9. Высшие растения и их ткани
Ключевые вопросы
Какие признаки высших растений способствовали их процветанию на Земле?
Почему мхи и печеночники имеют более мелкие размеры, чем большинство наземных растений? Какие изменения в жизненном цикле сопровождали эволюцию сосудистых растений?
Почему семена и пыльца благоприятствуют выживанию цветковых растений?
Какое значение имеют растения для человека? Какие специфичные взаимоотношения существуют между цветковыми растениями и насекомыми, птицами и млекопитающими? Каково происхождение этих взаимоотношений?
Каковы основные органы и ткани высших растений? Каковы их функции?
Почему покрытосеменные считаются более высокоорганизованными, чем голосеменные?
Считается, что два типа высших растений - Bryophyta (мхи и печеночники) и Tracheophyta (папоротники и семенные растения) - произошли от линии зеленых водорослей (Chlorophyta), по-видимому, через древнего представителя Charophyta. Биохимические доказательства в поддержку этой гипотезы приведены в таблице 4. Логично предположить, что ранние наземные растения обитали в пресной воде на мелководье и их выживание на суше первоначально было адаптацией к засушливым условиям.
Признаками, которые, по-видимому, способствовали успешному развитию самых древних наземных растений, были следующие: 1) многоклеточность и размер; 2) клеточная дифференцировка; 3) короткий жизненный цикл с половым размножением, приводящим к максимальному увеличению генетического разнообразия, необходимого для естественного отбора; 4) структуры и механизмы, предотвращающие высыхание и поддерживающие газообмен; 5) многоклеточные половые органы для защиты гамет; 6) внутреннее оплодотворение для защиты эмбрионов.
Первые наземные растения, которые обнаружены в виде ископаемых остатков, существовали около 400 млн. лет назад. На протяжении 100 млн. лет с момента их появления многие области Земли были покрыты густыми лесами, населенными различными груцпами растений, которые в настоящее время или вымерли, или представлены очень небольшим числом видов. История эволюции наземных растении - одна из самых удивительных, хотя много волнующих вопросов пока еще не решено.
9.1. Один из основных типов высших растений, Bryophyta, не содержит сосудистой ткани, что приводит к ограничению размеров этого растения
Мхи, печеночники и роголистники типа Bryophyta не имеют ни корней, ни настоящих листьев. Прикрепляются они с помощью нитевидного, не впитывающего влагу ризоида и имеют уплощенные стебли, которые иногда неправильно называют листьями. Мхи всасывают воду и неорганические вещества через стебли. Для оплодотворения необходима водная среда. Подвижные сперматозоиды образуются в мужских половых органах, или антеридиях, растения мха, которое является гаметофитным поколением (рис. 9-1). После дождя сперматозоиды подплывают к яйцеклеткам, находящимся в женских половых органах, или архегониях. Оплодотворение приводит к образованию зиготы, развивающейся внутри архегония в поколении спорофита. Спорофит образует ножку со спорангием, содержащим споры, которые распространяются ветром. Каждая спора может сформировать протонему, молодое нитевидное растение, на котором образуются почки, дающие начало гаметофитному растению.
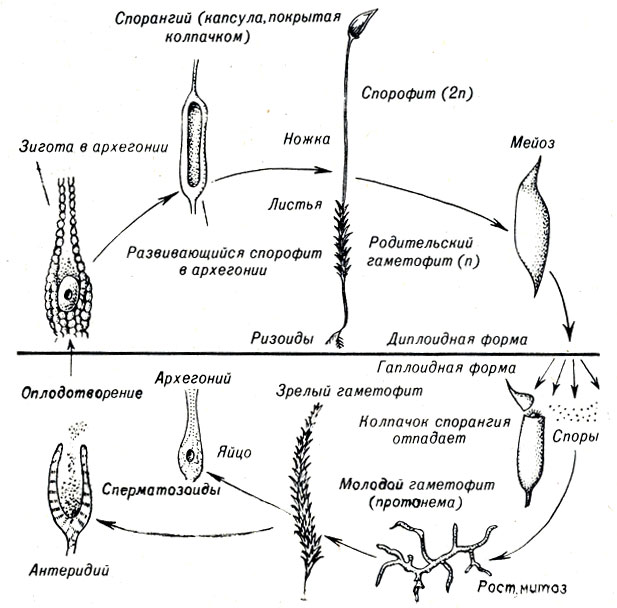
Рис. 9-1. Жизненный цикл мха Polytrichum (тип Bryophyta)
Существуют тысячи видов мхов, однако наиболее значительным по биомассе является Sphagnum, растущий на болотах в различных климатических зонах, от умеренной до арктической. Ежегодные слои прироста Sphagnum разлагаются, превращаясь в торф - горючее, наиболее широко используемое в северных странах. Торфяные болота благодаря консервирующей способности позволяют получать важную историческую и научную информацию. В них сохраняется пыльца растений и даже человеческие останки. В некоторых болотах были найдены настолько хорошо сохранившиеся останки древнего человека, что удалось получить информацию об особенностях питания человека несколько тысяч лет назад.
Печеночники менее заметны, чем мхи, и обычно располагаются на скалах, стволах деревьев или бревнах. Они обнаруживают тенденцию к уплощению и являются листостебельными по виду (рис. 9-2).

Рис. 9-2. Marchantia - обычный печеночный мох (тип Bryophyta). (Фото R. Speck.)
9.2. Сосудистые растения представляют второй основной тип высших растений. Сосудистая ткань дает им решающее преимущество перед мохообразными в конкуренции за солнечный свет и жизненное пространство
В отличие от мхов и печеночников, которые растут, образуя густую подстилку на почве, сосудистые растения (тип Tracheophyta) в большинстве своем растут вверх по направлению к солнцу. Несмотря на то что сосудистые растения варьируют в размере от крошечной Wolffia, примерно с булавочную головку, до гигантского дерева секвойи более 100м в высоту, они сходны во многих отношениях. В целом они имеют: 1) обширную корневую систему, служащую для прикрепления и для поглощения питательных веществ из почвы; 2) широкие листья, содержащие сосудистую ткань и органы фотосинтеза; 3) сильные древесные стебли, способные удерживать их в вертикальном положении. Очевидно, благодаря сосудистой ткани эти растения освободились от ограничения в размере.
Сосудистые растения были и остаются на протяжении более 400 млн. лет доминирующей формой растений на Земле. Их доминирование обусловлено, с одной стороны, васкуляризацией и, с другой, значительной модификацией жизненного цикла, в котором имеется чередование между бесполым (спорофитным) и половым (гаметофитным) поколениями. Трахеофиты отличаются от мохообразных главным образом тем, что гаметофитная часть их жизненного цикла сильно сокращена. У мхов поколение спорофита является отдельным растением, которое, вырастая из женского полового органа, распространяет споры и затем отмирает. У папоротников спорофит является преобладающим поколением, а гаметофит существует как отдельное, но сильно редуцированное растение. Эта тенденция в направлении редукции гаметофитного поколения, заложенная у мхов и папоротников, доведена до крайней степени у семенных растений, где гаметофит существует как паразит, находящийся в наружных покровных тканях спорофита. Спорофит производит два вида гаплоидных спор: мегаспоры, которые дают начало яйцеклетке, и микроспоры, развивающиеся в пыльцу. Пыльца и яйцеклетка представляют собой растения, соответственно, мужского и женского гаметофитного поколений. Оплодотворение восстанавливает диплоидный набор хромосом, и эмбрион развивается в следующее поколение спорофита.
Это основное изменение в жизненном цикле, несомненно, привело к значительному превосходству семенных над мохообразными, поскольку пыльцевые зерна, которые могут противостоять сильному холоду и засухе, являются более надежным средством распространения гамет, чем подвижные сперматозоиды. Появление семян также способствовало прогрессивной эволюции сосудистых растений, поскольку они прорастают только в благоприятные периоды.
9.3. Пять основных групп сосудистых растений последовательно доминировали на Земле: одна группа, по-видимому, вымерла, три другие сохранились частично, в то времй как семенные растения, особенно цветковые, стали доминирующей формой и продолжают эволюционировать
Изучение ископаемых форм растений показало, что первыми распространившимися на суше формами были псилопсиды (подтип Psilopsida). Они представляли собой простые растения, не имеющие корней или листьев, но с ветвистыми зелеными стеблями, способными к фотосинтезу. Ксилема стебля была достаточно прочной, чтобы поддерживать их рост в высоту до 10-90см.
До недавнего времени многие исследователи считали, что два представителя этого подвида выжили: Psilotum (метелочковый папоротник, растущий на юго-востоке США, рис. 9-3) и Tmesipteris, найденный на южных островах Тихого океана. Полученные недавно доказательства говорят о том, что эти организмы являются скорее примитивными папоротниками, чем псилопсидами. Если это так, то псилопсиды, по-видимому, вымерли.

Рис. 9-3. Фотография метелочкового папоротника (Psilotum nudum). (Фото R. Speck.)
Плауны (подтип Lycopsida) впервые появились 350-400 млн. лет назад. В течение примерно 100 млн. лет они развились в деревья высотой до 30 м, которые росли густыми лесами. Хотя эти деревья имели листья и корни, у них отсутствовали некоторые преимущества семенных деревьев, развившихся позже. Впоследствии плауны не выдержали конкуренции за солнечный свет и жизненное пространство и стали вымирать. Наиболее известным сохранившимся плауном является Lycopodium (или живучка ело-чковидная), часто используемый в качестве новогодней зелени (рис. 9-4).

Рис. 9-4. Живучка елочковидная, Lycopodium, с терминально расположенными спорангиями. (Фото M.Stewart.)
Хвощи (подтип Sphenopsida) оставили след в ископаемых остатках примерно в то же время, что и плауны, и некоторые их представители, развившиеся также в гигантские древовидные растения, были преобладающими формами в лесах каменноугольного периода. До настоящего времени сохранился один род Equisetum (рис. 9-5). Растения этого рода редко поедают животные из-за того, что их эпидермальные клетки содержат неперевариваемые кремневые отложения. Американские колонисты называли эти растения "чистящий камыш" и использовали их для чистки посуды. Equisetum имеет корни, полый узловатый стебель с сосудистой тканью по периферии и чешуйчатые листья, радиально расположенные в узлах. Жизненный цикл этих растений включает способные к фотосинтезу отдельные спорофитные и более мелкие гаметофитные растения. Гаметофит производит подвижные сперматозоиды.

Рис. 9-5. Чистящий камыш (Equisetum) неподалеку от группы вечнозеленых секвой в Калифорнии. (Фото автора.)
Папоротники (подтип Pteropsida) также были многочисленной и процветающей группой растений каменноугольного периода (рис. 9-6).

Рис. 9-6. Древовидный папоротник (Cyathea) и травянистый папоротник (Glycenia) в бобовниковом лесу Пуэрто-Рико. (Фото М. Stewart.)
Однако в настоящее время известно около 11 000 видов папоротников, большинство которых распространено в тропиках. Их размеры варьируют от нескольких сантиметров до гигантских форм.
Все лиственные формы папоротников являются спорофитами, а их гаметофит, много меньших размеров, существует отдельно.
Они были первыми наземными растениями, у которых возникли листья, пронизанные сосудами. Кроме того, большая площадь листа позволяла более эффективно осуществлять фотосинтез. Эти качества, несомненно, дали им преимущество над конкурентными формами. Стебли папоротников не очень прочные, и поэтому они обычно растут в защищенных местах, в узких ущельях, оврагах и небольших долинах.
9.4. Семенные растения, являющиеся, несомненно, самой прогрессивной формой наземных растений, впервые возникли как голосеменные
У семенных растений (подтип Spermopsida) появились два основных преимущества по сравнению с более ранними наземными растениями, включая папоротники. Образование пыльцы устранило зависимость от водной среды, необходимой для передвижения сперматозоидов. Формирование семян позволило видам выживать в неблагоприятных условиях (сильная жара, холод, засуха или темнота), в которых само растение не могло бы существовать. Пыльца и семена способствовали развитию новых эффективных путей, облегчающих половое размножение и распространение видов.
Представители класса Gymnospermae (голосеменные) были первыми семенными растениями, появившимися примерно 350 млн. лет назад. Постепенно эта группа заняла господствующее положение в лесах и привела к вымиранию почти всех других форм за исключением нескольких видов плаунов, хвощей и папоротников. Значительно позже в зоне умеренных лесов голосеменные были сами вытеснены покрытосеменными, или цветковыми деревьями.
Современные голосеменные имеют хорошо известных представителей трех групп. Хвойные (шишконосные) включают сосны, пихты, болиголовы, ели, кипарисы, секвойи и можжевельники. У можжевельников шишки подверглись модификации и образовали "ягоды". У других хвойных созревают шишки двух типов: пыльцевые и семенные (рис. 9-7). Мужскими и женскими гаметофитами являются микроспоры и мегаспоры на чешуйках соответствующих шишек.
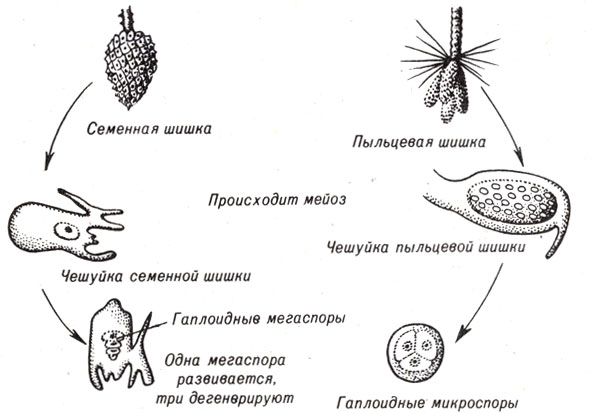
Рис. 9-7. Два типа шишек на сосне или другом хвойном дереве - семенная шишка и пыльцевая шишка - и развитие женского и мужского гаметофитов внутри них
Хвойные наиболее успешно развивались в условиях, неблагоприятных для покрытосеменных растений с жесткой древесиной. Хвойные леса занимают огромные пространства, простираясь от арктической тундры на юг на многие сотни километров. В горных районах на небольших высотах произрастают жесткодревесные растения, уступающие место на больших высотах хвойным, которые распространяются до границы леса. Хвойные леса растут на границе с пустынями, где почва слишком сухая для поддержания жизни жесткодревесных растений.
Некоторые хвойные встречаются среди самых крупных и старейших растений на Земле. Секвойи в Калифорнии вырастали на протяжении последних 3000-4000 лет до высоты бблее 100 м и имели стволы диаметром около 10м.
Одно такое дерево содержит достаточно древесины, для того чтобы построить небольшой дом. Пиломатериалы, получаемые из секвойи, дугласовой пихты, ели и сосны составляют более чем три четверти общего запаса лесоматериалов США.

Рис. 9-8. Гиннго (Cainkgo btloba) и один из его листьев. (Фото Т, Н. Esekett.)
Из представителей двух других групп голосеменных стоит упомянуть гинкго двухлопастный, Ginkgo biloba (рис. 9-8), которое вряд ли могло сохраниться др настоящего времени, если бы его не культивировали как дерево, дающее тень, и саговники (рис. 9-9), тропические пальмоподобные деревья со стеблями и широкими листьями. (Настоящие пальмы являются покрытосеменными.) Гинкго и саговники, в отличие от всех других голосеменных, образуют голую яйцеклетку, которая оплодотворяется подвижными сперматозоидами? В этом отношении эти растения уникальны среди голосеменных.

Рис. 9-9. Cycas - саговнин, растущий в ботаническом саду. Замбия, Ньясаленд. (Фото М. Stewart.)
9.5. Цветковые растения (класс Angiospermae) представляют собой вершину эволюции царства растений
Цветковые растения, или покрытосеменные (покрытые семена), появились относительно недавно на нашей планете и в сравнительно короткое время, примерно в 130 млн. лет, стали преобладающей формой земной растительности. Многие из первых покрытосеменных были, по-видимому, деревьями, хотя возможно, что более ранние мелкие представители не сохранились в летописи ископаемых форм. Ранние покрытосеменные деревья постепенно вытеснили своих травянисты х родственников. Успешное развитие покрытосеменных позволило им дивергировать в различных направлениях, дав начало многим известным не древесным растениям, таким, как травы, травянистые растения, кустарники, и множеству цветковых растений, которые произрастают в полях, лесах, болотах, пустынях, горах и на отмелях.
В целом примерно четверть миллиона покрытосеменных были описаны и получили названия. Ботаники предполагают, что почти такое же количество еще ждет своего описания.
9.6. В отличие от более древних голосеменных растений покрытосеменные появились в одно время с насекомыми, птицами и млекопитающими; вероятно, поэтому у них развились определенные специфические взаимоотношения и взаимодействия
Размножение голосеменных зависит исключительно от ветра и потоков воздуха, необходимых для распространения пыльцы и семян, поскольку наземные животные появились позже 300 млн. лет назад. Покрытосеменные же развивались как раз в то время, когда сим биотические взаимоотношения с насекомыми, птицами и млекопитающими могли способствовать их эволюции. У цветковых растений появились признаки, которые привлекали внимание насекомых, птиц и млекопитающих. Образующийся нектар (перерабатываемый пчелами в мед) привлекал насекомых, которые переносили пыльцу от растения к растению и обеспечивали таким образом перекрестное опыление. Даже использование пыльцевых зерен в качестве пищи способствовало их распространению.
Эйснер (Т. Eisner) недавно показал, что изображения цветков на пленке, чувствительной к ультрафиолетовому свету, являются образцами опознания (рис. 9-10), которые служат сигналом для некоторых насекомых, указывающим, что "пища может находиться здесь". В сферу взаимодействия покрытосеменных растений с животными обычно включен только один вид каждого из них, и поэтому отсутствие необходимого вида животных может привести к прекращению размножения растения.

Рис. 9-10. Калужница болотная (Caltha palustris): А. В видимом свете. Б. В ультрафиолетовом свете. Темные, поглощающие ультрафиолет, базапьные зоны лепестков являются 'указателями' нектара. (Фото Т. Eisner.)
Не у всех покрытосеменных опыление зависит от насекомых. У некоторых форм растений оно осуществляется с помощью ветра, вследствие чего миллионы людей страдают от так называемой сенной лихорадки. Лихорадка обычно вызывается пыльцой деревьев, летний насморк - травянистыми растениями, и аллергические заболевания ранней осенью - амброзиями и шалфеями (рис. 9-11). У цветковых растений семена распространяются ветром чаще, чем пыльца, например у одуванчика или клена.

Рис. 9-11. Амброзия (Ambrosia trifida) - причина широко распространенного недуга многих людей, страдающих осенней сенной лихорадкой. (Фото R. Speck.)
Птицы и млекопитающие также играют важную роль в распространении семян. Существуют самые различные способы распространения семян, например выплевывание их обезьянами (тропические фрукты), перенос на шерсти медведя (дурнишники), хранение под землей запасов, не использованных белками (шишки, орехи и т. д.), или прохождение через кишечник животных и откладка в плодородных фекальных массах.
Об одном аспекте размножения цветковых растений следует упомянуть особо, поскольку он очень важен для эволюции этой группы. Речь идет о двойном оплодотворении - образовании двух ядер сперматозоида, одно из которых оплодотворяет яйцеклетку, а другое - эндосперм, обеспечивающий эмбрион питательной средой.
Многочисленные преимущества, которые приобрели покрытосеменные, привели к возникновению большого разнообразия функций и мест обитания растений (рис. 9-12). Существуют многие водные цветковые растения подобно Elodea и Lemna (ряска), некоторые из них являются морскими подобно валлисне-рии (Zostera).

Рис. 9-12. Группа покрытосеменных: А - Zostera marina-валлиснерия, одна из немногих морских представителей. Б - Elodea - элодея, часто используется в аквариумах, В - Iris - ирис, однодольное. Г - Heiianthus - подсолнечник. Д - Sarracenia - непентес. Е - Cuscuta - повилика, паразитическое цветковое растение. Ж - Drosera - росянка. 3 - Dionaea muscipula - венерина мухоловка (абориген Каролины, США)
Повилика - паразитирующее растение - не способна к фотосинтезу и поэтому обвивается вокруг других растений для получения питательных веществ.
Цветковое растение вертляница также не способно к фотосинтезу и получает необходимые питательные вещества из грибов.
Росянки и венерины мухоловки стали плотоядными и питаются пойманными насекомыми.
9.7. Покрытосеменные подразделяются на два отряда - - однодольные и двудольные
Самые первые листья, которые появляются из растительного эмбриона семени, называются семядолями. Примерно треть всех покрытосеменных имеют одну семядолю и известны как однодольные растения (отряд Monocotyledonae).
Эта группа включает такие хорошо известные растения, как осоки, ситники, касатики и лилии, тюльпаны, амариллис, орхидеи, кукуруза, лук, пальмы и некоторые другие.
Более многочисленными являются двудольные растения (отряд Dicotyledonае), имеющие две семядоли. Они включают горох, морковь, магнолию, одуванчик, тыкву, подсолнечник и фактически все кустарники и деревья.
Эти две большие группы растений также отличаются структурой их тканей.
9.8. Растения необходимы человеку, так как служат ему пищей, укрытием и источником кислорода. Они синтезируют не только массу полезных веществ, но и небольшое количество сильно действующих ядов
Исследователи, изучающие доисторический период развития человека, считают, что первоначально он был травоядным и стал плотоядным или всеядным только после открытия огня. Даже сегодня большая часть продуктов питания человека состоит из растений и растительных продуктов.
Большинство растений, употребляемых в пищу человеком, являются покрытосеменными. Мы едим листья лука, шпината, капусты, стебли сельдерея и ревеня, корни моркови и редьки, клубни картофеля, цветки цветной капусты и каперсы, чашелистики артишоков, завязь томатов, эмбрионы кукурузы и фасоли, побеги люцерны, эндосперм кокосовых орехов (молоко) и семена подсолнуха и тыквы.
Фактически все используемые в пищу растения появились относительно быстро, в течение короткого периода существования человека, и многие их изменения были получены путем тщательного отбора растений, селекционируемых по некоторым желательным признакам. Разведение растений успешно практиковалось задолго до появления науки генетики. Процесс селекции продолжается, и фрукты, зерно и овощи становятся более вкусными (иногда), сочными и устойчивыми к заболеваниям.
Большинство напитков производят из растений. Кофе и какао получают из бобов, а чай - из листьев; все они обладают тонизирующим действием.
Из растений получают многие лекарственные препараты. Хинин (для лечения малярии и мышечных спазм) извлекают из коры дерева; опиум, из которого изготовляют успокаивающие средства и героин, получают из мака; дигиталис, сердечный стимулятор и тоник, получают из наперстянки. Многие растительные продукты, используемые в качестве лекарственных препаратов, принадлежат к классу алкалоидов. Курительный табак содержит очень сильный и ядовитый'4 алкалоид - никотин, который оказывает сильное воздействие на сосуды и центральную нервную систему. Широкое использование табака для курения вызывает тревогу среди экспертов здравоохранения. Например, существуют доказательства, что от 50 000 до 100 000 американцев ежегодно умирают от постепенно развивающихся эффектов его пагубного воздействия. Если бы этот погребальный звон мог быть предсказан до того, как табак приобрел популярность, то нет сомнения в том, что его использование было бы запрещено законом. Сегодня эта пагубная привычка достигла в популяции такой степени распространения, что справиться с ней пока не удается никакими мерами принуждения.
Растительным продуктом является и марихуана, которую получают из сорной травы Cannabis sativa. Этот препарат обладает способностью изменять настроение и поведение человека. Употребление марихуаны довольно широко распространилось в ряде стран Америки и Западной Европы. Недавно полученные данные позволяют предполагать, что постоянные психические расстройства и потеря полрвой потенции являются возможными последствиями продолжительного использования марихуаны.
9.9. Процветание высших растений можно понять только на основе знания их клеточной и тканевой структуры
Все растения, описанные в этой главе, имеют общий план строения, их клетки соединяются в ткани, а они, в свою очередь, в органы, каждый из которых выполняет определенную функцию в целом организме.
Корни, стебли и листья являются основными органами, поддерживающими жизнь растения.
Прежде чем перейти к рассмотрению функциональных органов растений, необходимо вспомнить характерные признаки типичной неспециализированной растительной клетки. Как вы помните, растительная клетка состоит из цитоплазмы, окруженной цитоплазматической мембраной, и твердой клеточной оболочки.
Все, что содержится внутри клеточной оболочки, известно как протопласт.
Вакуоль занимает центральную часть зрелой клетки и отделена от цитоплазмы вакуолярной мембраной, или тонопластом.
Таким образом, можно себе представить, что растительная клетка в принципе похожа (но не по форме) на футбольный мяч - кожаное покрытие, противостоящее давлению изнутри, представляет собой клеточную оболочку, камера - живую цитоплазму, а воздух в мяче - вакуоль.
9.10. Клеточная оболочка отчасти обеспечивает ригидность растений. Она состоит из микрофибрилл, заключенных в матрикс пектина, и других веществ
Ригидность растений обусловлена комбинацией следующих факторов: составом клеточных стенок и тургорным давлением самих клеток.
Вначале около растущей клетки образуется первичная оболочка: будучи относительно тонкой и эластичной, она может расти с клеткой до определенного предела. Основным компонентом первичной оболочки является целлюлоза, полимер глюкозы. Целлюлоза вытянута в микрофибриллы, которые сплетены вместе и заключены в пектин, другой полимер Сахаров. Такая Структура микрофибрилл и пектина образует жесткую, упругую оболочку.
Когда растительные клетки созревают, они обычно формируют новую, более плотную вторичную оболочку внутри первичной. Вторичная оболочка является более устойчивой к растяжению, и ее большая ригидность обусловлена присутствием воска, лигнина и целлюлозных волокон. Твердость растительной ткани обусловлена высоким содержанием лигнина. В отличие от целлюлозы лигнш! не вплетен в стенку, а врастает в пространство между целлюлозными волокнами.
Клеточные оболочки, будучи ригидными, сохраняют свою проницаемость. Существующие в них отверстия создают возможность соседним клеткам поддерживать связь между собой путем образования протоплазматических мостиков. Эти протоплязматпческие соединения - плазмодесмы (рис. 9-13) - вытягиваются через тонкие отверстия в оболочке, называемые порами.
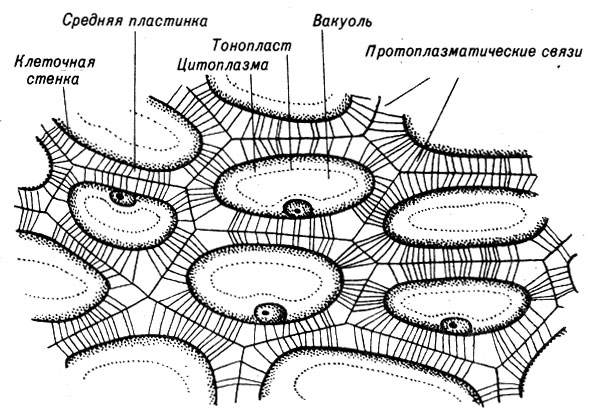
Рис. 9-13. Схема клеток в питательной ткани семени
Между оболочками соседних клеток имеется слой гибкого межклеточного "цемента" - средняя пластинка, которая состоит в основном из пектина. Она формируется до появления первичных или вторичных клеточных стенок, поскольку является структурой, которая образует первую перегородку между делящимися дочерними клетками.
9.11. Вакуоли служат местом отложения запасных питательных веществ и накопления продуктов диссимиляции, они также помогают регулировать движение воды от клетки к клетке
Вакуоли, занимающие до 90% объема клетки, используются для накопления Сахаров, крахмала и белков, которые могут потребоваться клетке в будущем. Отходы также откладываются в вакуолях, где они время от времени формируют кристаллы или гранулы инертного материала.
Возможно, наиболее важной функцией вакуоли является ее роль в регуляции движения воды от клетки к клетке.
9.12. Защитная ткань, основная ткань и проводящая ткань - все они произошли из меристематической ткани
В растении имеются определенные области - меристемы, где при благоприятных условиях происходит недетерминированный рост клеток. Клетки, из которых состоят меристемы, являются неспециализированными, тонкостенными и содержат много мелких вакуолей. Эти клетки впоследствии дифференцируются, образуют лиоо защитную ткань, такую, как поверхностный слой, либо эпидермис листьев, корней и коры стебля, либо основную ткань, включающую губчатую паренхиму, в которой происходит фотосинтез, либо проводящую ткань, такую, как ксилема и флоэма, которые связаны с транспортом веществ.
9.13. Ткани в стебле растения обеспечивают опору, защиту, рост и транспорт питательных веществ и воды
Помимо поддержания листьев для получения достаточного количества солнечного света, стебель функционирует как транспортная система, несущая воду и питательные вещества всем частям растения.
Два вида проводящих, или сосудистых, тканей ответственны за эту функцию. Ксилема переносит воду с растворенными в ней веществами вверх от корней к стеблям и листьям; флоэма транспортирует органические вещества, включая питательные вещества, образованные в результате фотосинтеза листьями, к остальным частям растения. Обе эти ткани развиваются из апикальных меристем или из другой меристематической ткани, камбия. Каждый сезон клетки, которые находятся на внутренней поверхности камбия со стороны водно-транспортной зоны дерева, дифференцируются, образуя толстостенные проводящие воду клетки. Эти клетки ксилемы образуют древесину дерева. Клетки ксилемы погибают, как только достигают зрелости, но и после этого вода и минеральные соли могут пассивно проходить через них.
Некоторые длинные тонкие клетки ксилемы называют трахеидами (рис. 9-14). Эти клетки расположены в виде длинных цепей, которые перекрывают друг друга. Каждая трахеида имеет около миллиметра или меньше в длину и пронизана порами, позволяющими воде проходить через боковые части оболочки клеток и окружающие трахеиды к другим частям растения. Таким путем вода доставляется от клетки к клетке, от корней вверх, через стебель к листьям.
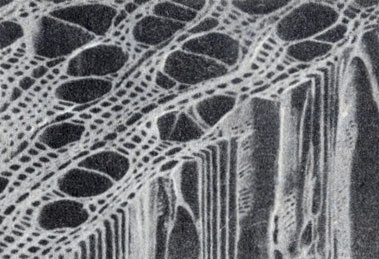
Рис. 9-14. Минрофотография поперечной и радиальной поверхности древесины новозеландского бука (Nothofaqus fusca), полученная с помощью сканирующего электронного микроскопа. Видны тонкие трахеиды и значительно более крупные сосуды ксилемы. (Фото R. R. Exley, В. G. Butterfield и В. A. Meylan и juornal of Microscopy.)
Второй тип ксилемной клетки называют сосудистым элементом, он напоминает микроскопический цилиндр. Клетки соединяются концами и образуют трубки, называемые сосудами ксилемы. Вода проходит через сосудистые элементы без препятствий, так как концевые части оболочки зрелых клеток утрачиваются. Таким образом сосуды ксилемы образуют прямые линии трубок, которые тянутся от корней к листьям. Вторичные стенки сосудов ксилемы не такие толстые, как стенки трахеид, и дают меньшую опору растению. Древесина обычно содержит как трахеиды, так и сосудистые элементы.
9.14. В отличие от ксилемы, доставляющей растению воду и минеральные соли, флоэма транспортирует питательные вещества
Клетки флоэмы развиваются из внешней поверхности камбия. Они не имеют сильно утолщенных стенок, и концевые части оболочки клеток не разрушаются. Они напоминают группу удлиненных цилиндров, соединенных концами друг с другом и связанных порами, которые находятся в концевых частях оболочек. Эти части оболочек называют ситовидными пластинками. Хотя ядра клеток флоэмы распадаются, их цитоплазма остается живой и они активно транспортируют молекулы питательных веществ. Цитоплазма клеток движется через отверстия ситовидных пластинок, образуя таким образом непрерывный ток питательных веществ вдоль ситовидной трубки (рис. 3-6).
По всему растению расположены относительно неспециализированные паренхимные основные клетки. Паренхимные клетки специального типа, клетки-спутники, объединены с клетками, составляющими ситовидные трубки. В отличие от клеток ситовидных трубок клетки-спутники имеют функционирующие ядра, которые, по-видимому, регулируют деятельность клеток ситовидных трубок, лишенных ядер. Когда гибнет клетка-спутник, связанная с ней клетка ситовидной трубки также погибает.
В умеренном климате ксилема древесных растений (не однолетних) имеет годичные кольца. Каждый год в течение сезона роста камбий производит новую ксилему и новую флоэму. Весной, когда много воды , образуются крупные клетки ксилемы, а поздним летом - мелкие. Весенняя и летняя древесина вместе составляют годичное кольцо. Путем подсчета этих колец можно определить возраст дерева.
9.15. Кора служит для защиты стебля
Внутренняя зона коры древесного стебля состоит в основном из меристема-тического слоя, называемого пробковым камбием, снаружи расположена пробка (рис. 3-6). Стенки клеток самого наружного слоя импрегнированы суберином, водоотталкивающим веществом. Это предохраняет внутренние слои стебля от высыхания, а пробковые клетки - от пропитывания водой. В результате они постепенно отмирают и отпадают. Одновременно из пробкового камбия образуются новые пробковые клетки.
Организация тканей стебля в концентрических цилиндрах является типичной для двудольных. Однодольные растения имеют те же самые ткани, с несколько иным строением. Пучки проводящих тканей у них разбросаны по всей сердцевине стебля, представляющей собой центральную массу свободных паренхимных клеток (рис. 9-15).
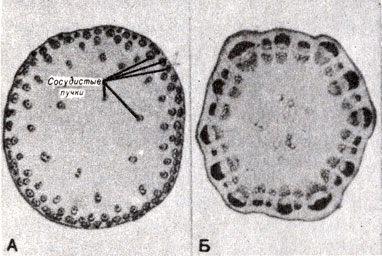
Рис. 9-15. Поперечный срез стебля. A. Zea mays (кукуруза, однодольное). Б. Helianthus (подсолнечник, двудольное). (Фото автора.)
9.16. Ткани корня всасывают воду, растворенные минеральные вещества и фиксируют растение в почве
Корни имеют такое строение, как и стебли, однако у них отсутствует сердцевина, а сосудистая система имеет несколько иное строение. На самом конце корня находится корневой чехлик, который защищает апикальную меристематическую зону делящихся клеток. Новые клетки, образуемые в меристеме, удлиняются и в конечном счете дифференцируются в ксилему, флоэму, эпидермис и другие ткани. В зоне созревания клетки периферийной части корня образуют длинные выросты клеточных оболочек, называемые корневыми волосками (рис. 9-16). Это сильно "увеличивает площадь поверхности корня, через которую он может всасывать воду и минеральные соли. Под эпидермисом корня находится кортекс, многоклеточный слой, самые внутренние клетки которого представляют собой энтодерму, окружающую сосудистую ткань. На радиальных стенках энтодермальных клеток находятся каспаровы полосы, восковые утолщения, которые препятствуют прохождению воды и минеральных солей, минуя ткань энтодермы.
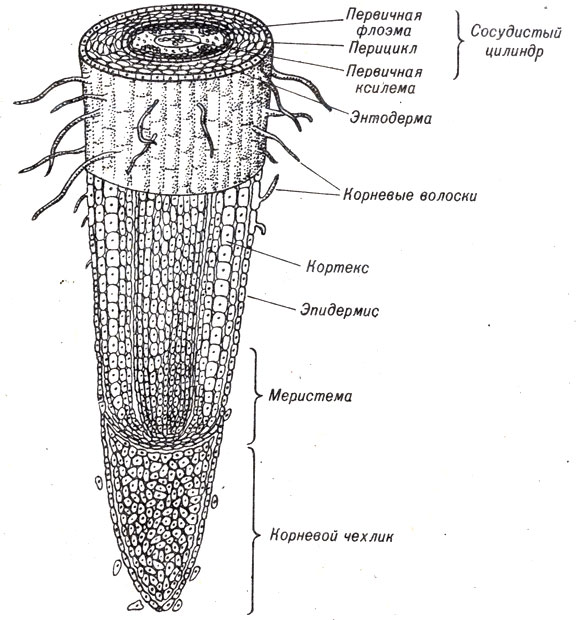
Рис. 9-16. Строение корня, показанное на поперечном и продольном срезах
Корень может также являться местом накопления крахмала и других питательных веществ.
9.17. Лист состоит из двух пористых слоев эпидермиса, между которыми расположена прослойка паренхимных клеток
Листья образуются из меристематической ткани, которая дифференцируется в специализированные клетки. В основном лист состоит из верхнего и нижнего слоя защитных клеток с фотосинтезирующими паренхимными клетками и ветвями сосудистой системы между ними.
В листьях паренхимные клетки имеют большое число хлоропластов (рис. 9- 17). Оболочки паренхимных клеток тонкие и эластичные. Поскольку вторичных оболочек у паренхимных клеток не образуется, они остаются достаточно гибкими, что, в свою очередь, придает эластичность растению.
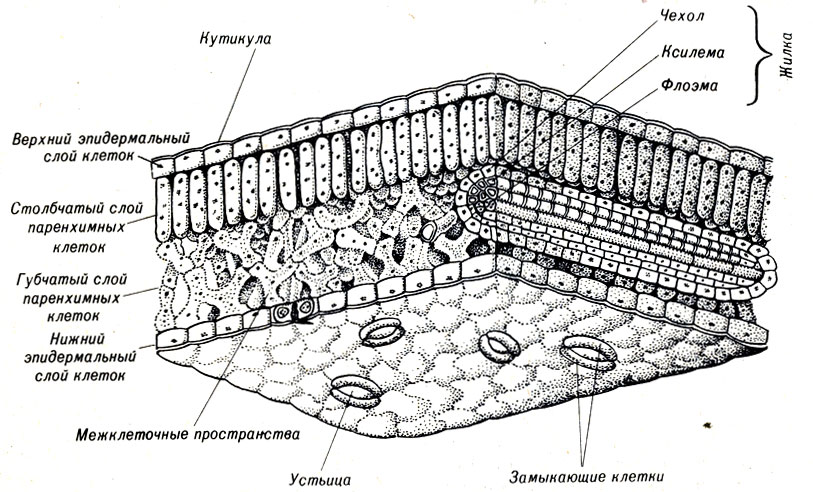
Рис. 9-17. Строение листа, представленное в виде схемы на вырезанном кусочке
Через несколько слоев паренхимных клеток проходят сосудистые пучки в форме жилок. Они содержат как ксилему, так и флоэму и находятся в прямом контакте с основной частью растения. Вода, поступающая через клетки ксилемы, особенно важна для осуществления интенсивного газообмена, происходящего в листьях. В свою очередь, листья посылают через флоэму в другие части растения некоторые синтезируемые в них сахара.
Паренхимные клетки листа ограничены с двух сторон слоями эпидермальной ткани, состоящей из уплощенных, плотно прилегающих друг к другу клеток. Единственным пространством между клетками являются поры, или устьица, через которые диффундирует воздух. Днем фотосйнтезирующие листья осуществляют фотосинтез, поглощая из воздуха СО2 и выделяя О2 и пары воды.
Листья растений имеют специальные структуры, которые регулируют поступление СО2 и потерю О2 и паров Н2О. Парные эпидермальные клетки регулируют открытие устьиц, ведущих во внутренние части листьев, где обычно осуществляется фотосинтез.
Устьица открываются, когда замыкающие клетки, окружающие их, находятся в состоянии набухания, и закрываются при спадении замыкающих клеток в результате потери воды. Снижение фотосинтеза при недостатке воды, закрытие устьиц является важным предохранительным механизмом против увядания и гибели растения. Когда устьица закрыты , вода не испаряется и растение противостоит высыханию.
При избытке воды на свету в замыкающие клетки поступают ионы К+ и АТФ, образующиеся в результате фотофосфорилирования в процессе фотосинтеза. Это приводит к набуханию замыкающих клеток и открытию устьиц.
На 1 см2 поверхности листа яблони расположено примерно 1000 устьиц. Даже в том случае, когда все поры открыты , их общая площадь составляет малую часть площади листа.
Другие клетки эпидермиса обладают водоотталкивающими свойствами благодаря кутину, восковому покрытию наружных клеточных стенок.
9.18. Растения имеют два типа основной ткани - колленхиму и склеренхиму
Колленхима состоит из удлиненных клеток, имеющих толстые клейкие оболочки. Клетки колленхимы являются первыми опорными клетками, которые формирует растение. После закладки клеточных оболочек они удлиняются и создают опору для листьев, корней и стеблей. Клетки другой ткани, склеренхимы, имеют толстые, упругие клеточные оболочки и выполняют опорную функцию у взрослых растений. После достижения клетками зрелости они перестают расти, затем отмирают и запустевают, но при этом продолжают поддерживать растение.
На сайте https://stavropolsm.com вы сможете найти девушку, соответствующую вашим интересам и предпочтениям.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'