22. Адаптация - основной итог эволюции
Ключевые вопросы
Каким образом дрейф генов обусловливает неадаптивную эволюцию?
Какие данные свидетельствуют о том, что эволюция является прежде всего адаптивной?
Несмотря на ряд убедительных доказательств, что эволюция является прежде всего адаптивной и очень редко подвергается влиянию неадаптивных механизмов, некоторые ученые полагают, что неадаптивные механизмы, такие, как мутация, миграция и дрейф генов, представляют собой достаточно важные факторы, обусловливающие неадаптивную эволюцию. Поэтому, прежде чем перейти к рассмотрению некоторых доказательств адаптивной эволюции, мы остановимся на процессе генетического дрейфа.
22.1. Дрейф генов - это изменение частоты аллелей в результате влияния случайных процессов
Дрейф генов чаще всего наблюдается в небольших популяциях, число особей в которых недостаточно велико для действия закона средних чисел. Изменения аллельных частот не имеют приспособительного значения, и их трудно предсказать, иначе говоря, они бесцельно дрейфуют. Но имеются случаи, особенно среди изолированных островных видов, когда дрейф генов оказывался важным фактором эволюции.
Существует экстремальная форма генетического дрейфа, в основе которой лежит эффект родоначальника. Представьте себе, например, остров, на котором нет птиц. Представьте также, что один раз в сто лет соответствующий по силе и направлению шторм в соответствующее время года забрасывает на остров нескольких птиц. Если птицы выживают после путешествия, шансы на возвращение у них ничуть не лучшие, чем шансы остаться на острове. И они, конечно же, не теряют времени в ожидании другого шторма для возвращения. И если местообитание оказывается подходящим для их существования, они основывают новую колонию.
Вскоре остров становится переполненным этим видом птиц. Но происхождения всех особей данного вида связано с теми первооснователями, чей маленький генофонд дал начало целой популяции.
Эти основатели не могли нести с собой все разнообразие генов их родительской материковой популяции. Они имели только свои собственные гены, составляющие лишь часть генофонда родительской популяции. Поэтому новая колония может отличаться от материковой популяции.
Если, например, в определенном o локусе материковой популяции было 100 аллелей, а первооснователей на острове - пять, тогда на острове незамедлительно элиминируются по крайней мере 90% аллелей материковой популяции, т. е. эффект родоначальника предполагает, что дрейф генов возникает в том случае, если несколько особей крупной популяции образуют новую популяцию на какой-нибудь изолированной территории.
Известно несколько примеров действия эффекта родоначальника в изолированных популяциях человека. Некоторые из них были замечены в общинах ами-шей, члены которых вступают в родственные браки. Одно из таких наследственных заболеваний было впервые описано в 1860г. С тех пор зарегистрировано только 100 случаев заболевания, 55 из которых имели место в небольшой группе (8000 человек) амишей в Пенсильвании. Предками всех 55 была пара, прибывшая в Америку в 1774 г. Частота аллеля в этой группе составляет около 0,065. Это намного выше, чем в любой другой известной популяции. Естественно, средняя приспособленность этих людей должна быть ниже, чем в том случае, если бы не произошел дрейф генов.
22.2. Параллелизм и конвергенция подтверждают концепцию о том, что значение адаптивной эволюции более велико, чем неадаптивной, и поэтому в любой окружающей среде направление эволюционного процесса можно предсказать
Для того чтобы проверить, действительно ли естественный отбор преобладает над неадаптивными механизмами, следовало бы провести такой идеальный эксперимент: поместить несколько популяций в одну и ту же новую среду отдельно друг от друга. Если эволюция является адаптивной, тогда каждая популяция в процессе эволюции должна стать лохожей на остальные, потому что, вероятно, существует какой-то наиболее оптимальный путь приспособления в этой среде и адаптивная эволюция должна осуществить его. А неадаптивная эволюция, особенно случайная, дает неодинаковый результат во всех популяциях, обитающих в данной среде.
Природа проводила такие эксперименты множество раз. Иногда результатом являлся параллелизм, а иногда конвергенция. В случае параллелизма отдельные популяции сначала имеют одинаковые фенотипы, потом сильно изменяются, но тем не менее всегда сохраняют некоторое сходство друг с другом. При конвергенции они сначала совершенно непохожи, а потом в ходе эволюции приобретают сходство (часто довольно значительное) (рис. 22-1).
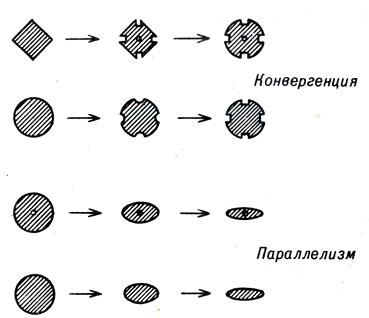
Рис. 22-1. Схематическое изображение конвергенции и параллелизма
Такие "эксперименты" в природе происходят потому, что во многих частях земного шара могут возникать сходные условия окружающей среды, но популяции, которым надлежит адаптироваться к ним, в разных местах часто принадлежат к разным видам.' Например, возьмем популяцию роз, которая должна приспособиться к существованию в пустыне в Мексике, и популяцию молочая, которая должна адаптироваться в условиях такой же пустыни в Танзании. Если межцу этими двумя популяциями произойдет конвергенция, то это должно быть связано с тем, что эволюция является адаптивной и действует таким образом, чтобы изменить растения и животных в наиболее приспособленные для этой среды формы. Но их конвергенция не щюкраща-ется, потому что оба вида были такими разными вначале, что каждый из них, по-видимому, сохранил некоторые основные признаки, отражающие их истинное происхождение.
22.3. Имеется много примеров конвергенции между плацентарными и сумчатыми животными
Во многих частях земного шара для организмов существует возможность зарываться в землю. Но, однако, кроты (плацентарные) распространены не так широко. В Австралии и Африке вместо них обитают другие группы животных.
Одно из сумчатых австралийских животных имеет чрезвычайное сходство с кротом: у него плохое зрение, короткий хвост, довольно толстое, округлое тело, широкие передние копающие ноги и мех, позволяющий ему передвигаться в любом направлении под землей. Очевидно, чтобы жить в таких же условиях, как крот, нужно и быть похожим на него. Конечно, вряд ли нам удалось предсказать эволюцию крота, хотя этот пример конвергенции дает основание предполагать, что когда-нибудь мы сможем предсказывать эволюцию. Это базируется на том, что эволюция является прежде всего адаптивной. Если бы эволюция была случайной и неадаптивной, то такие случаи конвергенции были бы чрезвычайно редкими или, возможно, вообще не существовали.
Сумчатые животные в процессе эволюции приспособились к обитанию в различных условиях, в частности в таких, в которых живут плацентарные животные Нового и Старого Света. Сумчатый волк, например, конвергентно сходен с обычным волком. Он гонится за добычей, опираясь на пальцы ног, как и волки. Существуют также сумчатые летяговые животные, сумчатые крысы и мыши (рис. 22-2).
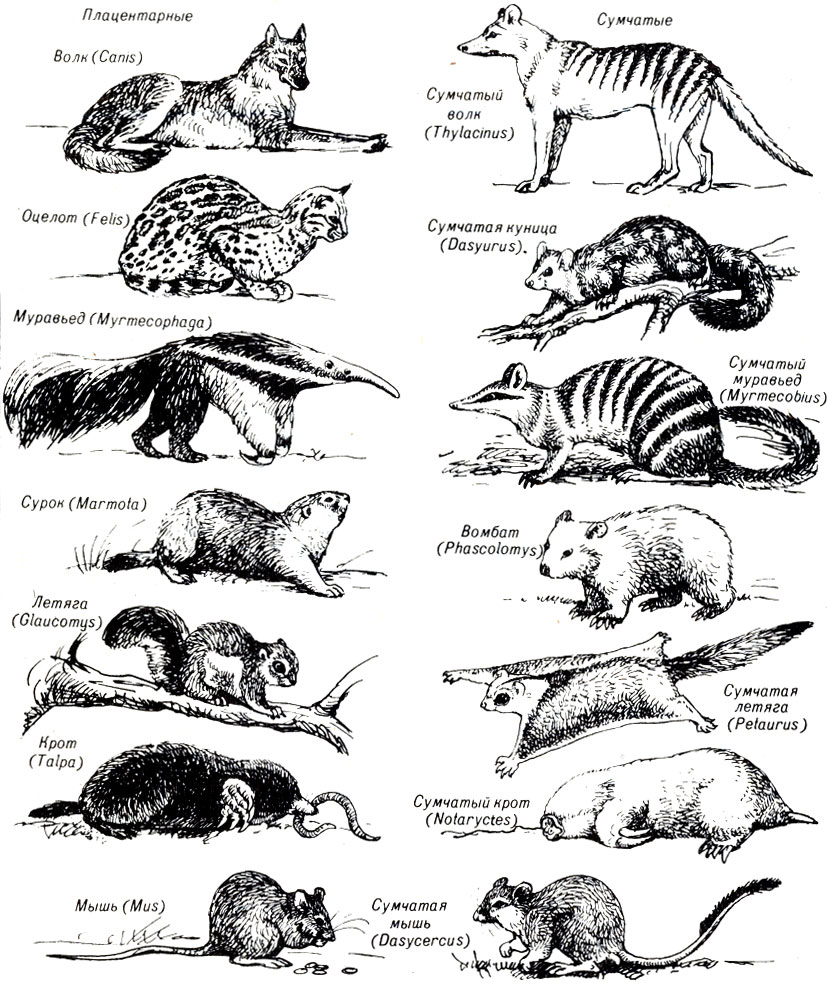
Рис. 22-2. Некоторые сумчатые животные и их плацентарные экологические эквиваленты. (Из: Life: An Introduction to Biologi. George Gay lord Simpson and William S. Beck, copyright 1965, Harcourt Brace Javanovich, Inc.)
Вероятно, самая удивительная конвергенция сумчатых и плацентарных животных произошла у недавно вымерших саблезубых животных (рис. 22-3).
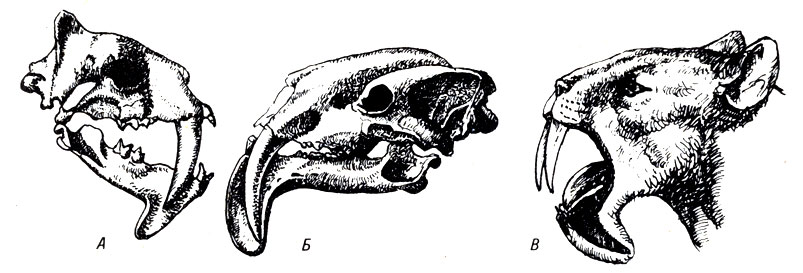
Рис. 22-3. Черепа сумчатых (А) и плацентарных (Б) саблезубых животных. В. Реставрация сумчатого саблезубого животного. (Из: And Replenish the Earth. © 1974, Michael L.Rosenzweig.)
Саблезубые хищники могли достаточно широко открывать рот, так что острый зуб выступал наружу. Затем они вонзали зуб глубоко в тело своей добычи с помощью необычайно сильных мышц шеи. Этому способствовали и другие приспособления скелета, такие, как длинные клыки, особое строение нижней челюсти, позволявшее предохранить зубы от повреждения, а также модификации в строении черепа, позволявшие широко раскрывать рот, чтобы из него выступали зубы, и очень крепкие кости для прикрепления шейных мышц. Плацентарные саблезубые (северное полушарие) и сумчатые (Южная Америка) имели все эти приспособительные признаки. Представьте только, сколько случаев конвергенции в строении мышц сопровождали конвергенцию строения скелета. Но некоторые отличительные признаки черепа сумчатых не изменились, поскольку в этом не было необходимости. Изучая их череп, можно легко определить, что две формы саблезубых животных произошли от совершенно разных предков.
22.4. В условиях пустынь в результате конвергентной эволюции у организмов развились специфические адаптивные признаки
Растения пустынь эволюционировали в колючие и сочные виды, приспособленные сохранять влагу. У многих из них отсутствуют листья и фотосинтез осуществляется в стеблях. Листья имеют сравнительно большую площадь поверхности, поэтому при испарении теряют очень много влаги. К жизни в пустынях Нового Света приспособились кактусы - близкие родственники роз. В пустынях Старого Света влияние окружающей среды привело к развитию у растений, принадлежащих к совершенно другому семейству, Euphorbiacae, такого же комплекса адаптивных механизмов, как у Cactacea (рис. 22-4). Однако цветки молочая сохраняют свою обычную форму, и поэтому их легко можно отличить от кактусов.

Рис. 22-4. Кактусы и сходные с ними по внешнему виду Euphorbia. (Из: And Replenish the Earth, Rosenzweig.)
Один из случаев конвергенции кактусов и других пустынных растений затрагивает их биохимические свойства. Для осуществления фотосинтеза растениям необходим СО2. Чтобы получить его, их устьица должны быть открыты. Но при усвоении СО2 через открытые устьица теряется влага. (Эта потеря воды может быть очень значительной, поскольку СО2 необходим растениям при солнечном свете в период сильной жары.)

Рис. 22-4. Кактусы и сходные с ними по внешнему виду Euphorbia. (Из: And Replenish the Earth, Rosenzweig.)
Кактусы и Crassulaceae приспособились одинаково. Их устьица открываются ночью, когда влажность в пустыне высокая, температура сравнительно низкая и потеря воды минимальная. В это время СО2 накапливается, вступая в реакцию с определенными молекулами, возможно одинаковыми в обоих семействах.
Когда наступает день и температура повышается, растения закрывают устьица и используют СО2, накопленный за ночь.
Условия пустынь оказали влияние и на животных. Покрытые редкой растительностью, пустыни Северной Америки населены многими видами кенгуровой крысы, а в пустынях Африки и Ближнего Востока водятся тушканчики (рис. 22-5). Эти виды животных отличаются только тем, что все ocoби надсемейства кенгуровой крысы (включая гоферовых животных и некоторых довольно распространенных крысовидных грызунов) имеют как бы отороченные мехом защечные мешки, а тушканчики не имеют.

Рис. 22-5. А. Кенгуровая крыса. Б. Тушканчиковая сумчатая крыса. (А представлен проф. К. Schmidt - Nielsen. Б - из: Brehm's Tierleben, Bibliographishes Institut, Leipzig and Vienna, 1914.)
Можно было бы привести намного больше примеров конвергенции у птиц, насекомых, рыб и других организмов.
22.5. Подобно конвергенции, мимикрия позволяет предсказать направление эволюции
В случае с мимикрией можно заранее сказать, каким будет приспособительный фенотип. При мимикрии одни организмы становятся похожими на другие, обладающие высокой степенью приспособленности.
Одну из разновидностей мимикрии называют мимикрией Бэтса, по имени описавшего ее известного естествоиспытателя XIX в. X. В. Бэтса (Н. W. Bates). Мимикрия "Бэтса предполагает участие по крайней мере трех, а возможно, и четырех видов организмов. Должен существовать потребитель, способный распознавать форму, цвет, запахи и т. д. своей любимой пищи и пищи, которая ему неприятна. Должен существовать неприятный для потребителя пищевой вид (который горек на вкус или ядовит) и съедобный вид, напоминающий несъедобный. (Возможно, также должна присутствовать другая съедобная цища, которая отличается от несъедобной.) Несъедобный вид называют моделью, a тот, который напоминает ее, - имитатором.
Поведение съедобного имитатора основывается как на способности потребителя распознавать несъедобную модель, так и на своем сходстве с моделью. Преимущество имитатора состоит в том, что, поскольку он напоминает неприятную на вкус форму, он реже становится добычей. Поэтому смертность организма-имитатора сокращается, а приспособляемость увеличивается.
Один из наиболее известных случаев мимикрии Бэтса наблюдается у бабочек. Гусеницы бабочки данаиды (Danaus plexippus) (рис. 22-6, В) питаются растением ваточником, которое содержит ядовитые химические соединения. Очевидно, поэтому взрослые бабочки становятся совершенно несъедобными. Бабочка Limenitis archippus очень похожа по внешнему виду на данаиду (рис. 22-6, Б), но ее гусеницы питаются неядовитыми растениями. Когда гусеница превращается в бабочку, то последняя потенциально является лакомой пищей для любой птицы, которая может отличить ее от данаиды. Но, поскольку в результате мимикрии Limenitis archippus имеет большое сходство с данаидой, это узнавание превращается в чрезвычайно сложную проблему для дтщь И тголько немногие люди, имеющие специальный опыт, могут различать этих бабочек. В результате птицы вынуждены искать пищу в другом месте.
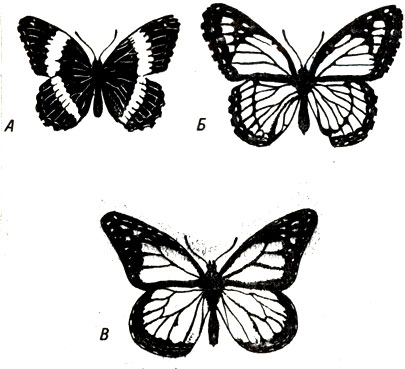
Рис. 22-6. Мимикрия Бэтса, Имитатор, модель и более примитивная форма бабочки, родственная имитатору. В. Бабочка данаида (Danaus plexippus) является моделью и неприятна на вкус. Б. Ее съедобный имитатор - Limenitis archippus; A. L. arthemis напоминает форму, от которой произошла бабочка L. archippus. (Из: Ecological chemistry, Ц. P. Brower. Copyright. © Scientific American, Inc. Авторское право сохранено.)
Я. ван Зандт Бровер (J. Van Zandt Brower) показала значение несъедобной модели для съедобного имитатора. Она скармливала бабочек Limefiitis archippus посаженным в клетку неопытным хохлатым сойкам. Птицы ели их охотно. Но когда их стали кормить бабочками данаидами, они съели всего несколько штук. Как только сойки почувствовали, как неприятна на вкус бабочка данаида, они перестали питаться и данаидами и Limenitis archippus.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'