23. Видообразование
Ключевые вопросы
Что такое изменчивость видов?
Как происходит образование видов в результате географической изоляции?
Как происходит образование видов в результате полиплоидии?
Существует ли равновесие между образованием и вымиранием видов?
Почему существует такое большое разнообразие живых организмов?
За время, прошедшее с тех пор, когда жил Дарвин, эволюционисты узнали, как происходят мутации, каким образом гетерозис поддерживает мутационную изменчивость и как различаются процессы адаптации в зависимости от окружающей среды.
Но до сих пор мы не имеем удовлетворительного объяснения существующего разнообразия форм жизни. Во первых, необходимо понять, как образуются новые виды и какие в этом направлении достигнуты успехи.
Однако любопытно, что, хотя основная работа Дарвина называется "Происхождение видов", успехи в этом принадлежат не ему.
То, что он в действительности описал и объяснил, - это изменчивость видов, а не процесс, в результате которого из одного вида образуются два различных.
Во-вторых, необходимо выяснить, как может выжить такое множество видов.
Здесь, как вы увидите, наши знания далеки от совершенства.
23.1. Вид, распространенный на большой территории, может выглядеть совершенно различным в разных условиях окружающей среды
Обычный злак, например ячмень, проявляет экогеографическую изменчивость, т. е. изменение внешнего вида в зависимости от характера окружающей среды, в которой он растет. Во влажном климате ячмень часто имеет жесткий стебель, который предотвращает полегание растения, но на ветреных сухих равнинах, например, западной части США, более приспособленным оказывается ячмень с гибким стеблем, потому что зерно с жесткого стебля может сдуть ветер.
Экогеографическая изменчивость возникает потому, что отдельные фенотипы неодинаково приспособлены к разным условиям окружающей среды.
Когда вид изменяется в зависимости от условий местообитания, это обычно происходит почти незаметно. Так, например, ящерицы в одной долине почти не отличаются от ящериц, обитающих в соседней долине. А ящерицы этой второй долины почти такие же, как в третьей долине, и т. д. Но если мы придем на тридцатую долину и сравним ее ящериц с ящерицами первой, то обнаружим заметные различия между ними. Такое постепенное изменение внешних признаков вида при переходе от одной части ареала к другой называется клином. Это довольно распространенное явление. Более того, у одного вида такое изменение может охватить сразу несколько признаков: размер тела может увеличиваться на более высоких широтах, черный цвет может стать более интенсивным по мере увеличения влажности, а коричневый - по мере увеличения температуры и т.д. Таким образом, вид изменяется почти непрерывно. По одному признаку его особи в зоне А могут быть сходными с особями этого вида в зоне Б. Но по какому-то другому признаку те особи, которые находятся в зоне А, сходны с особями, которые находятся в зоне В.
Требуются обширные исследования, чтобы описать такое многообразие форм, а по мере совершенствования научных описаний и. открытия новых промежуточных организмов все труднее и труднее становится определение границ между видами.
Частично решение этой проблемы заключалось в классификации организмов на подвиды, т. е. в том, чтобы дать им третье название дополнительно к роду и виду. Например, длиннохвостый горностай, живущий на северо-востоке США, не имеет на голове выделяющихся отметин. Там его называют Mustela frenata noveboracensis. В Техасе особи этого же вида имеют отметину в виде белой "уздечки", которая помогает им маскироваться, и их называют Mustela frenata frenata (рис. 23-1). Но дело в том, что многие длиннохвостые горностаи имеют только часть - "уздечки". Практически же можно встретить животных, отражающих все промежуточные ступени перехода от Mustela frenata noveboracensis к Mustela frenata frenata. Где же тогда должна пролегать граница между этими двумя подвидами?
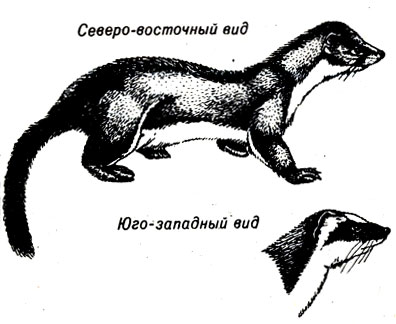
Рис. 23-1. Длиннохвостые горностаи с 'уздечкой' на голове и без нее. (Из: Field Guide to the Mammals, Burt and Grossenheider, Houghton - Mifflin.)
Ответ таков: она должна быть проложена на карте более или менее произвольно. Чаще всего такого места, где можно было бы точнр прочертить линию границы, не бывает вообще.
У людей для обозначения подвидовых групп употребляют термин раса.
23.2. Все биологи признают, что люди образуют один общий вид
Людой принято подразделять на четыре расы: монголоидную (включая американских индейцев), негроидную, кавказскую и австралоидную (черные жители Австралии, которых иногда называют аборигенами). Однако человеческие расы сливаются друг с другом так же, как подвиды американских горностаев, и таксономисты могли бы отстаивать существование 60 рас в равной степени, как и четырех. В учебнике для средней школы выделяют три расы: монголоидную, негроидную и европеоидную.
Хотя термин "расы" в значительной степени несовершенен, кто будет отрицать, что человеческий генофонд проявляет географическую изменчивость? Швед обычно имеет более высокий рост, чем итальянец; у жителя Пекина более заметна складка верхнего века, чем, например, у индейцев штата Аризона, а норвежцы имеют более светлую кожу, чем жители Конго.
Многие человеческие признаки проявляют географическую изменчивость, но, поскольку цвет кожи является наиболее отличительным признаком, мы используем его в качестве примера. Подвергся ли цвет кожи эволюции? Если да, то когда и почему?
23.3. При адаптации к окружающей среде общая окраска многих видов животных, а также цвет кожи у человека проявляют географическую изменчивость
Изменчивость цвета кожи у человека и внешней окраски животных осуществляется в основном за счет одного и того же пигмента - меланина. Эта изменчивость часто бывает параллельной у многих родственных видов.
Иначе говоря, если один вид, живущий в определенном регионе, является темным, то другие виды этого же региона тоже, вероятно, будут темными.
Вильям Хованиц (William Hovanitz) провел тщательное изучение пигментов в крыльях бабочек, обитающих в Калифорнии.
Он обнаружил параллельную эволюцию окраски крыльев у большинства из них.
Меланиновые пигменты обычно более интенсивны в прохладной влажной среде.
Хованиц распределил бабочек-шашечниц (Melitaea) соответственно интенсивности их окраски (рис. 23-2). Наиболее интенсивно окрашенные формы бабочек находятся в верхней части каждой колонки. Все эти формы обитают в основном в прохладных влажных северных областях Калифорнии. Наиболее светлые формы - в жаркой сухой пустыне Мояве (Mojave Desert). Клинья меланиновой интенсивности соединяют оба эти региона.

Рис. 23-2. Формы четырех видов рода Melitaea, обитающие в Калифорнии. Бабочки в каждой колонке принадлежат к виду, название которого указано под колонкой. Черный, а также красно-коричневый и желтый цвета наиболее интенсивны у бабочек, расположенных в верхней части колонок, и заметно слабеют по мере понижения. (Из: And Replenish the Earth, Rosenzweig)
Хованиц получил аналогичные результаты при изучении рода Argynnis.
Очень часто исследователям удается найти связь между фенотипическими и экологическими клиньями. Такие связи нашли отражение в экогеографических правилах.
Например, правило Аллена (Allen) устанавливает, что выступающие части тела у птиц и млекопитающих обычно бывают короче в более холодном климате.
Правило Бергманна (Bergmann) заключается в том, что птицы и млекопитающие, обитающие в более высоких широтах, бывают крупнее особей своего вида, живущих ближе к экватору.
Правило Рапопорта (Rapoport) указывает, что различные виды ногохвосток (отряд Collembola; примитивные бескрылые насекомые, которые чаще всего обитают в почве) имеют более темную окраску в условиях холодного климата.
Поскольку экбгеографическая изменчивость проявляется параллельно у многих видов, можно полагать, что она является адаптивной. Наиболее убедительным доказательством этого служат виды, производящие два потомства в год, каждое из которых отличается по внешнему виду, очевидно, в зависимости от необходимости приспосабливаться к различным сезонам. Например, бабочка Pieris napi, имеющая горчично-белую окраску, производит более темное потомство весной, в относительно холодный в Калифорнии сезон, следующий за периодом дождей, и более светлое потомство летом, когда погода жаркая и сухая (рис. 23-3). Ясно, что этот вид адаптировался к изменяющимся условиям среды. Более темные бабочки лучше приспособлены к холодному сырому климату, чем светлые. А светлые бабочки более приспособлены к теплому сухому климату.
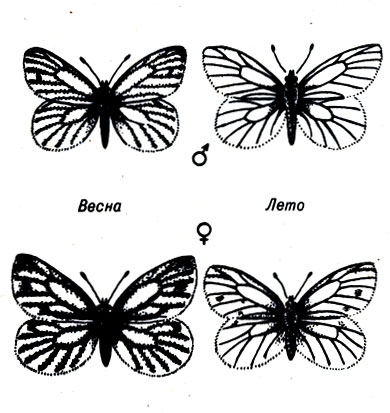
Рис. 23-3. Взрослые особи Pieris napi в разные сезоны, но из одного региона (Центральная Калифорния). Более темная форма - весенняя, более светлая - летняя. (Из: And Replenish the Earth, Rosenzweig.)
Однако мы не знаем причины изменения приспособленности. Для выяснения этого необходимы эксперименты, подобные тем, которые произвел Кеттлвелл (Kettle well) на пятнистых бабочках Biston betularia.
23.4. Правило Глогера, которое устанавливает связь окраски птиц с четырьмя основными климатическими условиями, по-видимому, распространяется и на млекопитающих
К. Л. Глогер (С. L. Gloger), немецкий биолог, живший в XIX в., предложил первое экогеографическое правило в 1833 г. Оно указывало, что популяции птиц в теплом влажном климате состоят из особей с черной окраской, в теплом сухом климате - из особей с коричневой окраской, в холодном сухом климате - из очень бледных особей и в холодном влажном климате - из менее черных, но все же темных форм.
| Среда | Холодная | Теплая |
| Влажная | Менее черные | Очень очерные |
| Сухая | Бледные | Коричневые |
С тех пор было установлено, что правило Глогера распространяется не только на птиц, но и на млекопитающих. Оно относится и к людям. Правило Глогера действовало у 80% исследованных видов птиц и млекопитающих.
Однако бабочки только частично подчиняются этому правилу, а некоторые насекомые совсем не следуют ему.
Хотя мы не знаем, почему изменения, описанные в данном правиле, увеличивают приспособляемость, интересно то, что они все же повышают ее. По крайней мере они увеличивали приспособляемость в предшествующие времена. Однако сейчас, когда люди так сильно изменили окружающую среду, географические, расовые модели прошлого не могут, по-видимому, являться оптимальными для будущего.
23.5. Первые люди могли быть черными, коричневыми или желтыми, но не белыми
Широкое распространение Homo erectus дает основание предполагать, что различия в цвете кожи существовали до того, как сформировался человеческий вид. Поскольку наши предки были млекопитающими и поскольку млекопитающие обычно подчиняются правилу Глогера, очень возможно, что наши предки тоже подчинялись этому правилу. Если они жили в хододном северном климате, они могли быть белыми. А те, которые населяли тропические леса, где выпадали большие осадки, были, по-видимому, черными.
Воздействия, которые трансформировали таз, челюсти и мозг человека и превратили его, таким образом, в Homo sapiens, не были связаны с цветом кожи. Возможно, один из таких "очеловечивающих" аллелей возник в Малайзии, другой - в Пакистане, еще один - в Гвинее и четвертый - в Китае. Никто точно не знает, сколько было аллелей, но вряд ли все они возникли в одной популяции и в одном месте. Затем каждый новый аллель постепенно распространялся благодаря естественному отбору в огромном генофонде популяции гоминид по всему свету. Но зачем должно изменяться то, что уже хорошо адаптировалось? Зачем должен изменяться цвет кожи?
Некоторые расовые разновидности, возможно, появились не так давно. Для того чтобы успешно существовать в холодном климате, человеку, по-видимому, необходима была большая часть культурного багажа каменного века: хорошие орудия, огонь и знание того, как одеваться и строить для себя жилища. Возможно, что предки человека жили и в холодных северных районах, но мы точно знаем, что они населяли тропики. Вполне вероятно, что "черная раса" берет свое начало со времени происхождения человечества и что только после того, как люди сделали прогресс в сторону цивилизации, они смогли мигрировать и заселить более холодную Европу, оказавшись в климате, который вызвал эволюцию прозрачного эпидермиса.
До сих пор мы Говорили об изменении видов, или филетической эволюции, но не рассматривали вопрос об истинном образовании видов, при котором из одного вида офразуютря два или более. Было предложено несколько совершенно различных механизмов видообразования, по крайней мере два из них являются особо важными.
23.6. Виды характеризуются как по их внешним признакам, так и по поведению при спаривании
Мы с. вами перелистали много страниц, не дав до сих пор определения вида, но сейчас, переходя к обсуждению происхождения видов, нам необходимо будет знать точно, о чем мы говорим. Существует двойное определение вида.
1. Фенотипический вид - самая старая концепция вида. Это популяция организмов, в которой особи двух наиболее различающихся фенотипов больше сходны друг с другом, чем с любой особью любого другого вида. Иначе говоря, это совокупность сходных фенотипов.
2. Биологический вид - это совокупность организмов, скрещивающихся между собой в природе. Эта концепция вида имеет много преимуществ. Поскольку таксономисты практически могут наблюдать скрещивание в естественных условиях, это позволяет им объективно классифицировать организмы на виды. Особи одного биологического эида являются эволюционной единицей. Таким образом, биологический вид является естественной классификационной единицей.
Большинство (если не все) другие единицы классификации, такие, как типы и классы, до некоторой степени произвольны. Например, птицы и крокодилы в своем происхождении связаны довольно близко, и в действительности крокодилы даже более близки к птицам, нежели к черепахам. Предки черепах отделились от общих предков крокодилов и птиц около 280 млн. лет назад, а дивергенция предков птиц и крокодилов произошла примерно 230 млн. лет назад. Однако биологи считают, что намного удобнее объединить крокодилов и черепах в класс пресмыкающихся (вместе с ящерицами, змеями и гаттериями), а птиц выделить в отдельный класс.
В отношении биологических видов такая произвольность недопустима. В природе каждая особь может скрещиваться с другой особью или нет. Если в результате естественного* скрещивания гены организма А могут стать частью генов потомков организма Б (и наоборот), тогда А и Б являются особями одного биологического вида. Однако, поскольку у некоторых организмов половое размножение отсутствует или по крайней мере является очень редким, мы не можем отвергнуть концепцию феноти-пического вида.
Остановим наше внимание на биологическом виде. Каким же образом размножающаяся половым путем популяция приводит к возникновению другой популяции, скрещивание с которой становится невозможным? Имеются по крайней мере два ответа на этот вопрос: в результате географической изоляции и полиплоидии.
23.7. Имеются данные о том, что географическая изоляция является основным механизмом процесса видообразования
Мы уже знаем, что организмы стараются приспособиться к условиям конкретной окружающей среды. Поэтому можно, наверное, предположить, что если бы существовала какая-то географическая преграда, разделяющая две группы организмов одного биологического вида, то эволюция каждой группы пошла бы в более или менее различном направлении. Представьте популяцию североамериканских белок по обеим стрройам Большого каньона в США. На каждой стороне животные живут в сосновом лесу, но они разделены экологически непригодным пустынным руслом реки Колорадо. Высота каньона с северной стороны примерно на 30,5 м больше чем с южной стороны, поэтому воздействие окружающей среды, испытываемое обеими популяциями, должно несколько отличаться.
Популяции белок на обеих сторонах каньона действительно различаются между собой. Белки, живущие на северной стороне, имеют белый хвост и черное брюшко; белки южного берега обычно имеют серо-белое брюшко и хвост, темный с дорсальной стороны. Иногда случается увидеть южную белку в северной популяции и наоборот; таким образом, нам ясно, что отбор поддерживает существование разных фенотипов на обеих сторонах каньона, хотя мы и не знаем почему.
Если бы эволюция этих популяций продолжалась, предположим, еще его или двести тысяч лет, то между ними могли возникнуть значительные различия по мере адаптации каждой популяции к испытываемому ею давлению. Это может выразиться в изменении числа хромосом в одной популяции или, возможно, в появлении новой ферментной системы в другой.
Во всяком случае, существует вероятность, что, если мы обеспечим этим популяциям возможность скрещиваться (заполнив Большой каньон), они не смогут этого сделать или же их гибридные потомки (так же как мул, потомок осла и кобылы) будут стерильны и нежизнеспособны. Это будет означать, что произошла эволюция двух видов из одного. А иначе вопрос о том, представляют ли они один или дьа вида, будет оставаться спорным -и не будет иметь биологического значения.
Последовательность географического видообразования предстает в следующем виде:
1. Один биологический вид разделяется на две или более изолированные популяции какой-то географической преградой.
2. Каждая популяция эволюционирует самостоятельно в своей среде обитания.
3. Воссоединение популяций создает возможность для возобновления скрещивания, но популяции не участвуют или не могут участвовать в скрещивании. Они становятся самостоятельными биологическими видами.
Имеется много фактов, дающих основание предполагать, что такая последовательность является основным механизмом видообразования.
23.8. Самые убедительные данные о географическом видообразовании получены при исследовании архипелагов
Когда Дарвин посетил Галапагосские острова во время путешествия на "Бигле", его удивили необычайные формы птиц,, которых он там наблюдал. Хотя общее количество видов птиц было менее десяти, зато один из них, Geospizidae (Дарвинов вьюрок), насчитывал более десяти разновидностей. Трудно было поверить, что все эти разновидности были занесены через океан более чем за 1000 км, отделяющих архипелаг от Южной Америки, и Дарвин предположил, что все они произошли от одного вида.
Дарвин считал, что первые вьюрки дали начало популяции, которая постепенно распространилась на пяти больших островах архипелага, а потом и на маленьких удаленных островах. Возможно, что в маленьких, частично изолированных популяциях на удаленных островах частота генов значительно изменилась в результате дрейфа генов. В конечном счете эти популяции стали настолько отличаться от популяций крупных островов, что скрещивание между ними стало невозможным. Поэтому, когда особи популяций удаленных островов возвращались на основные острова архипелага, они образовывали ядро новых популяций. Им приходилось вести конкуренцию с существующими вьюрками за пищу, места гнездования и т. д., и это привело к еще большей дивергенции между видами. Новые популяции стали питаться не только одним нектаром, но и насекомыми, семенами, ягодами и т. д. Потомки этих птиц в конце концов попали на более удаленные острова, и цикл начался заново, т. е. в результате дрейфа генов и естественного отбора в условиях изоляции начиналось образование нового вида. Такой процесс видообразования и дивергенции называется адаптивной радиацией, и на Галапагосских островах вьюрки приспособились использовать такое множество экологических ниш, что образовался один вид вьюрка, который по своему поведению напоминает дятла. Особи этого вида питаются насекомыми, обитающими в коре и под корой деревьев, но, не имея такого длинного клюва, как у дятла, они отламывают колючки или веточки кактуса и извлекают с их помощью насекомых.
Более чем в 1000 км от Галапагосского архипелага находится Кокосовый остров, один из наиболее изолированных участков суши. Здесь мы находим только один вид Дарвинова вьюрка.
Кокосовый остров по размеру равен малым Галапагосским островам. Он представляет собой тропический лесной остров, а в тропических лесах обычно водится больше видов птиц, чем в любой другой естественной среде. Поэтому можно было бы ожидать, что на Кокосовом острове обитает несколько видов птиц. Но на самом деле там только один. Почему? Очевидно, для возникновения изолятов вьюрка на этом крохотном острове недостаточно места. На нем нет географических барьеров, которые бы способствовали видообразованию. Поэтому, хотя кажется, что Кокосовый остров имеет благоприятные условия для образования более чем одного вида Дарвинова вьюрка, вряд ли это произойдет. Получено много убедительных данных, показывающих, что географические барьеры могут быть чрезвычайно важны в качестве первой ступени видообразования для некоторых видов животных.
23.9. Наличие одного или нескольких полных дополнительных хромосомных наборов называется полиплоидией; в плодовитом (фертильном) потомстве полиплоидия является механизмом быстрого видообразования
Обычно потомство, наследующее дополнительную хромосому или полный дополнительный набор хромосом (полиплоидия), нежизнеспособно или стерильно. Это больше относится к животным, чем к растениям. Если полиплоид фертильный, значит, это новый вид, потому что он не может скрещиваться с родительским видом, имеющим меньшее число хромосом.
Полиплоидный организм может иметь двойные, тройные или четырехкратные наборы хромосом родительского вида (рис. 23-4) или может наследовать по одному набору от двух разных видов (рис. 23-5). Первый случай называется аутополиплоидией, второй - аллополиплоидией.
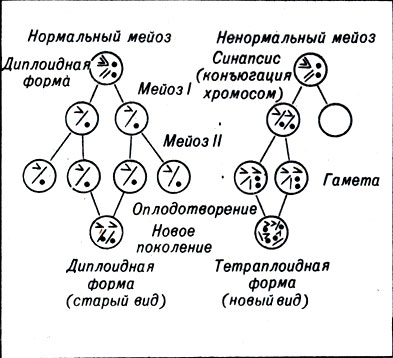
Рис. 23-4. Видообразование в результате аутополиплоидии
Условием полиплоидии являются изменения в процессе мейоза. Гаметы, которые должны сформировать полиплоид, образуются тогда, когда в силу случайности все хромосомы сдвигаются в сторону одной из дочерних клеток. Другая дочерняя клетка вообще не содержит хромосом. Дочерняя клетка с хромосомами образует две диплоидные (а не гаплоидные) гаметы путем обычного клеточного деления.
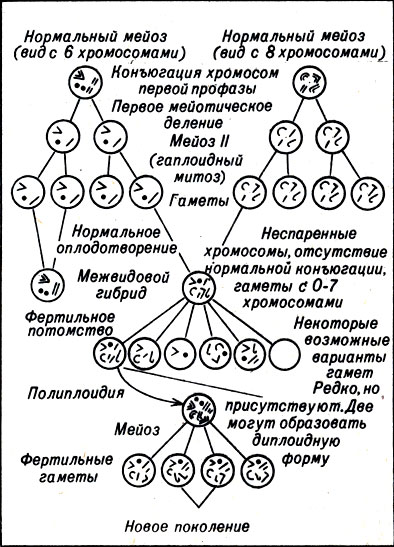
Рис. 23-5 Видообразование в результате аллополиплоидии
Иногда аутопоЛйплоиды получаются от слияния диплоидной и обычной гаплоидной гаметы. В этом случае они являются трип лендами, но они стерильны, потому что их третий набор хромосом не имеет пары в первом мейозе, поэтому триплоиды не могут образовывать нормальные гаметы. Например, бананы являкЭтся триплоидами и размножаются только вегетативным путем, поскольку они не дают жизнеспособных семян.
Если новый полиплоидный вид возник в результате гибридизации двух разных видов, мы называем его аллополиплоидом (от словаа allo - разный). Аллополиплоидия у растений часто приводит к появлению необычно крупного цветка или плода и поэтому используется человеком. Она возникает несколько иначе, чем аутополиплоидия. Первый этап - это гибридизация двух несходных, но нормальных гаплоидных гамет (рис. 23-5). Продуктом этого является гаплоидная зигота, часть хромосомного набора которой наследуется от одного родителя, другая часть - от другого.
Такой гибрид практически стерилен, потому что ни одна из его хромосом не может нормально спариваться во время мейоза. Каждая из его хромосом притягивается к одной из дочерних клеток отдельно от других.
Иногда, совершенно случайно, все хромосомы сдвигаются только к одной ^дочерней клетке. Образуется функциональная гаплоидная гамета с тем же числом хромосом, что и у взрослого гибрида. Если две такие гаметы сливаются, возникает диплоидная зигота. Когда такой гибрид созревает, в нем может происходить нормальный мейоз, так как каждая из его хромосом имеет себе пару. Таким образом, он способен размножаться и к тому же жизнеспособен, это, следовательно, новый гибридный биологический вид.
23.10. Полиплоидия привела к образованию новых видов как в естественных, так и в лабораторных условиях
Очевидно, многие виды растений возникли в результате полиплоидии. Г. Л. Стеббинс (G. L. Stebbins), известный эволюционист-ботаник, установил, что примерно четверть или даже треть растительных видов в мире образовались в результате полиплоидии. Полиплоидия свойственна и некоторым животным.
Чтобы обнаружить полиплоидию в природе, ученые исследовали хромосомы организмов, отыскивая случаи удвоения и утроения числа хромосом. Например, у различных видов пшеницы число хромосом равно 14, 28 или 42. Вид пшеницы с 28 хромосомами мог быть аутополиплои-дом или аллополиплоидом от скрещивания двух видов, имеющих по 14 хромосом, а вид с 42 хромосомами мог быть аллополиплоидным гибридом двух видов, содержащих 28 и 14 хромосом соответственно. Дикие виды хлопчатника имеют 26 хромосом, культурные-52. У плоского червя Dendrocoelum lacteum 16 хромосом, а родственный ему вид D. infernale имеет 32 хромосомы. Можно привести еще много примеров видообразования в результате полиплоидии.
23.11. Специализация способствует сохранению разнообразия форм, возникших в результате видообразования
Если различные виды выполняют одинаковую экологическую роль, то один из них может исчезнуть в результате межвидовой конкуренции. О таком виде говорят, что он подвергся конкурентному исключению. Виды могут избежать конкурентного исключения, если кавдый из них по-своему специализируется. Если бы не происходило специализаций, то в одно время и в одном месте мог бы существовать только единственный вид. Известно несколько форм специализации, которые способствуют сохранению видового разнообразия.
1. Вид способен адаптироваться к определенному местообитанию и оставаться там большую часть времени или даже все время. Например, некоторые виды усоногого рака обитают в нижней (влажной) части литоральной зоны, в то время как другие виды адаптировались к обитанию в верхней (более сухой) части.
2. Вид способен адаптироваться к определенному времени суток или года и оставаться неактивным все остальное время. Например, белоногая карманчи-ковая мышь впадает в спячку зимой, а ее конкурент, кактусовая мышь, впадает в летнюю спячку в жаркий сухой период.
3. Вид может принадлежать к определенному трофическому типу в зависимости от способа питания. Растения могут существовать вместе со своими потребителями, травоядными животными. Травоядные и плотоядные животные тоже могут жить в одной среде. Их специализация заключается в приспособлении друг к другу и к окружающей среде. Плотоядные приспособлены к потреблению травоядных, травоядные приспособлены к потреблению растений.
4. Приспособленность вида может зависеть от ограничения его пищевого поведения. Это приводит к наиболее сложной форме специализации. Ее называют распределением ресурсов, поскольку она влечет за собой распределение пищевых ресурсов внутри сообщества. Например, волк охотится в основном за крупными копытными млекопитающими; он сам слишком крупное животное, чтобы существовать, питаясь мышами и кроликами. С другой стороны, койот охотится главным образом за кроликами и крысами; он слишком мал, чтобы ловить лосей и оленей. Серая лисица, которая еще меньше, чем койот, питается в основном мышами.
Такая специализация не только предотвращает конкурентное исключение хищников, она помогает предотвратить вымирание и тех видов, за которыми охотятся. Например, мышам приходится приспосабливаться к лисам, а оленям - к волкам.
23.12. Количество обитающих на Земле видов остается более или менее постоянным
Исследования показывают, что видообразование в результате географической изоляции и полиплоидии постоянно приводит к появлению новых видов. В то же время происходит вымирание видов. (Ископаемые данные показывают, что почти все виды, обитавщие в древности, сейчас не существуют. Вымерло более 99% видов.)
Имеющиеся факты указывают, что между образованием и вымиранием видов образуется равновесие. На рисунке 23-6 представлено количество семейств, существовавших, по ископаемым данным, во время каждой эпохи. За исключением значительного увеличения количества семейств, наблюдаемого в тот период, когда организмы начали заселять Землю, и кажущегося их увеличения в самом недалеком прошлом (вероятно, благодаря тому, что наши знания об этом периоде более полные), имеющиеся данные действительно позволяют предполагать, что количество семейств животных, живших в разное время, мало изменилось за 600 млн. лет в океане и за 350 млн, лет на земле. Обратите внимание, что в Период примерно от 350 до 125 млн. лет назад количество семейств колеблется в пределах 325. Поскольку среднее число видов на одно семейство, по-видимому, явдяется постоянным, то и общее количество видов оказывается удивительно стабильным.
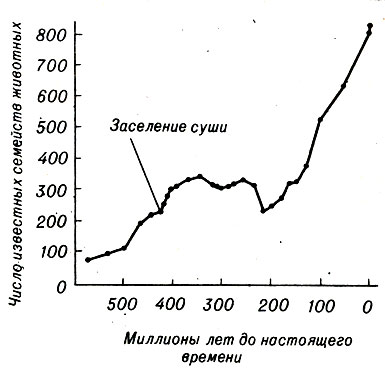
Рис. 23-6. Число известных семейств ископаемых в разные геологические эпохи. (Из: And Replenish the Earth, Rosenzweig.)
Такая длительная стабильность предполагает существование определенного устойчивого состояния. Если число видов превысит постоянную величину, скорость вымирания должна превысить скорость видообразования; если число видов уменьшится, скорость видообразования должна превысить скорость вымирания. Имеются некоторые факты, подтверждающие это, например данные об ископаемых Северной и Южной Америки.
Несколько миллионов лет назад Южная Америка представляла собой огромный остров (как сегодня Австралия). В Северной и Южной Америке были очень схожие условия окружающей среды, и многие формы организмов, обитавшие в Севбрной и Южной Америке, конвергировали. Виды, выполняющие одинаковую роль в разных регионах, называют экологическими заместителями. Когда Северный и Южный Американские континенты соединились в результате вулканической деятельности и образовалась Центральная Америка, стали происходить беспрецедентные миграции: северные виды двинулись на юг, а латиноамериканские виды на север. Это привело к появлению в Северной Америке броненосцев и опоссума, в Южной Америке появились олени и пумы.
Это также привело к контакту многих экологических заместителей и к значительному увеличению количества видов во многих районах. Территориальная конкуренция между заместителями породила волну вымирания. А это, в свою очередь, опять снизило количество видов до существовавшей ранее величины.
Нельзя надеяться, что можно предотвратить вымирание видов. Вымирание - естественный процесс, и он будет продолжаться. Более того, пока будет происходить вымирание видов, пока будут изменяться условия на Земле и пока будет происходить отбор наиболее жизнеспособных организмов, устойчивых к заболеваниям и вредным воздействиям окружающей среды, до тех пор будут продолжаться филетическая эволюция и видообразование. Этот процесс, по-видимому, бесконечен. Пока существует жизнь, будут происходить изменения. Возможно, Гераклит был прав, когда много лет назад утверждал, что наиболее постоянным в мире является изменение. В самом деле, если нужно выбрать один важный объединяющий фактор, характеризующий жизнь и отличающий ее в конечном итоге от чисто физических и химических явлений, это будет эволюция, направляемая естественным отбором. Все живое подвергается эволюционному изменению, а все остальное зависит от условий среды, в которой существует жизнь.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'