно своеобразные "радары", предупреждающие об опасности. В случае утери хобота он довольно быстро регенерирует вновь. Полость тела обширная и несегментированная. Единственный след сегментированности предков у взрослых форм - это наличие у некоторых представителей эхиурид 2-3-4 пар нефридиев. Нефридии эхиурид очень сходны с нефридиями сипункулид, но в еще большей степени теряют железистые функции и служат для выведения наружу зрелых половых продуктов. Гроздевидные скопления половых продуктов развиваются внутри полости тела, и яйца по достижении половозрелости захватываются нефростомом, а иногда полностью набивают нефридий.
Наружное отверстие нефридиев (нефропор) снабжено двумя крепкими половыми щетинками, помогающими при оплодотворении (рис. 304,5). Для части эхиурид (сем. Веnelliidae) известен резко выраженный половой диморфизм с развитием самцового паразитизма. Микроскопические самцы паразитируют в особых отделах нефридиев самок, где и осуществляют оплодотворение яиц. Они уже настоящие паразиты, лишенные пищеварительной системы и ряда других органов. Экспериментально было показано, что из большей части яиц, развивающихся на хоботе самки, выходят самцы, которые спускаются по хоботу к телу самки и проникают затем через половое отверстие, располагающееся вблизи хобота на брюшной стороне тела, внутрь нефридиев.
Из яиц, развивающихся вне контакта с самками, выходят главным образом самки.
У эхиурид хорошо развита кровеносная система. В самый задний отдел кишки открываются две анальные железы, выполняющие выделительные функции. Они состоят из одного или нескольких иногда ветвистых канальцев, обычно покрытых большим количеством железистых папилл с воронками на концах.
Эхиуриды - типичные детритоеды, их кишечник обычно бывает набит большим количеством детрита.
Очень типично для эхиурид спиральное дробление и личиночные формы, не оставляющие сомнений в близости эхиурид к полихетам. Это весьма типичные трохофоры с хорошо выраженной зачаточной сегментацией, которая в дальнейшем исчезает. Значительное своеобразие строения, не сходного с какой-либо другой группой кольчатых червей, малое число видов эхиурид, приуроченность большого количества видов к абиссальным глубинам океана могут быть истолкованы как доказательства большой их древности.
В северных морях обитает всего 3-4 вида эхиурид. Главное разнообразие их приурочено к тропическим водам. Около трех десятков видов характерно для больших глубин океана, где иногда они являются характерной группой населения.
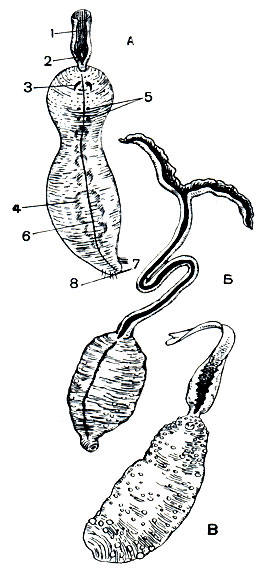
Рис. 304. Представители класса эхиурид. А - Echiurus echiurus: 1 - хобот; 2 - придаток хобота; 3 - брюшные щетинки; 4 - петли кишечника, просвечивающие сквозь стенку тела; 5 - половые отверстия; 6 - брюшная медиальная линия; 7 - анальное отверстие; 8 - анальные щетинки; Б - Bonellia viridis; В - Thalassema gigas
Класс сипункулиды (Sipunculoidea) (добавочный класс к высшим червям)
Сипункулиды - сравнительно небольшая группа (около 300 видов) типично морских червей. По ряду признаков их можно отнести к высшим червям как дополнение к типу аннелид.
Сипункулиды в процессе эволюционного развития далеко отошли от основного ствола кольчатых червей. Тело сипункулид гладкое, лишенное совсем каких-нибудь придатков, в том числе параподий и щетинок. Оно вытянуто и снабжено хоботом, который может выбрасываться далеко вперед, удлиняя ход в грунте. На конце хобота находится ротовое отверстие, окруженное венцом щупалец. Шупальца служат для сортировки частиц, годных в пищу. Анальное отверстие находится в передней трети тела. Кишечник значительно длиннее тела, и его нисходящая и восходящая части закручены винтом одна вокруг другой. Тело сипункулид погружено в грунт или в пористую массу известковых водорослей или кораллов, а хобот выставлен наружу. При таком положении тела анальное отверстие оказывается над поверхностью грунта и фекалии выделяются в окружающую воду.
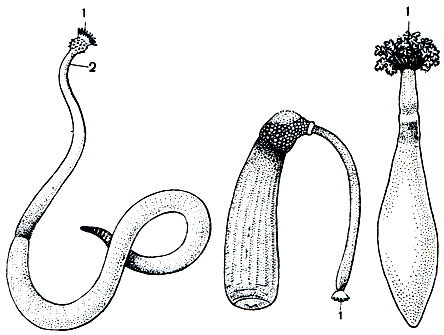
Рис. 305. Представители класса сипункулид: Слева направо: Golfingia vulgare; Aspidosiphon steenstrupii; Dendrostomum pyroides. 1 - венчик щупалец вокруг ротового отверстия; 2 - хоботок
В передней части тела помещаются один, два или несколько пар нефридиев, имеющих железистую стенку и сложенный вдвое канал. Нефростомами нефридии открываются в полость тела, а нефропорами - наружу.
Нервная система сипункулид развита слабо, что соответствует их малоподвижному и скрытому существованию. От окологлоточного нервного кольца по брюшной стороне тела отходит нервный тяж. Органы чувств отсутствуют.
Полость тела поделена на три части. Однако перегородки выражены слабо. Полостная жидкость перегоняется мускулатурой, и сипункулиды движутся гидравлическим способом.
Сипункулиды раздельнополы. Половые продукты формируются в полости тела и по созревании выводятся наружу через нефридии. Оплодотворение яиц происходит в воде.
Спиральное дробление яиц и наличие личинок типа трохофоры сближает сипункулид с полихетами. У некоторых сипункулид на стадии поздней трохофоры развиваются щетинки, которые потом исчезают. Таким образом обнаруживается связь этих животных с кольчатыми червями, особенно на ранних стадиях развития. Но по мере развития признаки сходства все более утрачиваются. Поэтому в последнее время все большее число исследователей склонно выделять сипункулид в самостоятельную группу как добавление к высшим червям. В щупальцевом аппарате и в передней части тела сипункулид имеется особая система каналов, несколько напоминающая амбулакральную систему иглокожих. Иногда их считали родственными мшанкам и форонидам. Но и до настоящего времени систематическое положение сипункулид остается не ясным.
Сипункулиды обитают во всех морях и океанах (вплоть до больших глубин), образуя значительную и характерную часть донного населения.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'