10. Физиология высших растений
Ключевые вопросы
Какие свойства воды наиболее важны для понимания физиологии растений?
Как вода движется по растению? Существуют ли другие механизмы, помимо осмоса?
Как листья используют воду?
Для осуществления каких функций растениям требуются минеральные вещества и какие?
Как растения используют азот? Откуда они его получают?
В главе 9 было показано, что огромного разнообразия и процветания достигли в процессе эволюции растения подтипа Spermopsida, или семенные растения, и особенно цветковые (покрытосеменные). Наблюдающий такое разнообразие физиолог естественно задает вопрос: какие же свойства позволили так широко распространиться этим растениям? Например, как вода достигает вершины огромного мамонтового дерева? Для того чтобы понять этот и другие важные вопросы физиологии растений, необходимо познакомиться с физическими и химическими свойствами воды, из которой они в основном состоят.
10.1. Физические свойства воды можно понять на основании ее молекулярной структуры
Вода, в отличие от других обычных соединений, существует как газ, жидкость и твердое вещество в пределах нормальных температурных колебаний, существующих на Земле. Более того, при сравнении с другими подобными ей химическими соединениями точки замерзания и кипения воды кажутся удивительно высокими. Эти неожиданные свойства воды обусловлены ее необычной молекулярной структурой.
Молекулы воды являются полярными, они несут слабый отрицательный заряд на атомах кислорода и слабый положительный заряд на атомах водорода. Вследствие этого электростатически притягивается кислород других молекул и между ними образуются водородные связи. При достаточно низких температурах молекулы воды сохраняют связь друг с другом, образуя решетчатую структуру, известную как лед. Естественно, встречающаяся структура льда является менее плотной, чем вода в жидкой форме. Если бы было наоборот, то лед образовывался бы на дне озер. Это происходит с другими соединениями, замороженная форма которых является более плотной, чем жидкость.
При температуре выше 100°С водородные связи между молекулами воды постоянно формируются и разрушаются, но ниже 100°Св каждый данный отрезок времени существует достаточное количество связей для удержания молекул воды вместе, в результате образуется жидкость. Именно для разрушения водородных связей требуется большое количество тепловой энергии, чтобы разъединить молекулы воды и перевести ее либо из твердого состояния в жидкое, либо из жидкого в газообразное. Например, чтобы превратить. 1 г воды в водяной пар, требуется 102 кДж. Вся эта энергия расходуется на разрушение связей между молекулами воды и освобождение их друг от друга: при этом температура воды не изменяется. Энергию в 102 кДж тепла называют скрытой теплотой парообразования. Нагреваемые солнцем клетки листьев могли бы свариться, если бы тепло не удалялось путем испарения воды. По такому же принципу потоотделение охлаждает человеческое тело.
Вода имеет высокое специфическое тепло. Это означает, что вода может поглощать большое количество тепловой энергии, не изменяя температуры.
Наличие водородных связей придает молекулам воды способность к прочному сцеплению и поверхностному натяжению. Подъем воды от корней дерева до листьев на высоту 70 м и более был бы невозможен, если бы не эта способность воды к сцеплению.
10.2. Вода движется в клетки и из клеток растений благодаря осмосу
Кратко познакомившись с физическими свойствами воды, мы можем теперь перейти к рассмотрению того, как она ведет себя в организме растения. В общем, растению требуется такое количество воды, какое может обеспечить листьям или другим фотосинтезирующим частям нормальное функционирование. По мере расходования воды клетками листа она должна быстро восполняться, иначе лист увянет и погибнет. Вода, необходимая листьям, поступает через корни. Механизм этого процесса в общих чертах представляет собой диффузию в ответ на осмотический градиент.
Осмос можно определить как специальный случай диффузии воды через полупроницаемую мембрану в ответ на разницу в концентрации воды по обе стороны-мембраны. У наземных растений осмос происходит через клеточные мембраны корневых волосков и других клеток в корне. Осмотическое давление в этих клетках и клетках, проводящих воду от корневого волоска до ксилемы и далее до листьев, много выше, чем осмотическое давление почвенной воды, вследствие высокой концентрации растворенных солей и Сахаров в клетках. Следовательно, вода стремится войти в эти клетки, понижая их осмотическую концентрацию. Прежде чем поступающая в корень вода достигнет ксилемы и будет транспортирована дальше, она должна пройти либо через клетки, либо между ними к энтодерме. Энтодермальные клетки имеют специальные полупроницаемые мембраны, через которые осуществляется осмос. Поскольку существует градиент в концентрации воды от почвы к корневым волоскам, далее к клеткам энтодермы и ксилеме, вода фактически течет в направлении этого осмотического градиента под давлением, известным как корневое давление.
10.3. Ни корневое давление, ни всасывание, ни активность стебля не ответственны за транспорт воды. Как же может вода достигнуть листьев на вершине стометрового мамонтового дерева?
Хотя корневое давление может быть продемонстрировано на ряде растений, давление редко бывает достаточным, чтобы заставить столб жидкости подняться более чем на несколько метров. Несмотря на это, деревья в жаркий летний день доставляют десятки литров воды к верхушкам ветвей.
Удивительный транспорт воды в растениях нельзя объяснить наличием процесса всасывания. Гипотеза, что растения имеют систему нагнетания воды из корней путем всасывания, с первого взгляда может показаться возможной. Однако известно, что, когда насос нагнетает воду в трубу (так же как в сосуд ксилемы), он фактически дает возможность атмосферному давлению толкать воду вверх. В данном случае атмосферное давление способно поднимать столб жидкости примерно на 10 м. Таким образом, всасывание не может объяснить, как вода достигает вершины дерева высотой 100 м.
В начале века была популярна гипотеза, что растения могут каким-то образом активно перекачивать воду вдоль стеблей. Некоторые ботаники утверждали, что в этот процесс вовлечен определенный пульсирующий ритм, или "жизненное движение". Гипотеза была опровергнута немецким ботаником, срубившим двадцатиметровый дуб и каким-то образом поставившим его в огромную пробирку с пикриновой кислотой, которая убивала клетки растения по мере продвижения вверх по дереву. Через три дня он заменил кислоту водой. Как и. предсказывалось, вода поднялась до верхушки дерева так же легко, как и прежде. Следовательно, было установлено, что активное всасывание воды клетками, требующее их метаболической активности, не играет решающей роли в перемещении влаги вверх по растению.
10.4. Лучшее объяснение транспорта воды дает теория натяжения-сцепления
Теория натяжения-сцепления при транспорте воды в растениях, впервые сформулированная три четверти века назад, основывается на двух свойствах воды: на способности к сцеплению, или на степени, с которой ее поляризованные молекулы проявляют тенденцию притягивать друг друга, и на ее адгезивности к гидрофильным веществам. Вместе эти свойства дают возможность тонкому непрерывному слою воды, который существует в трахеидах соснового дерева или в более эффективных сосудистых элементах клена, противостоять разрыву и вытеканию через боковые стенки сосудов ксилемы. Движение воды в этих непрерывных столбиках осуществляется благодаря тому, что клетки листьев используют ее на вершине столба для фотосинтеза и теряют некоторую ее часть через устьица в результате испарения. Однако не вся вода, выделяемая через устьица, представляет собой отходы; она также регулирует температуру листьев в жаркую погоду.
Хотя для физиологов растений недостаточно ясны некоторые детали теории натяжения-сцепления, ее правомерность может быть установлена простым, однако не щадящим растение экспериментом.
Если ввести булавку или другой острый предмет через кору в ксилему, например, молодого растения дуба, а затем извлечь их, то можно услышать продолжительное "шипение". Оно обусловлено поступлением в поврежденные сосуды ксилемы воздуха, который разрушает столб жидкости.
Когда листья теряют воду в результате испарения, создается градиент давления вдоль водяного столба от почвы через ксилему к листьям. Если потеря воды происходит достаточно быстро, как это имеет место в жаркий летний день, давление внутри ксилемы становится ниже атмосферного и увеличивается поверхностное натяжение. Когда поверхностное натяжение в соке ксилемы превышает осмотическое давление в клетках корня, вода втягивается вверх. Вместе с водой транспортируются минеральные вещества, необходимые для осуществления метаболических процессов в клетках листьев. Натяжение в этих условиях не эквивалентно всасыванию. Одним всасыванием нельзя объяснить транспорт воды у растений.
10.5. Листья, осуществляющие фотосинтез, используют воду для различных целей. Фотосинтез регулирует скорость испарения воды и газообмена через устьица
В реакциях фотосинтеза вода под действием света расщепляется, выделяя свободный кислород и образуя соединения, которые служат для восстановления диоксида углерода (IV) в реакциях, протекающих в отсутствие света. Таким образом, вода является основным "сырьевым материалом" для фотосинтеза. Вода также смачивает поверхность фотосинтезирующих клеток внутри листьев и способствует поглощению другого сырьевого материала для фотосинтеза - углекислого газа.
Суммарная потеря воды при испарении с поверхности фотосинтезирующих клеток в листьях растения называется транспирацией, ее скорость регулируется активностью устьиц. Устьица открываются на свету и закрываются в темноте. Фотосинтез контролирует эти движения, обеспечивая энергией процесс переноса ионов калия в клетки устьиц. Этот процесс, в свою очередь, регулирует осмотический потенциал, необходимый для открытия устьиц.
На транспирацию уходит около 95% воды, поглощенной растениями, оставшиеся 5% используются для роста и фотолиза воды. В течение сезона роста одно растение томата теряет около 130 л воды в результате транспирации. Пальмовое дерево, растущее в жарком сухом климате, где испарение происходит быстро, может использовать 380 л воды в день для транспирации, фотосинтеза и регуляции температуры.
Потеря воды листьями в процессе транспирации является важной функцией, обеспечивающей транспорт растворенных минеральных солей и других веществ к тканям, которые в них нуждаются.
10.6. Для роста и осуществления полного жизненного цикла растениям необходимо около 16 основных минеральных веществ
В прошлом столетии ученые, изучавшие водные культуры, установили, что для роста растений необходимы азот, фосфор, калий, кальций, магний, сера и железо. Эти элементы нужны растениям в больших количествах, и поэтому их называют макроэлементами.
В нынешнем столетии в исследованиях, проведенных на водных культурах, были установлены еще девять необходимых растениям элементов, или микроэлементов. Последние требуются в меньших количествах, и их функции в растении менее четко определены. К микроэлементам относятся железо, бор, марганец, цинк, медь, молибден, кобальт, хлор и натрий.
Среди всех этих элементов наиболее важную роль играет азот. Многое известно о том, как он функционирует в растениях. Хотя свободный азот (N2) составляет 80% земной атмосферы, высшие растения не имеют механизмов для утилизации этого исключительного инертного соединения. Они получают азот из почвы, где он существует в виде неорганического нитрата. Нитрат, в отличие от некоторых других минеральных веществ, попадает в почву не за счет выветривания и механического разрушения горных пород.
Непосредственным источником нитрата являются продукты распада растений и животных, в организме которых азот входит в состав органических веществ.
В результате деятельности почвенных бактерий органические соединения теряют связи углеродов с водородом и превращаются в неорганический нитрат (рис. 10-1).
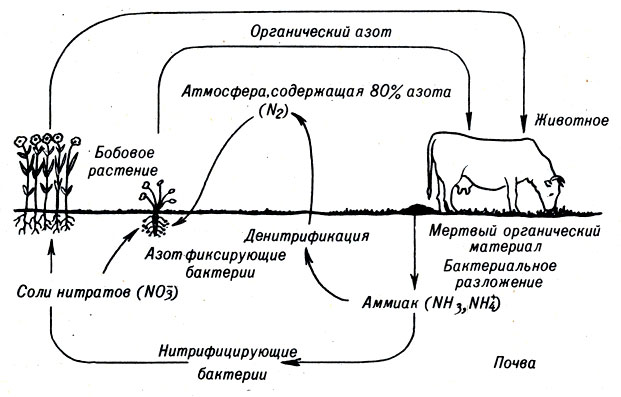
Рис. 10-1. Круговорот азота в природе
В этой форме азот поглощается корнями растений.
Примерно такой же цикл превращений происходит в водной среде, где растения поглощают неорганический нитрат и используют его для образования аминокислот, белков, нуклеиновых кислот и т. д. Часть растений и животных возвращают азот в окружающую среду в составе органических отходов их жизнедеятельности либо в виде останков собственных организмов. Затем, как и на суше, бактерии превращают органические азотсодержащие соединения в неорганические.
Растения потребляют нитрат и вновь переводят его в органическую форму, главным образом в аминокислоты.
Образование аминокислот может осуществляться в корнях, но чаще всего происходит в листьях, к которым транспортируется нитрат.
10.7. Путем трансаминирования некоторые аминокислоты, синтезированные в корнях, превращаются во все виды аминокислот, необходимых растению
Аминокислоты, особенно глутаминовая кислота, синтезируемая в корнях, поднимаются вверх по растению с током воды. Там происходит процесс, известный как трансаминирование, при котором аминогруппа (NH2) одной аминокислоты последовательно переносится к ряду других соединений, вследствие чего образуется новая аминокислота, например:
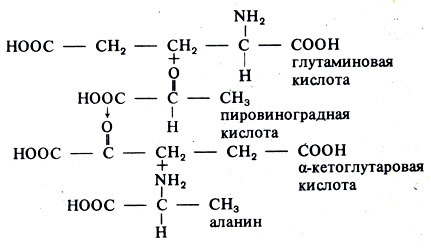
Таким путем синтезируются все 20 аминокислот, необходимые для построения белков. Растения также синтезируют витамины (кофакторы) и необходимые им метаболиты. Таким образом, они являются значительно более независимыми, чем животные. Последние не могут производить все необходимые аминокислоты путем трансаминирования и, следовательно, должны получать многие из них с пищей.
10.8. Весь азот, используемый растениями и животными, первоначально поступает из атмосферы и превращается в приемлемые для использования формы азотфиксирующими бактериями или сине-зелеными водорослями
Фиксация азота является процессом, при помощи которого определенные сине-зеленые водоросли и бактерии, например Rhizobium, превращают атмосферный азот в азотные соединения, такие, как аммиак.
Rhizobium и другие азотфиксирующие бактерии обитают в тканях корней бобовых, таких, как клевер, люцерна, горох, соевые бобы, и вызывают образование неровных белых клубеньков, называемых узелками (рис. 10-2). Бактерии, находящиеся в этих узелках и существующие в симбиозе с бобовым растением, поглощают из корня растворенные сахара и, в свою очередь, снабжают растение соединениями азота. Если корни бобовых оставить в почве после окончания сезона роста, они будут служить богатым источником азота для получения следующего урожая на этом поле. Именно поэтому бобовые часто используются в севообороте.

Рис. 10-2. Клубеньки, содержащие Rhizobium, на корнях шелковой акации Albizzia. (Фото R. Speck.)
Многие растения не вступают в симбиотические взаимоотношения с азотфиксирующими бактериями и должны получать нитраты из почвы, образовавшиеся в результате распада растительного и животного материал а. В естественных условиях растение фактически возвращает весь используемый нитрат в почву, но в условиях культурного ведения хозяйства растения постоянно удаляются с земли, лишая таким образом ее естественного набора питательных веществ.
10.9. Нитраты и другие минеральные вещества становятся пригодными для растений благодаря процессу, известному как катионный обмен
Минеральные вещества, необходимые растению для роста, находятся в почве в различных формах. Они могут быть растворены в почвенной воде, адсорбированы на частицах глины или включены в состав органических и неорганических соединений. Минеральные вещества, содержащиеся в растворе почвенной воды и адсорбированные на частицах глины, наиболее пригодны для растений. В обоих случаях минеральные вещества существуют в виде ионов - положительно заряженных катионов, таких, как Са2+ (ионы кальция), Fe3+ (ионы железа) и К+ (ионы калия) или отрицательно заряженных анионов, таких, как нитраты, сульфаты, фосфаты;
Упомянутые катионы и ионы Н+ также проявляют тенденцию группироваться вокруг отрицательно заряженных частиц глины (рис. 10-3).
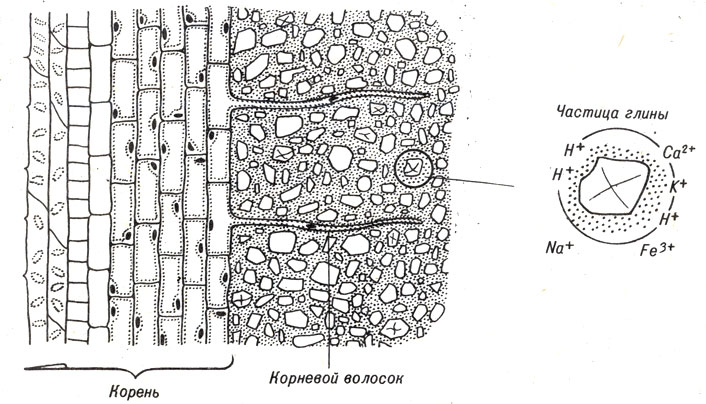
Рис. 10-3. Схема, показывающая взаимоотношения между корневыми волосками и отрицательно заряженными частицами глины, притягивающими катионы, которые являются питательными веществами для растений
Катионы, удерживаемые глиной, способны к обмену. Например, Н+ может заменить Na+ (натрий) или Na+ может заменить Н+.
Катионы, поглощаемые растением с поверхности частиц глины, заменяются другими катионами, обычно Н+. Этот процесс называется катионным обменом.
10.10. Растворенные минеральные соли поступают в корни путем диффузии и в результате процесса активного транспорта
Простой диффузией нельзя объяснить все потребление минеральных веществ, которое происходит у растений. Корни продолжают поглощать ионы минеральных веществ из воды даже после того, как концентрация в тканях корня становится больше, чем в воде. Если данный процесс осуществляется только за счет диффузии, то поток ионов в корни должен замедляться и прекращаться совсем, когда ионная концентрация в корне сравняется с концентрацией воды в почве.
Для активного накопления ионов против градиента концентрации требуется энергия АТФ. Если основой потребления минеральных веществ является активный транспорт, то он скорее имеет место в живых клетках корня, нежели в мертвых клетках ксилемы. По-видимому, энтодерма, окружающая сосудистый цилиндр корня, является тканью, обеспечивающей метаболический контроль.
Тело корня состоит из двух частей: неживой, называемой апопласт, и живой - симпласт. Вода, диффундирующая в растение из почвы, проходит вначале только через апопласт - клеточные стенки и межклеточные пространства - или через живые клетки, пока не достигнет энтодермы. Затем вода и растворенные минеральные соли должны проникнуть через слой энтодермы, т.е. пройти через живые энтодермальные клетки, которые являются частью симпласта (рис. 10-4). Эти клетки на радиальных стенках имеют восковые утолщения (каспаровы полосы), которые препятствуют дальнейшему межклеточному транспорту. Вся вода и минеральные вещества должны таким образом пройти через живую клеточную мембрану, прежде чем попасть в ксилему. Этот сложный путь дает растению возможность отобрать вещества, которые ему необходимы.
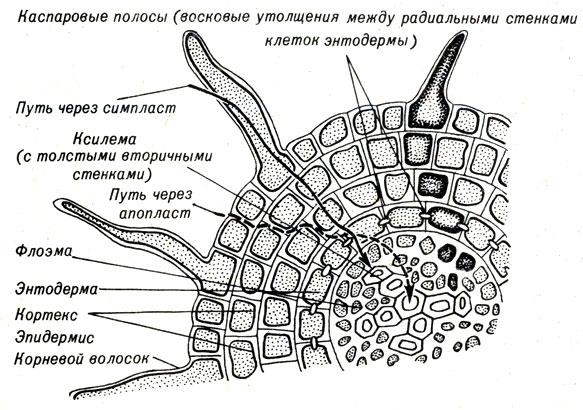
Рис. 10-4. Схема корня, показывающая симпласт, апопласт и пути, по которым вода и ионы могут попадать в корень. Пространства, занимаемые стенками, увеличены, чтобы иллюстрировать эти пути
В свою очередь, симпласт путем активного метаболизма накапливает ионы и переносит их через наружную мембрану клеток. Как только растворенные вещества попадают в энтодерму, они легко проходят к сосудам ксилемы, по которым пассивно переносятся к стеблю и листьям.
10.11. Растворенные питательные вещества распространяются по растению вместе с водой, которая движется по флоэме
Движение Сахаров, аминокислот, гормонов и даже вирусных частиц происходит главным образом в тканях флоэмы, которые находятся под корой деревьев и в кортексе у других растений. В настоящее время считают, что основная масса растворенных веществ переносится вместе с водой, которая быстро передвигается по флоэме. Это основной поток. Перемещение происходит за счет неравного осмотического давления в различных клетках. Например, осмотическое давление будет отличаться в клетке, активно использующей фотосинтетические продукты, и в клетке, еще находящейся в процессе фотосинтеза.
Долгое время не удавалось получить для исследования образец содержимого флоэмы (сока). Положение изменилось, когда в 1953 г. для этой цели стали использовать тлю. При питании тли помещают трубковидные хоботки в отдельную клетку флоэмы (рис. 10-5). Чистый сок может быть собран из
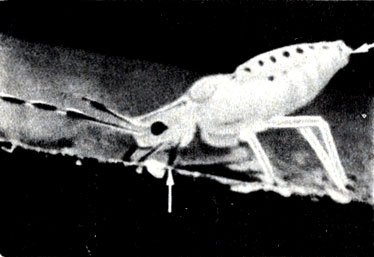
Рис. 10-5. Хоботок тли, сидящей на листе липы (стрелка), погружен в клетку флоэмы, такую же, как показана на схеме листа (рис. 9-17). Обратите внимание на каплю, выделяющуюся из брюшка. (Фото автора.)
хоботков, отделенных от тела насекомых. Анализ химического состава сока показал, что он содержит около 10-25% сахара. Это самая высокая концентрация, которая имеет место где-либо в растении. Движение Сахаров по флоэме происходит очень быстро. Некоторые исследователи считают, что это объясняется только одной диффузией.
10.12. Растения и их клетки обладают разными видами движений
Хотя растения не проявляют подвижность в том смысле, в каком она имеет место у большинства протист и многих животных, они не являются полностью неподвижными. В клетках изолированных высших растений наблюдается ци-топлазматическое течение, и хлоропла-сты могут перемещаться в ответ на изменение интенсивности и качества света - процесс, называемый фотокинези-сом. Предполагается, что эти типы движения могут быть связаны с движениями такого же рода у животных и протист.
Очень медленно движутся стебли высших растений - растут в направлении света. Корни растений обычно растут вниз в почву. Эти движения относятся к тропизмам, точнее, к фототропизму (движение в ответ на дневной свет) и геотропизму (движение в ответ на гравитацию). У растений могут также наблюдаться более быстрые движения в ответ на изменение освещенности. Подсолнечники в открытом поле весь день находятся лицом к свету, а их листья движутся вверх и вниз, чтобы получить максимальное количество света. Некоторые цветки открываются утром и закрываются ночью. Большинство этих движений является результатом изменения тургора клеток, вызываемого различиями скорости фотосинтеза и количества воды в отдельных клетках. Некоторые из этих движений происходят независимо от света и являются примерами эндогенных ритмов, или биологических часов.
10.13. Многие растения проявляют фотопериодичность. Их цветение регулируется длиной дня и зависит от спектрального состава светового потока
Одни растения цветут независимо от длины светового периода их суточного цикла, и их называют светонейтральными растениями; другие цветут только при условии длинного или короткого дня. Получаемый свет влияет на листья. Это было показано в экспериментах, в которых листья пересаживались с одного растения на другое с целью изменения времени цветения последнего. Различные фотопериодические эффекты были зарегистрированы в ответ на действие света красной и дальней красной частей спектра на белковый пигмент фитохром, обнаруженный во многих растительных клетках. Этот белок может существовать в двух формах, которые отличаются физиологической активностью и способностью поглощать свет. Используя свет определенной длины волны, удается регулировать состояние фитохрома в клетках растений и изменять эффекты длинного и короткого дня. Многие физиологические процессы высших растений и водорослей управляются механизмами, включающими фитохром. Выяснение этих механизмов представляет собой одно из актуальных направлений исследований в физиологии растений.
Данная глава, очевидно, поможет объяснить, как функционируют высшие растения, хотя существует много фактов в отношении растений, которые еще недостаточно ясны. Например, неоднократно сообщалось, что растения обладают сверхъестественными психическими силами, что они реагируют по-разному на "хороших" и "злых" людей. Большинство ученых рассматривают такие утверждения со здравым скептицизмом, пока эти факты не подтвердятся в строго научных условиях компетентными людьми.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'