4. Половое размножение и развитие животных
Ключевые вопросы
Каковы две основные функции половых желез?
Как влияет смена дня и ночи на цикл размножения?
В чем состоят основные отличия в строении сперматозоида и яйцеклетки?
Какие превращения происходят в сперматозоиде во время его контакта с яйцеклеткой?
Как реагирует яйцеклетка на прикрепление сперматозоида? Какие процессы в ней затем происходят?
Какое влияние на процесс формирования бластулы оказывает величина желтка?
Чем отличаются процессы гаструляции у зародышей морских ежей, лягушек, кур и млекопитающих?
Что такое внезародышевые оболочки?
Зависит ли строение тела эмбриона позвоночных от взаимного расположения трех первичных зародышевых листков, которые образуются после гаструляции?
Какова роль трех первичных зародышевых листков в формировании основных органов тела?
Животные, подобно растениям, адаптировались практически к любым условиям окружающей среды, в результате чего в процессе эволюции их размножение приняло многообразные формы. В данной главе более подробно будет рассмотрено развитие животных от зиготы до взрослого организма.
4.1. В гонадах (половых железах) вырабатываются половые гормоны и происходит развитие гамет (половых клеток) животных
Часто, на самых ранних этапах развития у подавляющего большинства животных будущие половые, или зародышевые, клетки обособляются от обычных, или соматических, клеток тела. Позднее формируются гонады (половые железы), в которых эти зародышевые клетки развиваются. Половые железы у самок называются яичниками, у самцов - семенниками. Именно в них образуются зрелые гаметы яйцеклетки и сперматозоиды.
Половые железы у животных сильно различаются по степени сложности и постоянству функционирования. Обычно половая железа функционирует постоянно и служит в качестве эндокринной железы, где вырабатываются гормоны и развиваются гаметы. У некоторых животных (например, у оболочников и примитивных хордовых) половые железы функционируют только в определенные сезоны или в особых случаях могут полностью исчезать на время между периодами размножения.
У большинства животных половые железы существуют в течение всей жизни, но их активность подвержена сезонным колебаниям. (Приматы - обезьяны, человекообразные обезьяны и человек - являются исключением из этого правила, поскольку они способны размножаться в течение всего года.) У ряда животных активность размножения зависит от длительности светового периода суток, фотопериода. В частности, при клеточном содержании птиц было обнаружено, что цикл их размножения можно регулировать искусственно, изменяя продолжительность дня. Животные с таким типом регуляции размножения адаптируются к определенному сезону, поэтому потомство у них появляется в наиболее благоприятное время. Так, размножение у быстро развивающихся животных, в частности у птиц и мелких млекопитающих, происходит весной или в начале лета. Молодняк, родившийся, в это время, способен перенести последующую зиму. У более крупных животных, таких, как олени, брачный период приходится на осень. В течение последующих нескольких месяцев зародыш растет и развивается в матке.
Роды происходят ранней весной. За летний период потомство вырастает и приобретает все свойства, необходимые для самостоятельного существования зимой.
4.2. Яичник является женской половой железой, в нем зародышевые клетки образуют яйцо, или яйцеклетку
Зрелые яйцеклетки отличаются от всех других клеток самки. Они имеют гаплоидное ядро, активную цитоплазму и запасы питательных веществ, различные у разных видов, которые принято называть желтком. Желток содержит жиры, белки и углеводы; он обеспечивает зародыш энергией и веществами, необходимыми для развития.
Яйцеклетки имеют разные размеры. У некоторых медуз диаметр яйцеклеток составляет всего 30 мкм, в то же время объем яйца у страуса почти 2 л. (В Африке бушмены используют скорлупу яиц страусов для хранения воды.) Яйцеклетки разных видов животных отличаются также по расположению желтка (рис. 4-1). У морского ежа, например, желток и активная цитоплазма распределены в яйцеклетке почти равномерно. В яйцеклетках лягушки ядро и основная часть активной цитоплазмы находятся в верхнем анимальном полушарии; более тяжелый желток располагается в нижнем - вегетативном полушарии. Следует отметить, что характер расположения желтка, по мнению некоторых исследователей, играет незначительную роль или вообще не оказывает влияния на развитие взрослого организма.
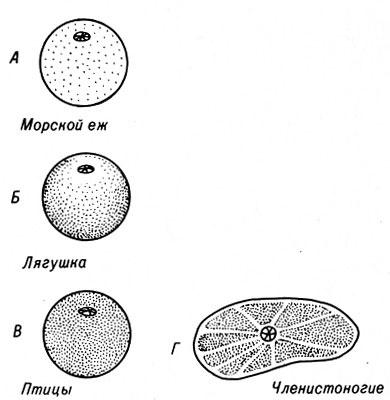
Рис. 4-1. Расположение желтка в яйцеклетках различных видов животных. А. Изолецитальное (желток расположен равномерно). Бив. Телолецитальное (желток расположен у одного из полюсов). Г. Центролецитальное (масса желтка расположена в центре и окружена тонким слоем прозрачной цитоплазмы). Приблизительные размеры яйцеклеток:Л. 0,1 мм, Б. 2 мм, 8. 20 мм, Г. 0,5 мм
4.3. Мужская половая железа, или семенник, - место образования сперматозоидов
Структура сперматозоидов у животных настолько разнообразна, что различие в их размерах и форме можно использовать для идентификации видов (рис. 4-2). Большинство сперматозоидов обладают подвижностью, которая обычно обеспечивается жгутиком. Гаплоидное ядро сперматозоида в большинстве случаев находится в его утолщенной головке.
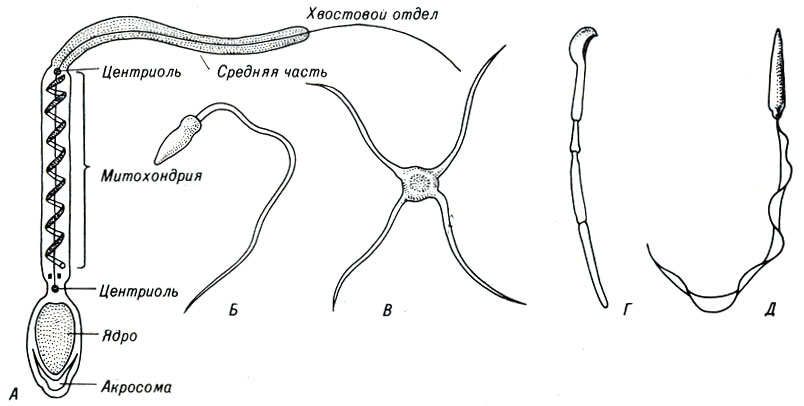
Рис. 4-2. Некоторые типы сперматозоидов и строение сперматозоида млекопитающих. Сперматозоиды: А - млекопитающих, Б - морского ежа, В - рака, Г - морской свинки, Д - жабы
Почти у всех видов животных в самой верхней части головки сперматозоида имеется небольшое образование, акросома, в котором находятся ферменты, играющие важную роль при слиянии яйцеклетки и сперматозоида.
4.4. Оплодотворение - сложный процесс, при котором яйцеклетка и сперматозоид взаимодействуют друг с другом
В сперматозоидах некоторых животных при сближении с яйцеклеткой наблюдается акросомалъная реакция. Микронити некоторых акросом неизвестным пока образом быстро реорганизуются и образуют акросомную нить, прокалывающую яйцеклетку. В сперматозоидах других видов животных образуются ферменты, которые способствуют проникновению сперматозоида через студенистую оболочку и другие покрывающие яйцеклетку слои.
В ответ на прикрепление сперматозоида в периферической цитоплазме яйцеклетки формируется конус оплодотворения (рис. 4-3), который увеличивается в направлении головки сперматозоида и поглощает его.
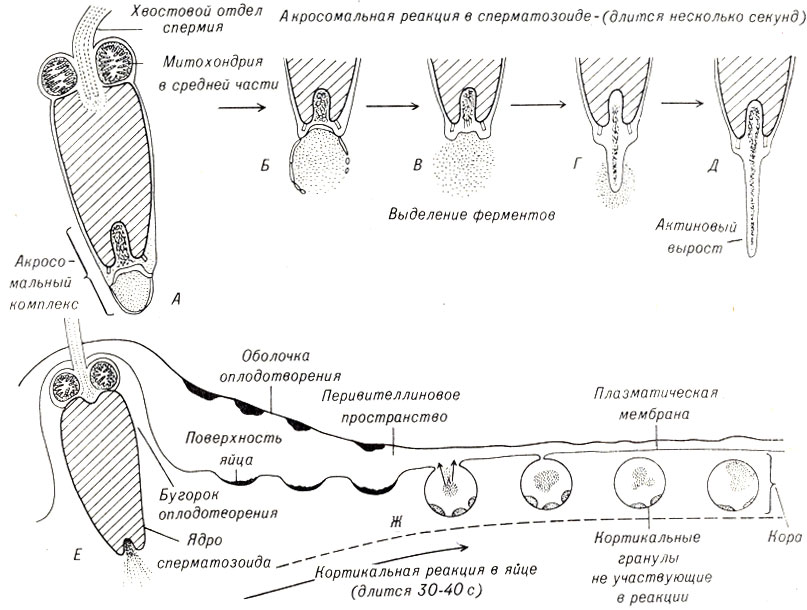
Рис. 4-3. Взаимодействие сперматозоида и яйца морского ежа в процессе оплодотворения; вверху - акросомальная реакция сперматозоида; внизу-кортикальная реакция, приводящая к формированию оболочки оплодотворения
Поверхностный слой яйцеклетки отвечает на активирующее влияние сперматозоида рядом различных структурных и химических изменений, названных кортикальной реакцией.
Через несколько минут после проникновения сперматозоида происходят самые разнообразные изменения в строении и функционировании яйцеклетки.
Скорость поглощения яйцеклеткой кислорода и жидкостей в это время обычно изменяется (увеличивается у одних видов, уменьшается у других). У некоторых видов над поверхностью яйцеклетки появляется оболочка оплодотворения и формируется защитная капсула. Яйцеклетки большинства видов животных утрачивают способность к оплодотворению дополнительным сперматозоидом.
Следует принять во внимание, что при оплодотворении сперматозоид играет двоякую роль: во-первых, он активирует развитие яйцеклетки; во-вторых, вносит свой гаплоидный набор хромосом, необходимый для образования диплоидного организма. Активация развития яйцеклетки происходит не сразу после проникновения сперматозоида. Развитие не начинается еще некоторое время, пока интактный сперматозоид находится в цитоплазме. Вероятно, в это время более важны для развития события, происходящие на поверхности яйцеклетки, - активация.
Слияние сперматозоидов и яйцеклеток происходит различными способами, которые распадаются на две основные категории - внешнее оплодотворение, когда сливаются гаметы водных животных, и внутреннее оплодотворение, когда при спаривании сперматозоиды вносятся в тело самки.
4.5. После оплодотворения происходит дробление одноклеточного яйца и формирование популяции клеток, называемой бластулой
Процесс клеточного деления, происходящий на ранних стадиях эмбрионального развития (дробление), по существу сходен с делением клеток у взрослого организма. За митозом следует цитокинез. Однако дробление отличается тем, что дочерние клетки становятся более мелкими, чем родительские, а скорость деления на ранних эмбриональных стадиях значительно выше (иногда всего 15 мин между дроблениями).
Процесс цитокинеза отличается у эмбрионов различных видов. Наиболее важным фактором, определяющим различия в цитокинезе, является характер распределения желтка.
Яйцеклетки морского ежа имеют небольшое количество желтка, который равномерно распределен в яйце. Ядро расположено близко к центру, поэтому в результате двух первых делений образуются клетки одинакового объема (рис. 4-4). Эти клетки, или бластомеры, делятся синхронно. После четвертого деления образуется группа из четырех клеток, микромеров, значительно меньших по величине, чем другие 12 клеток. Микромеры не содержат желтка, и их свойства в большей степени связаны с поверхностными свойствами яйца. Участки поверхности каждого бластомера, обращенные в направлении оболочки яйца, образуют полый шар из одного слоя клеток - бластулу. Внутренняя полость бластулы, заполненная жидкостью, носит название бластоцель.
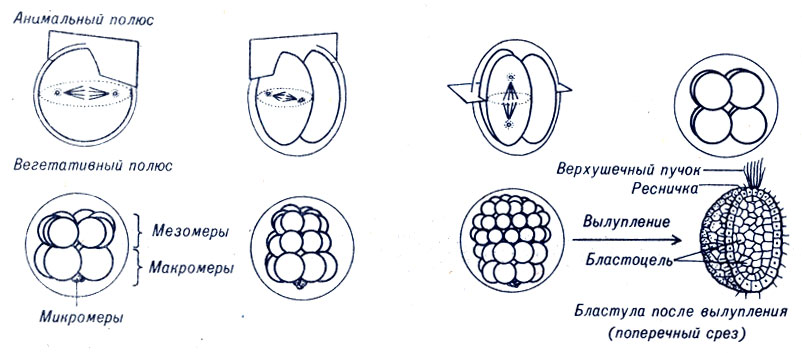
Рис. 4-4. Дробление яйца морского ежа на 2, 4, 8, 16, 32 и 64 клетки и реснитчатая бластула. Показаны плоскости дробления первых трех делений
Земноводные (например, лягушки и саламандры) образуют яйцеклетки с большими запасами желтка, чем морские ежи. Желток концентрируется у вегетативного полюса яйцеклетки, ядро же располагается ближе к анимальному полюсу. Первая борозда дробления постепенно делит яйцо вдоль анимально-вегетативной оси. Второе деление начинается до того, как первая борозда разделит яйцо на две полностью обособленные клетки. Борозды третьего дробления располагаются под прямым углом к первым двум и находятся вблизи экватора. В результате этого дробления анимальный полюс оказывается наверху, а вегетативный - внизу (рис. 4-5). Последующие дробления методично делят вещество яйца на более мелкие части, хотя скорость делений замедляется в вегетативной зоне. Очевидно, процессу формирования борозд в вегетативной зоне может препятствовать большое количество желтка. Какдв бы ни был механизм этого процесса, замедление скорости деления клеток вегетативной зоны приводит к постепенному увеличению их размеров от анимального полюса к вегетативному. Параллельно образуется наполненная жидкостью бластоцель.
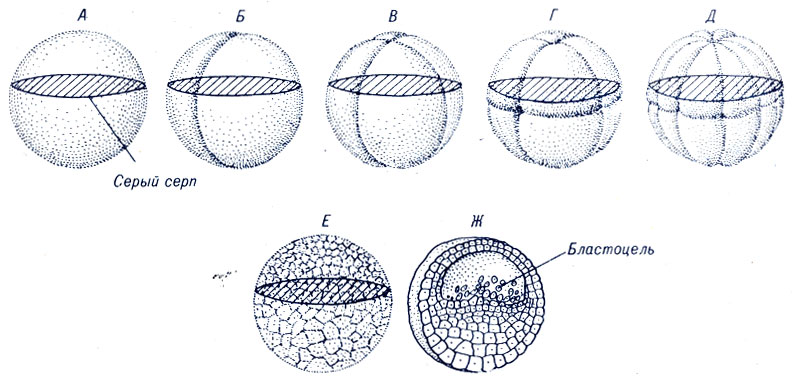
Рис. 4-5. Дробление яйца земноводных. После оплодотворения, напротив места проникновения сперматозоида, при перемещении пигмента к анимальному полюсу образуется серый серп
Цитоплазма яйцеклеток птиц представляет собой очень тонкий, не содержащий желтка зародышевый диск диаметром около миллиметра. Это белое пятнышко, обычно перемещающееся по поверхности желтка, если яйцо вылито на сковороду. В зародышевом диске происходит несколько делений до того, как яйцо отложено. Плоскость первых дроблений проходит под прямым углом к поверхности желтка. Создается впечатление, что развивающийся зародыш собирается существовать только в двух измерениях (рис. 4-6).
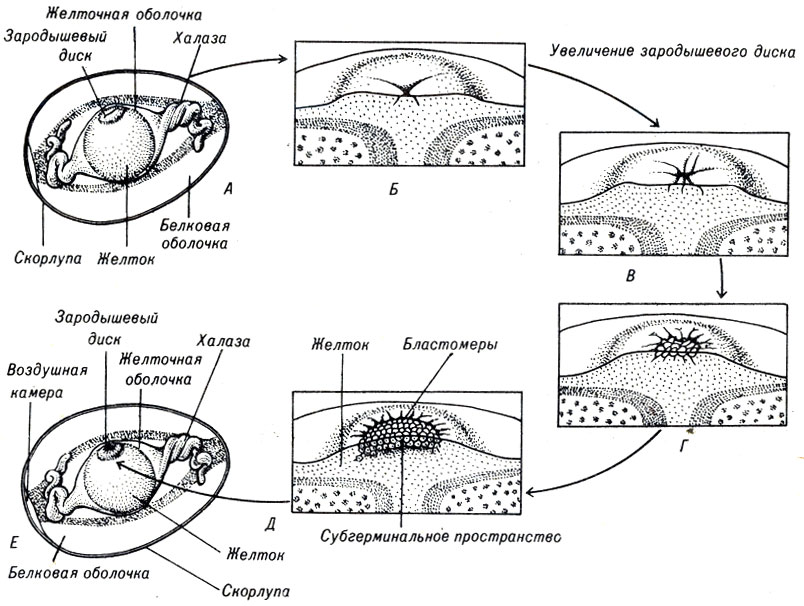
Рис. 4-6. Дробление зародышевого диска яйца птиц
При последующих дроблениях клетки становятся более мелкими, диск более утолщенным и состоящим из нескольких слоев клеток. Вскоре нижние клетки диска образуют свободнол ежащий слой - гипобласт, который отделен от верхнего слоя - эпибласта пространством, заполненным жидкостью. У птиц такое пространство является эквивалентом бластоцели (рис. 4-7). Густой, вязкий желток отодвигается от зародышевого диска, освобождая наполненное жидкостью субгерминальное пространство.
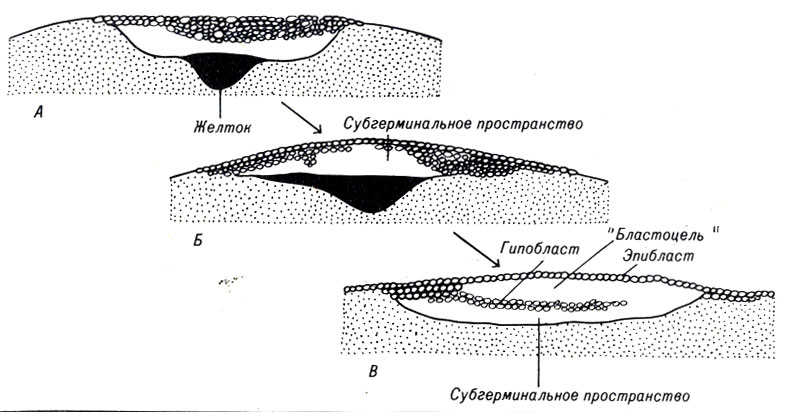
Рис. 4-7. Формирование первых двух эмбриональных клеточных слоев в яйце птиц
В яйцеклетках млекопитающих, так же как у морского ежа, содержится небольшое количество равномерно распределенного желтка. Поэтому при делении яйца образуется группа одинаковых по размеру клеток. Однако у человека и некоторых млекопитающих эти клетки очень быстрЬ разделяются на две группы. Одна становится источником клеток эмбриона, из другой же образуются клетки внеэмбриональных оболочек и плаценты, необходимых для существования зародыша в организме матери.
У млекопитающих оплодотворение происходит около устья яйцевода во время выхода каждой яйцеклетки в матку. Зародыши развиваются в процессе передвижения вниз по просвету матки. Внутри сформировавшегося в результате дробления плотного комка клеток шаровидной формы появляется заполненное жидкостью пространство, которое увеличивается и образует относительно большую полость. На этой стадии зародыш носит название бластоцисты. Клетки, из которых в дальнейшем должен развиваться эмбрион, образуют утолщение стенки бластоцисты, называемое внутренней клеточной массой. Остальные клетки бластоцисты составляют зону трофобласта, из которого образуются внеэмбриональные оболочки и плацента.
После того как бластоциста имплантируется в стенку матки, основные изменения происходят во внутренней клеточной массе. Между наружным слоем бластоцисты и внутренней клеточной массой формируется амниотическая полость. Клетки внутренней массы, примыкающие к полости бластоцисты, растут параллельно внутренней поверхности трофобласта, образуя новый слой - желточный мешок. Другая часть клеток внутренней клеточной массы составляет диск, отделяющий амниотическую полость от увеличивающегося желточного мешка. Этот диск - зародышевый щиток. В дальнейшем только из этой части развивается эмбрион (рис. 4-8).
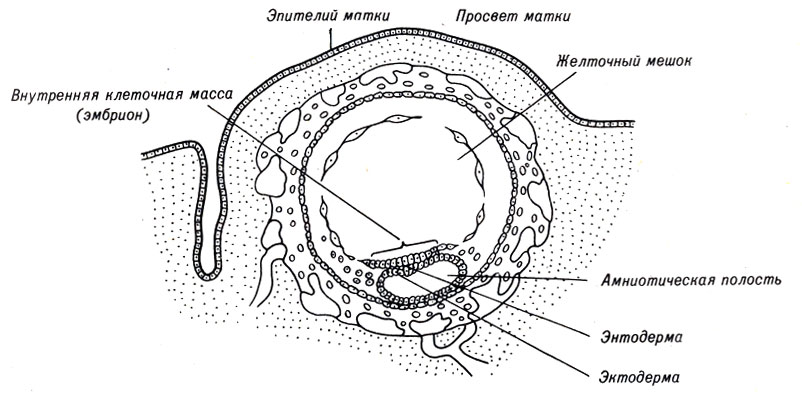
Рис. 4-8. Схема строения 12-дневного эмбриона млекопитающих, имплантированного в стенке матки
До сих пор развитие у рассмотренных нами животных было относительно простым, т. е. деление клеток приводило к формированию бластулы или подобной ей структуры, тем самым наступала стадия развития, после которой последуют более сложные процессы.
4.6. Гаструляция представляет собой процесс, в результате которого у эмбриона формируется первичная кишка, а затем три зародышевых листка
Строение большинства многоклеточных животных подобно конструкции типа "трубка внутри трубки" (первичная кишка, окруженная наружным слоем клеток эмбриона). Кишечная трубка может быть открыта с обоих концов или только с одного. В любом случае животное может захватывать пищу, переваривать ее и удалять нерастворимые остатки либо через то же отверстие (как у Hydra), либо, как у более сложных организмов, через другое, анальное отверстие.
Хотя пищеварительный тракт не используется зародышем до самого окончания эмбрионального развития, раннее формирование этой структуры крайне важно для установления будущей симметрии организма животного. Одновременно с развитием первичной кишки происходит образование у эмбриона нескольких слоев клеток, или, другими словами, начинается процесс гаструляции. На этой стадии развития эмбрион носит название гаструла.
Наиболее простым способом преобразования сферической формы зародыша в конструкцию типа "трубка внутри трубки" является втягивание части его поверхности внутрь, по мере того как вся структура увеличивается. У многих животных, в частности у морского ежа, имеющего крупную бластоцель, формирование первичной кишки, или архентерона, очень сходно с описанным выше процессом (рис. 4-9). Во время гаструляции деление клеток практически отсутствует. Процесс гаструляции осуществляется частично благодаря независимой активности перемещающихся внутрь клеток.
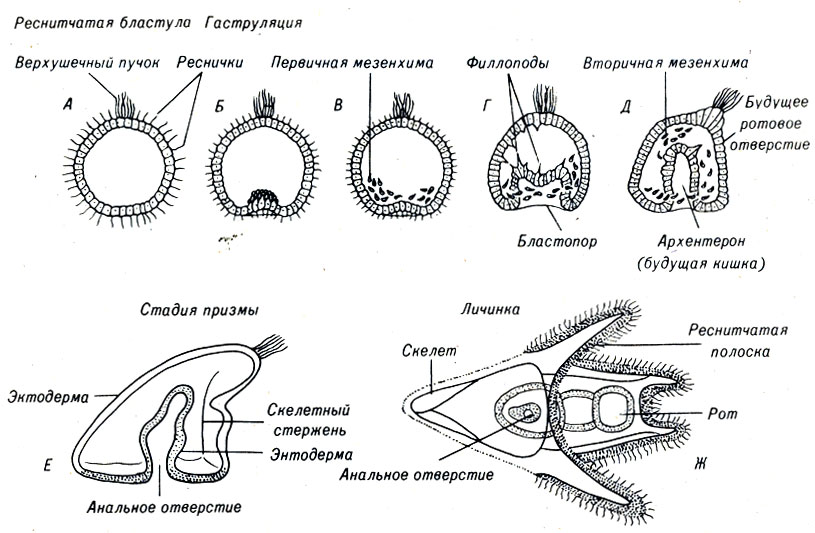
Рис. 4-9. Развитие морского ежа от стадии гаструлы до личинки
По мере развития первичной кишки у эмбриона морского ежа образуются длинные, тонкие ложноножки - филлоподии, соединяющие конец архентерона со стенкой анимального полюса. Укорачиваясь, они могут создавать дополнительные усилия, необходимые для гаструляции.
При формировании архентерона не только устанавливается основная ось эмбриона, но также происходит разделение его на внутренний и наружный слои. Затем между ними образуется третий слой, состоящий из свободно перемещающихся около бластопора клеток (рис. 4-9). Пласт клеток, образующий внутренние стенки первичной кишки, называют энтодермой; наружный слой клеток обозначается как эктодерма, а группу клеток, которая располагается между эктодермой и энтодермой, называют мезодермой. Эти три клеточных слоя обычно называют зародышевыми листками.
Каждый зародышевый листок образует определенные структуры, которые одинаковы у подавляющего числа животных, хотя у некоторых видов из тех же зародышевых листков образуются другие структуры. Тем не менее внутренний слой клеток кишечника, некоторых желез и внутренних органов развивается из энтодермы; кровь и кровеносная система, мышцы, скелет и другие опорные ткани образуются из мезодермы; в свою очередь нервная система, кожа и другие покровные ткани формируются из эктодермы.
Таким образом, гаструляция у морского ежа, устанавливающая главную ось тела, типична для многих животных. Незамкнутая первичная кишка представляет собой этап гаструляции, и его называют бластопором. В отличие от других животных у морского ежа бластопор сохраняется всю жизнь, выполняя функцию анального отверстия. В том месте, где передний конец архентерона соприкасается со стенкой зародыша, возникает ротовое отверстие (рис. 4-9). В течение нескольких дней происходит развитие яйца морского ежа до стадии личинки. Последующее развитие до взрослого состояния занимает намного большее время и включает в себя значительные изменения формы тела - метаморфоз.
Гаструла лягушек и аксолотлей в общих чертах сходна с аналогичной стадией зародышевого развития морского ежа, однако существует и много отличий в прохождении отдельных этапов гаструляции. У земноводных клетки, расположенные ниже экватора яйца, перемещаются внутрь, образуя на поверхности зародыша борозду или складку (рис. 4-10). Процесс гаструляции начинается с участка, содержащего непигментированную цитоплазму. Этот участок называют серый серп, он расположен несколько ниже экватора яйца, обычно напротив места проникновения сперматозоида. Позднее данная область яйца становится дорсальной (спинной) частью бластопора, которая располагается на задней поверхности зародыша и образует дорсальную губу. В это же время устанавливаются передняя, задняя, дорсальная, вентральная, правая и левая части эмбриона.
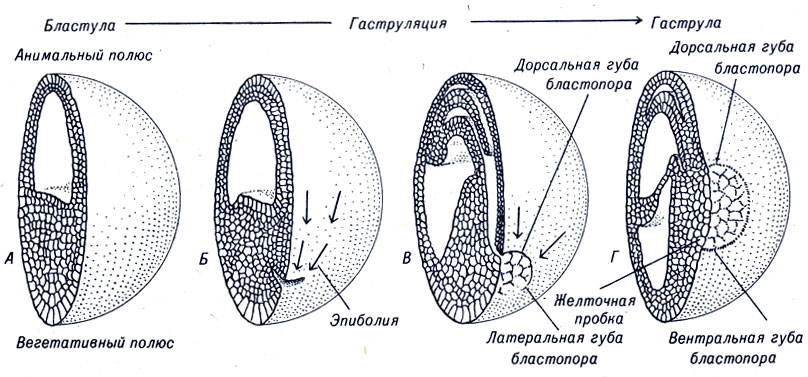
Рис. 4-10. Гаструляция яйца амфибий
Клетки, перемещающиеся внутрь, приобретают куполообразную форму, а поскольку гаструляция становится более интенсивной, этот купол быстро увеличивается и образует полость. Клетки, которые заворачиваются в бластопор, сохраняют связь как с втянутыми внутрь, так и с находящимися снаружи клетками. Гаструляция, таким образом, завершается формированием клеточных слоев.
К концу гаструляции у амфибий происходит существенная перегруппировка клеток зародыша. Наружный слой полностью сформировавшейся гаструлы - эктодерма - состоит из клеток, которые ранее покрывали около половины поверхности бластулы. Поэтому процесс гаструляции сопровождается значительным вытягиванием клеток. Клетки, которые прошли через дорсальную губу бластопора, образуют временный свод архентерона, а в будущем дадут начало энтодермальным клеткам, выстилающим стенки архентерона. Большинство клеток, которые втягиваются через губы бластопора, позднее входят в состав мезодермы.
Тем временем бластоцель перемещается вперед, сжимается и постепенно исчезает. У амфибий, как и у морских ежей, бластоцель служит местом, где клетки могут перемещаться в процессе гаструляции.
Несмотря на различия в содержании желтка, гаструла земноводных, по существу, сходна с такой же стадией развития морского ежа. Однако механизмы созревания яйцеклеток двух этих видов отличаются коренным образом.
У птиц, пресмыкающихся и некоторых млекопитающих поведение зародышевого диска в процессе гаструляции напоминает в общих чертах развитие бластулы у морского ежа и земноводных, хотя плоская структура зародышевого диска обусловливает и определенные различия. Напомним, что слой клеток, лежащий над бластоцелем, назван эпибластом, а клетки, ограничивающие полость снизу, - гипобластом (рис. 4-7). В процессе гаструляции гипобласт практически не перемещается и в дальнейшем образует энтодерму. Из эпибласта главным образом развиваются эктодерма и мезодерма. В процессе гаструляции у будущего заднего края бластодиска клеточные слои несколько утолщаются, а правая и левая половины эпибласта сближаются друг с другом, образуя срединную ось животного (рис. 4-11). Там, где соединяются два клеточных листка, возникает неглубокий желобок, заканчивающийся углублением, известным как гензеновский узелок. Эту похожую на желобок структуру называют первичной полоской, она представляет собой зону, где двигающиеся навстречу друг другу клетки встречаются, мигрируют внутрь и перемещаются к краям зародышевого диска между эпибластом и гипобластом.
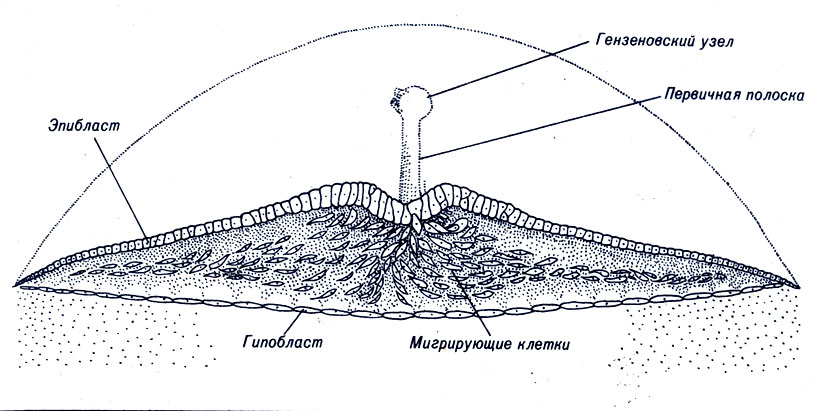
Рис. 4-11. Гаструляция яйца птиц
Большинство клеток, продвинувшихся внутрь от первичной полоски, остаются между эпибластом и гипобластом и образуют мезодермальный слой. Некоторые клетки, расположенные внутри зародыша, соединяются с гипобластом, который в дальнейшем называют энтодермой. Оставшиеся на поверхности клетки зародышевого диска составляют эктодерму. Первичная полоска, таким образом, эквивалентна бластопору, а эмбрион развивается спереди от этой структуры.
Из данного описания видно, что развитие зародышевых листков происходит сходно. Однако у многих животных процесс гаструляции протекает различными путями, хотя и приводит к одинаковому конечному результату. Например, не всегда зародыши имеют бластоцель. В этом случае небольшие клетки, образовавшиеся из анимального полюса яйца, смещаются на поверхность крупных клеток вегетативной зоны. Первичная кишка может сформироваться из популяции клеток, передвигающихся внутрь эмбриона, или из плотной группы клеток, которые позднее образуют полость архентерона с отверстием на поверхности.
4.7. После гаструляции происходит дифференцировка тканей и развитие органов
Сразу после гаструляции можно предсказать, из каких клеток будут образованы мозг, кожные покровы и т. п. Подобные предсказания основываются главным образом на расположении клеток, а не на какой-либо характерной особенности их формы или химического состава.
После гаструляции зародышевые клетки быстро дифференцируются и из них формируются органы. Этот процесс называют органогенезом. Ниже будет рассмотрено, как протекает этот процесс у позвоночных. Следует иметь в виду, что, хотя данные о развитии отдельных органов приведены в последовательном изложении, в эмбрионе многие важные события происходят одновременно.
4.8. Центральная нервная система развивается из эктодермы
Участие эктодермы в развитии зародыша позвоночных начинается с формирования центральной нервной системы, в частности головного и спинного мозга, периферических нервов. У большинства позвоночных полость дорсального нервного ствола начинает формироваться из участка дорсальной эктодермы, образующей вытянутую пластинку, которая затем свертывается в трубку. У лягушек и аксолотлей это очень динамичный процесс, некоторые этапы которого можно наблюдать невооруженным глазом (рис. 4-12).
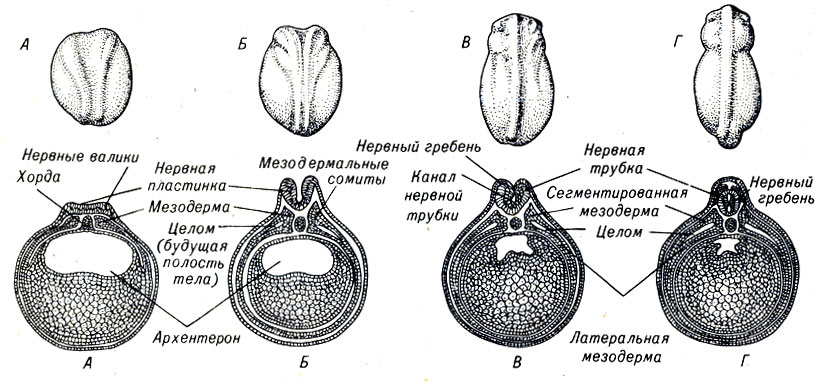
Рис. 4-12. Стадии нейруляции (образование нервной трубки), развитие хорды и мезодермы у зародыша амфибий: вверху - общий вид; внизу - поперечный срез зародыша
На будущей дорсальной стороне эмбриона сразу после того, как желточная пробка втягивается в бластопор, возникает утолщенная зона, ограниченная небольшими складками. Это образование называется нервной пластинкой и ее развитие приводит к формированию головного и спинного мозга. По мере того как складки (нервные валики) перемещаются к средней линии навстречу друг другу, нервная пластинка опускается и образует продольный желобок. Верхние края складок сдвигаются к центру, превращая пластинку в нервную трубку, покрытую клетками, находившимися ранее на внешней стороне гаструлы.
В процессе формирования нервной трубки клетки пластинки изменяют свою форму. Сразу после закрытия нервной трубки отдельные ее части расширяются с различной скоростью, в результате чего возникают вздутия, сужения и бугорки. Наиболее значительные вздутия - два глазных пузыря (будущие глаза), которые располагаются на переднем конце нервной трубки
4.9. Сердце развивается из мезодермы
Позвоночные имеют одно сердце, расположенное около средней линии тела в вентральной его части. Сердце формируется в результате слияния пары трубчатых структур, которые развиваются из зачатков мезодермы. В конце гаструляции эти структуры находятся под передними валиками нервной пластинки.
По мере смещения данных участков мезодермы к средней линии каждый из них образует тонкостенную трубку. При их объединении образуется общая трубка в передней части эмбриональной грудной клетки (рис. 4-13).
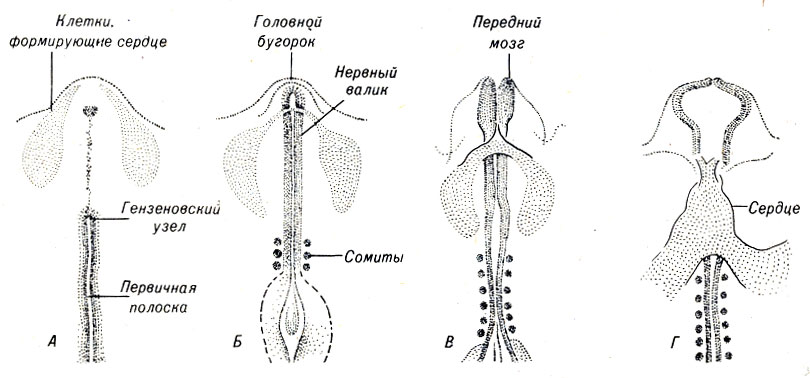
Рис. 4-13. Образование сердца куриного зародыша
Вскоре в этой структуре возникают сердечные сокращения, а затем формируются камеры сердца.
4.10. Хорда, сомиты и конечности развиваются из мезодермы
Из мезодермы формируется трубчатая опорная структура, называемая хордой, которая залегает под нервной трубкой. Данная структура является отличительной особенностью типа животных, который поэтому и называют Chordata.
У высших позвоночных опорную функцию позднее выполняет позвоночный столб.
Хорда представляет собой непрерывный тяж клеток, образующий блоки мезодермальной ткани - сомиты, из которых, в свою очередь, развиваются кости, хрящи, мышцы и слой дермы в коже (рис. 4-14).

Рис. 4-14. Зародыш птиц после 48 ч развития (слева). Показаны элементы нервной и кровеносной систем, а также парные сомиты (А). Справа - поперечные срезы эмбриона на уровне четырнадцатого (Б) и восемнадцатого (В) сомитов
В тех участках тела, где в будущем должны развиваться передние и задние конечности, появляются вздутия мезодермальной ткани - почки конечностей.
В процессе роста почки конечностей приобретают форму лопастей, в них формируются хрящи, дающие начало длинным трубчатым костям конечностей и мелким костям стопы, кисти.
Становление формы конечностей и пальцев частично связано с появлением локальных зон отмирания клеток (рис. 4-15).
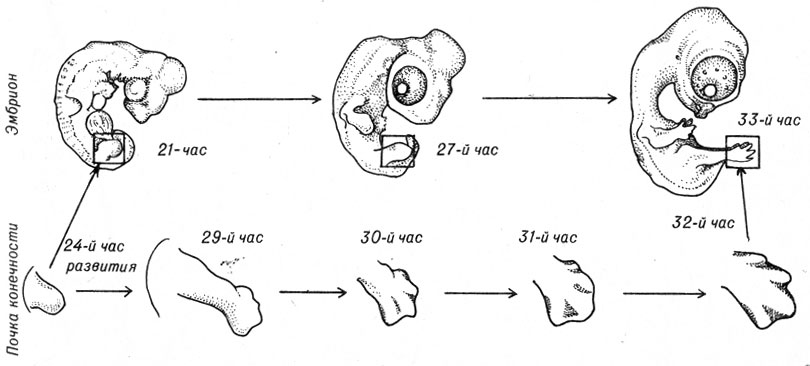
Рис. 4-15. Образование и развитие почки конечности. Показано влияние локальной гибели клеток на формирование пальцев. (Возраст эмбрионов указан в часах развития.)
4.11. Пищеварительный образуется из энтодермы тракт
У эмбрионов амфибий к концу гаструляции образуется первичная кишка, которая представляет собой полую трубку. У птиц и млекопитающих ее формирование происходит более сложным путем.
Первичная кишка недолго сохраняет форму простой трубки. В различных частях первичной кишки появляются выпячивания, из которых позднее развиваются дыхательная система: гортань, трахея, бронхи и язык и пищеварительные железы: печень, желчный пузырь, поджелудочная железа (рис. 4-16).
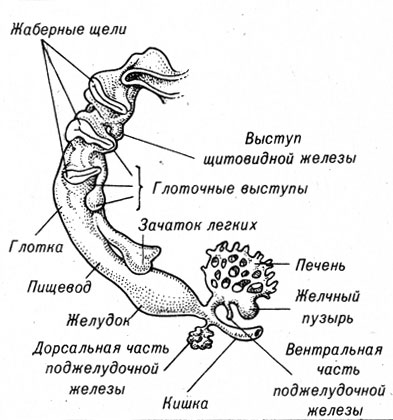
Рис. 4-16. Зачатки органов, развивающихся из первичной кишки млекопитающих
У эмбрионов всех позвоночных в передней части кишки развиваются жаберные дуги и жаберные щели. У рыб эти структуры сохраняются в течение всей жизни организма; у амфибий они функционируют до наступления метаморфоза, а у млекопитающих, включая человека, они являются только этапом в развитии отдельных частей гортани и среднего уха.
4.12. Развитие органов требует согласованного взаимодействия зародышевых листков и тканевых закладок
Из энтодермы образуется прежде всего внутренний слой первичной кишки, а из выпячиваний, которые на ней появляются, - внутренний слой дыхательных органов и пищеварительных желез. Мышечные стенки, соединительная ткань и кровь, относящиеся к висцеральным внутренним органам, берут начало от мезодермы. Большая часть нервной системы, которая формируется из эктодермы, развивается из нервной трубки, почти полностью состоящей из дифференцирующихся нервных клеток - нейробластов. По мере развития нервной системы происходит ее насыщение кровью и клетками соединительной ткани, образовавшимися из мезодермы. Мозг также развивается из отдельного участка нервной трубки, хотя его сосудистая сеть образуется из мезодермы.
В сущности, развитие любых органов позвоночных осуществляется в результате взаимодействия мезодермы и энтодермы либо мезодермы и эктодермы.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'