5. Принципы и механизмы процессов развития
Ключевые вопросы
Что означает способность яйцеклетки (или одного из ее бластомеров) к "регуляции"?
Что такое дифференцировка?
Происходила ли в развивающемся зародыше только специализация ядер клеток к считыванию определенных генов или они "забывают", как использовать информацию из любой части генома?
Как узнать, что клетки дифференцированы?
Является ли процесс дифференцировки у животных и растений необратимым?
Если яйцо морского ежа рассечено пополам, важно ли для развития личинки, в какой плоскости произведено разделение?
Что означает установление полярности?
Что такое эмбриональная индукция?
Как ткани развивающегося зародыша влияют друг на друга?
Какую роль в процессах развития играют гормоны?
В главах 3 и 4 было дано описание развития типичных представителей растений и животных без пояснения принципов и механизмов данного процесса.
В этой главе будут кратко приведены наиболее важные обобщения, которые можно сформулировать на основе экспериментального анализа развития.
Современные исследования в биологии развития в значительной мере связаны с изучением молекулярных и клеточных механизмов, которые лежат в основе процессов, происходящих в эмбриональном развитии.
5.1. Геном развивающегося организма подобен библиотеке, в которой отдельные группы клеток в разное время черпают необходимую информацию и поэтому развиваются
ДНК зиготы содержит больше генетической информации, чем обычно используется любой клеткой развивающегося зародыша; тем не менее в каждой клетке имеется полный набор генов, называемый геномом. На самых ранних этапах развития некоторые клетки эмбрионов животных сохраняют способность к регуляции, т. е. могут считывать любые части всей генетической "библиотеки", или генома, и, следуя полученным указаниям, синтезировать белки, необходимые для развития. Поэтому яйцо морского ежа, рассеченное пополам в определенном направлении, разовьется в две нормальные личинки, из которых в конечном итоге сформируются два взрослых организма морских ежей (рис. 5-1, А). Клетки эмбрионов млекопитающих также сохраняют способность к регуляции, правда, только до стадии имплантации в стенку матки. До этого два эмбриона могут быть объединены, а определенные части зародыша способны сформировать либо плаценту (внеэмбриональные оболочки), либо собственно эмбрион. Развитие близнецов также можно вызвать экспериментально.
Однако способность к регуляции у клеток постепенно снижается по мере развития эмбриона, и в дифференцированных частях зародыша клетки утрачивают способность считывать определенные участки генома. О таких клетках говорят, что они дифференцируются, т. е. начинают отличаться друг от друга по степени экспрессии различных генов. Гены контролируют синтез ферментов и структурных белков, наличие или отсутствие которых делают клетки отличными друг от друга. Экспрессия генов в клетках контролируется сложными механизмами и обычно большинство генов репрессировано, или "выключено". Однако во всех клетках эмбриона специфические последовательности генов избирательно дерепрессированы, или "включены", для того чтобы мог происходить синтез необходимых ферментов и структурных белков.
5.2. Экспериментально показано, что генетическая информация в ядрах дифференцированных клеток не утрачивается
В лаборатории Гёрдона (J. В. Gurdon) были проведены эксперименты, в которых ядра из клеток кишечника головастиков Xenopus (шпорцевой лягушки) трансплантировались в яйцеклетки с удаленным ядром. Пересаженные ядра в ряде случаев обеспечивали нормальное развитие до стадии головастика. Эти эксперименты показали, что генетическая информация, как правило, не утрачивается в дифференцированных клетках -она просто не считывается. Дифференцировка у эмбрионов животных, по-видимому, процесс необратимый. Известно, что клетки, в которых начали проявляться различия, могут культивироваться в течение многих поколений без потери способности к дифференцировке. Клетки большинства растений, даже будучи внешне дифференцированными, сохраняют способность к регуляции в подходящих условиях окружающей среды. Было установлено, что единичные клетки, выделенные из корня моркови, могут либо образовывать однородную популяцию дифференцированных клеток, либо, проявляя способность к регуляции, формировать различные группы клеток, способных развиваться в целое растение (морковь). Ключевой фактор, определяющий способность клеток моркови к регуляции, - химический состав окружающей среды. Этот же фактор имеет важное значение и для клеток животных, но в данном случае, вероятно, более существенным является химический состав цитоплазмы.
5.3. Развитие эмбрионов происходит по принципу "только вперед", и уже на ранних его этапах устанавливаются полярность, ось симметрии, определяются будущие части зародышей
Иллюстрацией наиболее ранней и наиболее поздней детерминации могут служить яйцеклетки двух распространенных морских беспозвоночных. Яйцеклетки морского ежа являются примером поздней детерминации. Если на двуклеточной стадии развития разделить оба бластомера яйца морского ежа, то каждый из них разовьется в целый организм. Аналогичный результат будет получен, если бластомеры разделить на стадии четырех клеток. Бластомеры яйцеклеток моллюска, разделенные на тех же дву- и четырехклеточных стадиях, будут развиваться в аномальную личинку, которая не способна сформировать взрослый организм. Иначе говоря, судьба двух клеток яйца моллюска предопределена - они детерминированы уже на этой ранней стадии развития.
Известно, что яйцо морского ежа способно к регуляции, так как каждый ранний бластомер может использовать всю содержащуюся в нем генетическую информацию для формирования целого организма. Однако нельзя быть уверенным, что в данном случае отсутствует детерминация другого рода. Наличие такой детерминации было показано в простом эксперименте. Если яйцо морского ежа искусственно разделить надвое, способность каждой половины развиваться нормально зависит от плоскости, вдоль которой яйцо было рассечено. Например, яйцеклетки одного из видов средиземноморских морских ежей, Раrаcentrotus lividus, имеют пигментированную полосу, которая разделяет яйцеклетку поровну при первом и втором естественно протекающем дроблении (рис. 5-1). Если одну из таких яйцеклеток искусственно рассечь вдоль одной из первых двух плоскостей дробления, то обе части (каждая из них содержит ядро) будут развиваться нормально. Если же яйцо рассечь параллельно пигментной полосе, каждая половина может продолжать делиться, но обе разовьются в аномальные эмбрионы разных типов. Верхнее полушарие рассеченного яйца разовьется в реснитчатую бластулу без каких-либо признаков энтодермы и первичной кишки; нижнее полушарие формирует в основном энтодермальные клетки. Эксперименты такого рода с оплодотворенными и неоплодотворенными яйцеклетками показывают, что цитоплазма яйца морских ежей не однородна, она обладает полярностью еще до деления клетки. Двум полюсам яйцеклетки были даны названия анимальный и вегетативный. Хотя до настоящего времени и не получено точных доказательств, можно предположить, что цитоплазма в области каждого из полюсов обладает различными химическими свойствами. В частности, некоторые химические вещества, имитирующие те или иные полярные субстанции, могут способствовать развитию яйцеклетки морского ежа в "анимальную" или "вегетативную" личинку.
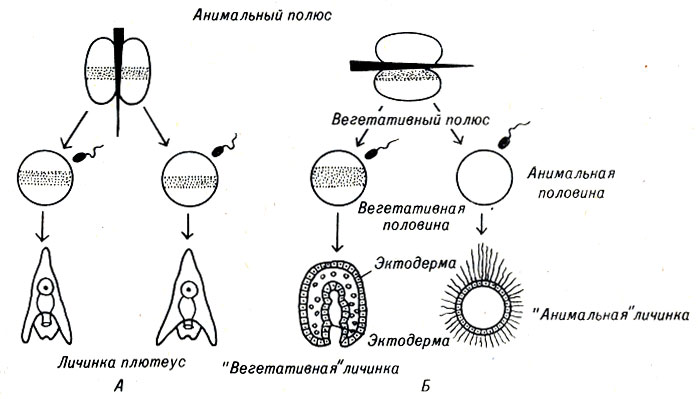
Рис. 5-1. Экспериментальное разделение неоплодотворенных яйцеклеток морских ежей при последующем их оплодотворении приводит к (А) нормальному развитию только в том случае, если пигментированная зона разделена пополам. В противном случае (Б) образуются дефектные 'анимальные' и 'вегетативные' личинки
С течением времени нормально развивающиеся зародыши достигают стадий эмбриогенеза, на которых "принимается решение", т.е. происходит детерминация, будет ли данная часть головой, а другая - хвостом или одна часть - животом, а противоположная - спиной. Другими словами, определяются оси симметрии: переднезадняя и спинно-брюшная. Кроме того, определяется, из каких клеток эмбриона будут развиваться отдельные органы. В большинстве случаев этапы, на которых происходят основные детерминации, установлены, описаны и экспериментально изучены. Однако причины, обусловливающие детерминацию на конкретных стадиях развития, остаются пока неизвестными.
5.4. Морфогенез - становление формы эмбриона - осуществляется в результате разнообразных процессов, протекающих внутри клеток, а также в результате взаимодействия клеток между собой
Процессы развития более динамичны, чем может показаться с первого взгляда. Если наблюдать развитие зиготы моллюска или морского ежа на последовательных кадрах кинопленки, можно убедиться, что оно представляет собой процесс, который почти постоянно испытывает разного рода воздействия, вызывающие сжатие, инвагинацию или изменения всей формы эмбриона. Такие формообразующие изменения называют морфогенезом.
Дробление - это морфогенетический процесс, при котором происходит деление яйца на две клетки, а в дальнейшем прогрессивное увеличение их числа. Образующаяся бластула может состоять из одного или нескольких слоев клеток, и обычно представляет собой полое образование. В определенное время в процессе клеточных делений, приводящих к образованию бластулы, некоторые клетки эмбрионов животных изменяют свою форму и приобретают способность к амебоидному движению, что также является морфогенетическим процессом.
Целые пласты клеток также могут претерпевать изменения формы и прочности сцепления друг с другом (адгезии). В этом случае клетки, расположенные внутри таких агрегатов, углубляются в пласт или покидают его, образуя складку, двойной слой клеток с полостью между ними, инвагинацию (углубление) или эвагинацию (выпячивание).
Примером наиболее ранней инвагинации является процесс, происходящий при гаструляции. Колбовидные клетки бластулы, которые первыми инвагинируют у дорсального края бластопора яйца амфибий, образуют примитивную, или первичную, кишку. Два слоя клеток, которые в этом случае формируются у эмбриона, приводят к появлению наружного слоя - эктодермы и внутреннего - энтодермы. У всех животных, кроме большинства примитивных, сходные клеточные процессы обусловливают формирование третьего слоя - мезодермы, расположенного между двумя другими.
5.5. У всех позвоночных, включая человека, формирование органов сопровождается последовательными специфическими тканевыми взаимодействиями первичных зародышевых слоев клеток эмбриона
Эктодерма, энтодерма и мезодерма - три первичных зародышевых слоя клеток эмбрионов животных - закладываются в самом начале гаструляции и сразу же вступают в.о взаимодействие друг с другом.
По мере того как клетки каждого из этих зародышевых слоев формируют органы, их жизнедеятельность контролируется не только собственными генами. Вступая во взаимодействие с рядом расположенными клетками, они испытывают влияние генов, считывающих в этих клетках другие участки генома. Тканевые взаимодействия и эмбриональная индукция приводят к тому, что одна группа клеток индуцирует развитие другой в определенном направлении. Развитие глаза позвоночных является примером, во-первых, эмбриональной индукции и, во-вторых, другого морфогенетического процесса, завершающегося образованием складок или изгибов клеточных слоев. Глаза в самом начале формирования представляют собой два симметрично эвагинирующих участка мозга, называемых глазными пузырями, которые вытягиваются и вступают в контакт с покрывающей их головной эктодермой. Как только произошла индукция, в обоих участках эктодермы наблюдается инвагинация клеток, образуется глазной пузырь, формирующий хрусталиковый пузырек и глазную чашу (рис. 5-2). В свою очередь, эти две структуры индуцируют покровный эпителий к образованию роговицы глаза, а окружающую мезодерму - к формированию защитных покровов глазного яблока. В том случае, когда глазной пузырь по каким-либо причинам не вступает в контакт с эктодермой, образования хрусталика и роговицы не происходит. Если в процессе формирования таза не происходит предусмотренного на определенном этапе клеточного взаимодействия, следующие этапы эмбриогенеза не наступают - таков основной принцип развития.
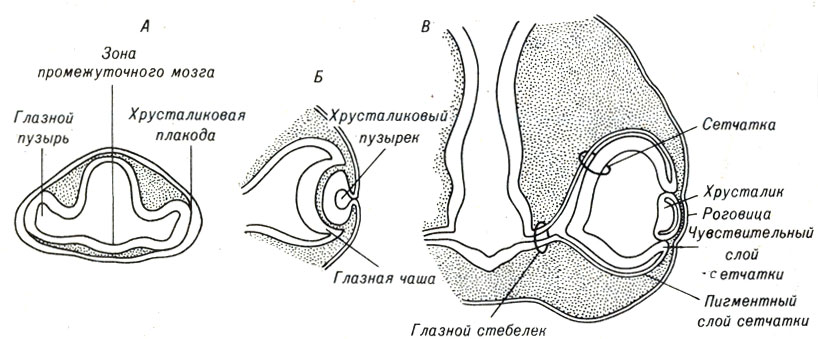
Рис. 5-2. Стадии формирования глаза у позвоночных, показанные на срезах головной части эмбриона
5.6. Клеточные взаимодействия могут осуществляться на расстоянии. Гормоны играют важную роль в развитии растений и животных
Гормон представляет собой вырабатываемое клеткой (или группой клеток) обычно в минимальном количестве вещество, которое влияет на расположенные вдали от нее восприимчивые клетки, так называемые клетки-мишени. Гормоны, таким образом, можно назвать химическими переносчиками информации. Различают гормоны развития и регуляторные гормоны. Первые участвуют в развитии растений и животных, вторые регулируют метаболические процессы. Некоторые гормоны могут первоначально действовать как гормоны развития на одном этапе жизни организма и как регуляторные - на другом. Чтобы проявилось действие гормонов в развитии, по-видимому, требуется активность специфических генов.
Ввиду того что у растений нет нервной системы, координация их жизнедеятельности всецело зависит от гормонов. Главным гормоном роста у растений является ауксин, названный так Вентом (F. W. Went), показавшим в 1930 г., что это вещество вызывает вытягивание (элонгацию) клеток. Автор помещал экстракты развивающихся растений в небольшие блоки агара и .прижимал каждый из них одной стороной к колеоптилю овса (лист, окружающий росток). Активность ауксина проявлялась, когда проростки овса отклонялись от блоков геля, поскольку, проникая в клетки на определенной стороне колеоптиля, ауксин вызывал их элонгацию. Позднее было обнаружено, что наиболее распространенным ауксином является индолуксусная кислота (ИУК), хотя активность ауксина могут проявлять и некоторые другие вещества.
Ауксины регулируют соотношение процессов роста у растений таким образом, что длина корней и ветвей находится в точной пропорции с их толщиной, а развитие кроны в целом сбалансировано с корневой системой. И наконец, ауксины индуцируют митозы в периферических участках зачатков корней. Вместе с другим гормоном - этиленом - они также регулируют процесс опадания, сбрасывания листьев. Этилен - летучий газ, который синтезируется в минимальном количестве плодами многих растений и является важным фактором их созревания. Кроме того, этилен может тормозить рост растений, в частности томатов.
Гормон растений цитокинин стимулирует клеточные деления в меристеме. Действуя совместно с ауксином, цитокинин оказывает разнообразные влияния на рост растений - в зависимости от количественного соотношения этих гормонов.
Значительно более направленное влияние на рост и развитие растений оказывают гиббереллины - вещества, впервые обнаруженные японскими исследователями у грибов Gibberella, которые вызывают заболевание саженцев риса. Гиббереллины синтезируются в активно растущих зонах растений и затем перемещаются в другие его участки. Эти гормоны активны в крайне малых количествах и часто оказывают значительное воздействие на организм растений. Например, при добавлении нескольких микрограммов гиббереллинов некоторые карликовые мутанты пшеницы вырастают до нормальной высоты. Данные гормоны также играют важную роль на самых ранних этапах развития ряда растений, например зерновых. Зародыши растений выделяют гиббереллин, который, проникая через эндосперм к наружному слою семени, индуцирует синтез ферментов, расщепляющих питательные вещества в эндосперме. Продуктами этого расщепления питается развивающийся зародыш.
Гормоны играют важную роль также в развитии большинства животных. Сравнивая сложные изменения, происходящие в эмбриогенезе млекопитающих (формирование глаза, уха), можно убедиться, что еще более загадочные изменения наблюдаются в процессе превращения головастика в лягушку, гусеницы в мотылька или личинки омара во взрослую особь. Процесс коренного изменения формы, свойственный амфибиям, насекомым и многим морским беспозвоночным , называют метаморфозом. Обычно он регулируется гормонами.
Очень подробно был изучен метаморфоз у бабочек тутового шелкопряда Cecropia. Установлено, что в осуществлении этого процесса принимают участие три гормона. Вначале из отложенных яичек выводится небольшая личинка (гусеница), которая начинает активно питаться и расти. Затем пара желез задней части мозга выделяет ювенильный гормон (общий для всех насекомых), который, действуя определенным образом, поддерживает личинку в ювенильном состоянии. Обратное действие оказывают два других гормона (антагонисты ювенильного). В частности, некоторые клетки мозга выделяют гормон, стимулирующий образование в загрудных железах гормональной смеси, - экдизон. Действие экдизона стимулирует линьку (сбрасывание экзоскелета) и метаморфоз . Однако на ранних стадиях существования в организме гусеницы содержится высокий уровень ювенильного. гормона, и поэтому в таких условиях действие экдизона ограничивается только процессом линьки. Позднее, когда уровень ювенильного гормона снижается, эффекты действия экдизона становятся бол ее выраженными, гусеница окукливается (формирует твердую оболочку) и в конечном итоге выходит из кокона в виде взрослой бабочки (рис. 5-3).
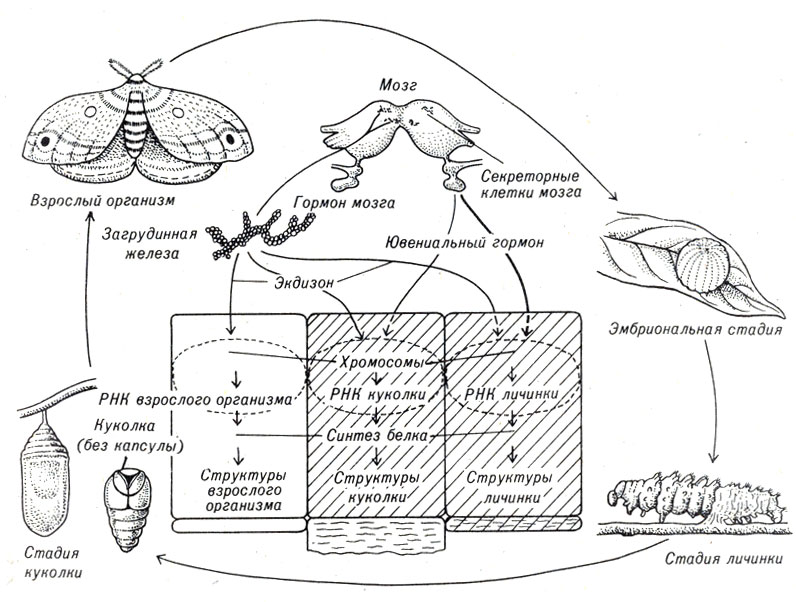
Рис. 5-3. Гормональный контроль метаморфоза у тутового шелкопряда (Cecropia)
Другой пример метаморфоза - превращение головастика в лягушку. Клетки гипофиза выделяют гормон, который стимулирует в щитовидной железе образование тиреоидного гормона - тироксина, регулирующего метаморфоз. Это приводит к регрессивным изменениям в некоторых тканях и органах - втягиванию жабер, сбрасыванию хвоста, выпадению зубов, а также к конструктивным изменениям - развитию конечностей и формированию всех признаков взрослого организма. Если головастику имплантировать небольшой кусочек щитовидной железы или ввести чистый тироксин, - начнется метаморфоз, который будет протекать нормальным путем.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'