21. Роль мутации в эволюции
Ключевые вопросы
Каков вклад мутаций в процесс эволюции путем естественного отбора?
Полезно ли большинство мутаций? Можно ли управлять мутациями?
Рональд Фишер (Ronald Fisher) дал самое лучшее определение мутации: "Мутация есть источник любого наследуемого изменения". Он написал это в 1930 г., более чем за два десятилетия до того, как ученые узнали, что гены состоят из ДНК, и начали различать мутации, вызываемые изменениями во внутренней структуре генов, и мутации, вызываемые изменениями в структуре и числе хромосом. Несмотря на эти открытия, определение Фишера остается одним из лучших, потому что оно не зависит от какого-либо определенного механизма мутаций.
Мутации служат сырьем для естественного отбора. Иначе говоря, они являются источником разнообразия живых организмов. Без мутаций естественный отбор не существовал бы; не было бы альтернативных фенотипов и, следовательно, не было возможности выбора. В процессе эволюции мутации приводят к наследственным изменениям, а естественный отбор "подхватывает" те из них, которые увеличивают приспособленность организмов, и "отбрасывает" остальные. На основе этой концепции Фишер сделал вывод, что чем выше наследственная изменчивость в популяции, тем быстрее отбор увеличивает среднюю приспособленность. Так принято считать сегодня, но не всегда мутацию связывали с естественным отбором для объяснения эволюции.
21.1. Многие эволюционисты считали, что эволюция происходит только благодаря мутациям
С конца 1700-х годов был выдвинут целый ряд эволюционных теорий. Многие ученые полагали, что эволюция происходит только в результате мутаций, или в виде адаптивной эволюции, которая увеличивает приспособленность, или в виде случайной эволюции, которая представляет собой случайное изменение, не связанное с приспособленностью. Согласно первой из этих теорий мутации возникают потому, что они необходимы для увеличения приспособленности. Теория адаптивной эволюции имела два варианта в зависимости от того, считалось ли учеными, что приобретенные признаки наследуются или нет.
Ламаркизм - теория эволюции, названная по имени o французского палеонтолога Жана Батиста де Ламарка (Jean Baptiste de Lamarck) (1744-1829), основывалась на том, что приобретенные признаки наследуются. Эта теория предполагает эволюцию в результате мутаций, потому что приобретение нового наследуемого фенотипа эквивалентно мутации. Ламаркисты полагали, что такие изменения увеличивали приспособленность, и любили иллюстрировать это на примере жирафы. Предки жираф, считали они, вытягивали свои шеи для того, чтобы достать листья с верхушек деревьев. Каждое последующее поколение, следовательно, рождалось с более длинной шеей. Этот процесс продолжался до тех пор, пока пропала необходимость вытягивать шею.
Такое утверждение вызвало несогласие многих ученых, и один из них провел эксперимент, в котором он отрезал хвосты у мышей на протяжении нескольких последовательных поколений, однако, как он и предполагал, ему не удалось получить мышей, бесхвостых от рождения. Ни один из наследуемых признаков не может модифицироваться в потомстве приобретенными признаками родителей. Однако приобретенный признак - пересадка парамеции дополнительного ротового отверстия - можно передать негенетическим путем.
21.2. Можно ли управлять мутациями?
Другой вариант теории адаптивной эволюции, как результата мутаций, был более сложным, чем ламаркизм, и сохранился до XX в. Возьмем в качестве примера эволюцию бактериальных штаммов, устойчивых к антибиотикам. Сторонники мутационной эволюции утверждали, что до того, как стали использоваться антибиотики, не было резистентных бактерий, но когда в окружающей их среде появился подобный препарат, он вызвал мутации, которые привели к устойчивости бактерий.
Дарвинисты не были с этим согласны. Они утверждали, что мутации устойчивости к антибиотикам существовали уже до того, как появился пенициллин. Но под воздействием новых условий среды изменилась относительная приспособленность бактерий и бактерии, устойчивые к пенициллину, стали весьма распространенными.
В своем эксперименте С. Лурия (S. Luria) и М. Дельбрук М Delbriick) доказали, что дарвинисты были абсолютно правы. Позднее Дж. и Е. Ледерберги (J. и Е. Lederberg) разработали методику, которую они назвали "методом отпечатков", и подтвердили эти результаты. Оба эксперимента заслуживают подробного описания, но, поскольку второй более прост для объяснения, мы остановимся на нем.
Существуют два основных способа выращивания бактерий в лаборатории - в чашке с желеобразной средой, называемой агаром, и в жидкой среде в пробирке. Каждая среда содержит питательные вещества, но бактерии в чашке с агаром растут на поверхности среды и остаются в фиксированном положении. В любую из сред можно добавить пенициллин, стрептомицин или другой антибиотик, и тогда все нерезистентные бактерии погибнут.
Обычно бактериологи перемещают бактерии с одной чашки с агаром на другую с помощью платиновой петли. Петлей подбирают несколько клеток и переносят их на новую поверхность для продолжения роста.
Новшество Ледербергов заключалось в использовании куска бархата размером с чашку вместо платиновой петли. Бархатный ворс заменил тысячи маленьких игл, каждая из которых захватывала несколько клеток, чтобы перенести их на новую поверхность.
Поскольку бактерии не передвигаются по чашке с агаром, каждое скопление клеток представляет собой генетически чистую колонию, выросшую из одной бактерии. Когда бархат касался поверхности новой агаровой среды, он оставлял отпечаток с предыдущей чашки, т. е. переносил отдельные клетки из каждой колонии, сохраняя их расположение по отношению друг к другу. Благодаря этому Ледерберги могли установить происхождение каждой бактерии в любом месте на новой чашке по положению исходной колонии на предыдущей чашке.
Ледерберги попытались доказать, что мутация устойчивости к антибиотикам произошла до их воздействия на бактерии. Они вырастили бактерии Е. coli на простом агаре и сделали отпечаток на чашке со стрептомицином. При этом выжило только несколько бактерий. Ледерберги определили местоположение их родительских колоний на исходной чашке с агаром. Затем из этих родительских колоний, т. е. из тех точек на исходной чашке, где были получены резистентные бактерии, с помощью петли часть бактерий переносили в пробирки с жидкой средой для быстрого роста. После этого их пересевали на чашки с новой средой, не содержащей антибиотика. Следует помнить, что эти бактерии и их прямые предки никогда не подвергались воздействию стрептомицина. Отпечатки этих новых посевов были сделаны на чашках со стрептомицином. В этот раз погибло гораздо меньше бактерий, чем при первом отпечатке, т. е. не подвергавшиеся воздействию антибиотика родительские формы резистентных бактерий имели мутацию (рис. 21-1).
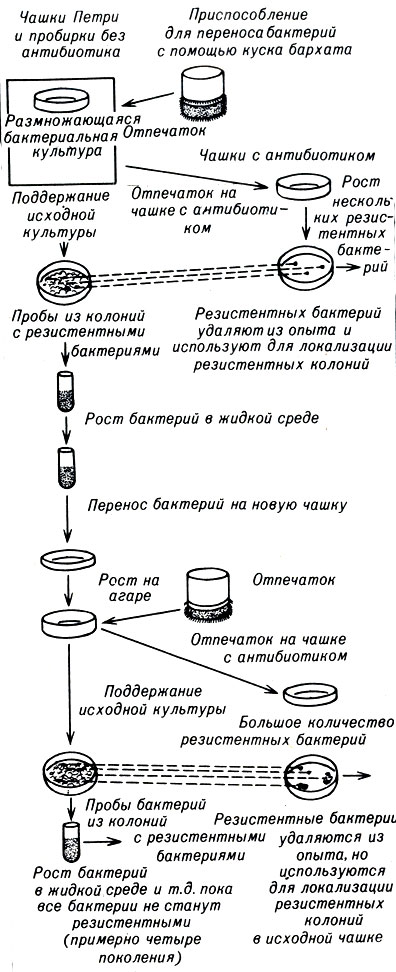
Рис. 21-1. Эксперимент Ледербергов с посевом методом отпечатков
Затем со второй чашки, где отсутствовал антибиотик, с помощью петли Ледерберги захватывали бактерии из тех колоний, которые соответствовали местоположению резистентных бактерий на второй чашке со стрептомицином. Эти захваченные бактерии содержали гораздо большую долю резистентных форм, чем в первом случае. Ледерберги размножали их в жидкой среде и вновь высевали на чашки. При третьем отпечатке на чашке с токсической средой количество резистентных бактерий оказалось еще больше по сравнению с предыдущими отпечатками: чашка была заполнена ими, хотя их предки никогда не вступали в контакт со стрептомицином, т. е. мутация произошла до того, как возникло новое давление окружающей среды. К четвертому посеву методом отпечатков все бактерии оказались резистентными.
Каждый этап этого эксперимента обогащал популяцию бактерий резистентными аллелями. Фактически Ледерберги заменили естественный отбор искусственным, производимым человеком. Они также провели опыты с другими антибиотиками и получили аналогичные результаты.
21.3. Мутации редко бывают полезными
Мутации не только не возникают, когда организму необходимо стать более приспособленным, но, даже если они возникают, они обычно бывают вредными. Менее 1% из сотен наблюдаемых мутаций у ячменя были полезными, а у человека из 10 типов мутаций с известной частотой 8 обусловливают низкую относительную приспособленность. Влияние же остальных двух типов мутаций, альбинизма и аниридии (отсутствия радужной оболочки глаза), на приспособленность неизвестно. В эксперименте с лабораторными популяциями Dro-sophila melanogaster, в которых частота мутаций увеличивалась с помощью действия рентгеновских лучей, 95% мух даже не дожило до взрослого возраста и очень малое количество мух дало плодовитое потомство.
Хотя обычно мутации бывают вредными, все же, когда возникает мутация, способствующая вопроизведению организмом потомства в своем поколении, естественный отбор приводит к увеличению той части популяции, которая несет данный аллель. Этот же механизм сдерживает распространение вредных аллелей. Именно благодаря тому, что естественный отбор увеличивает приспособленность и приводит к образованию популяций хорошо адаптированных особей, большинство мутационных изменений оказываются вредными. Например, если вы почти в совершенстве играете в теннис, тогда попытка использовать новые приемы почти наверняка помешает успешной игре.
21.4. Случайные мутации являются одним из факторов эволюции, но этот фактор играет меньшую роль по сравнению с естественным отбором
Мутации возникают независимо от того, увеличивают они приспособленность или уменьшают ее. Однако часто они приводят к изменению фенотипа и, таким образом, вовлекаются в эволюционный процесс. Тогда что же такое эволюционное давление, производимое мутацией? (Мы будем говорить о генной мутации, но те же принципы можно отнести к молекулярной и хромосомной мутациям или любому другому типу мутаций.)
Предподожим, что в локусе имеются два аллеля: В и Y. Существует шанс, что вредный аллель В превратится в полезный Y, a Y мутирует в В. Этот шанс может быть невелик, даже близок к нулю, но он существует. Каким же может быть конечный результат эволюции, обусловленный этими мутациями (в отсутствие отбора)?
Представьте себе фантастический эксперимент с двумя сосудами, содержащими аллели В и аллели Y. Представьте также, что в нем участвуют два джина (наподобие джинов из сказок "Тысяча и одна ночь"): один из них вынимает из сосуда аллели В, изменяет их в Y и бросает в сосуд с аллелями У. Другой джин делает обратное. Единственным правилом является то, что джин В->Y должен изменить точно X процентов генов В за поколение; за это время джин Y->В из меняет Z процентов генов F.
Который из джинов выигрывает? Ни тот, ни другой. Если начнет выигрывать джин Y->B, он исчерпает свой собственный набор генов Y. Это даст джину B->Y преимущество в дальнейшей игре. Фактически, когда число превращенных генов Y равняется числу превращенных генов В, достигается равновесие.
Генетики выяснили, что фактическая частота генных мутаций очень низка: от пяти на миллион до восьми на сто тысяч или около этого. Поскольку большинство мутаций бывают вредными, то их низкие частоты сами по себе факт благоприятный. Многие эволюционисты полагают, что частоты генных мутаций стали такими низкими в процессе эволюции, так как организмы с высокой частотой мутаций менее приспособлены. (Известны гены, влияющие на частоту мутаций.)
Мы уже отмечали, что во всех случаях, исключая гетерозис, один из двух аллелей данного признака элиминируется в результате отбора, направленного против него. Но элиминация аллеля не является полной, потому что существует возможность восстановления частоты этого аллеля. Когда аллель полностью исчезает, он возникает из другого аллеля в результате мутации. Хотя силы отбора и мутации не равны, между ними устанавливается равновесие. Обычно отбор доминирует, и мутации едва удается предотвратить полное исчезновение вредных аллелей.
Примером является аллель карликовости у человека. Люди с одним нормальным аллелем и одним аллелем карликовости являются хондродистрофиками (рис. 21-2). (Люди с двумя аллелями карликовости нежизнеспособны.) Хотя в результате отбора хондроди-строфики должны были бы исчезнуть, мутации поддерживают сохранение этого аллеля.
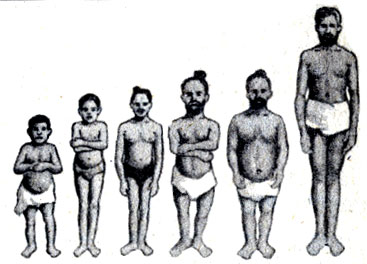
Рис. 21-2. Три типа карликовых взрослых людей. Два карлика слева от взрослого человека нормального роста являются хондродистрофиками. Всем этим лицам было более 20 лет в момент фотографирования. (Из: And Replenish the Earth. 1974, Michael L. Rosenzweig.)
Датский исследователь Е. Т. Морч (Е. Т. Morch) провел анализ в семьях 108 карликов и выяснил, что, хотя у карликов родилось только 27 живых детей, у их 457 нормальных братьев и сестер родилось 582 ребенка. Ясно, что отбор работал против карликовости. Затем Морч измерил частоты мутаций, изучив записи 94 075 новорожденных в Копенгагенской больнице. Только 10 из всех этих детей были карликами - хондродистрофиками. Но 2 из 10 были рождены родителями карликами, т. е. их карликовость не была обусловлена новой мутацией. Таким образом, из 94 073 потенциально нормальных новорожденных у 8 была мутация" карликовости. Поскольку каждый из 94 073 детей имеет два гена этого локуса, то общее число локусов было 188 146. В восьми произошла мутация, следовательно, мутационная частота (х) равна 8/188146, или 42,5 мутаций на миллион генов.
Такая низкая частота вовсе неудивительна. Мутаций же в сторону нормализации не наблюдалось. Между отбором, направленным против карликовости, и мутацией, образующей 8 аллелей карликовости на 188 146 нормальных аллелей, установилось равновесие. Равновесие, которое наблюдал Морч, выражалось в том, что частота аллеля карликовости составляла 10/188150.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'