3. Половое размножение и развитие растений
Ключевые вопросы
Как происходят процессы митоза и мейоза при чередовании половой и бесполой фаз размножения растений?
Чем отличаются спорофитное и гаметофигное поколения водоросли Ulva (морской салат)?
К какому поколению - спорофитному или гаметофитному - относится лесной папоротник? Каким образом у него развивается следующее поколение?
Какие мужские и женские половые органы имеются в цветке?
Какие формы мужских и женских гаметофитных поколений существуют у цветковых растений?
Чем отличаются опыление и оплодотворение?
Каковы основные части зародыша цветковых растений?
Как изменяются питательные потребности у зародышей растений в процессе их развития?
Какую пользу растениям приносит способность их семян находиться в покоящемся состоянии?
Какие факторы способствуют их прорастанию?
Что такое меристемы? Где располагаются меристемные ткани у развивающихся растений?
Преимущества полового размножения так же существенны для растений, как и для животных, но, поскольку высшие растения стационарны, у них выработались иные способы объединения двух гамет для формирования нового организма.
После оплодотворения начинается развитие зародыша растения.
В течение очень короткого времени оплодотворенная яйцеклетка развива-естся в полностью сформированное растение.
3.1. У большинства растений половое и бесполое размножение попеременно сменяют друг друга
Цикл размножения почти всех видов растений включает половую и бесполую фазы. Обычно объединение двух гамет приводит к формированию диплоидной клетки - зиготы, которая развивается в диплоидное растение бесполого спорофитного поколения. Спорофитным растение называют потому, что в результате мейотического деления оно образует гаплоидные споры. В свою очередь, из каждой споры развивается гаплоидное растение полового гаметофитного поколения, которое образует гаплоидные гаметы. Цикл размножения повторяется снова после соединения двух гамет, образования зиготы и восстановления диплоидного набора хромосом. Споры никогда не объединяются друг с другом, каждая спора независимо делится митотически и развивается в целое растение.
Относительно простым примером чередования поколений является широко распространенная водоросль Ulva, или морской салат. (Водоросли включают в себя ряд простейших форм растений. Их размеры колеблются от единичных микроскопических клеток до многоклеточных образований со стеблями длиной около 75 футов, или 23 м.) Ulva растет в мелководных участках моря и имеет тонкие, почти прозрачные, зеленые листья.
Некоторые из этих растений являются диплоидными спорофитами, другие - гаплоидными гаметофитами.
В определенное время года некоторые клетки диплоидных спорофитов делятся и проходят мейоз внутри старых клеточных оболочек. Образовавшиеся гаплоидные споры разрывают окружающую их оболочку и распространяются в воде с помощью имеющихся у них четырех жгутиков (рис. 3-1). Подвижные споры выглядят одинаковыми и не проявляют тенденции к слиянию друг с другом.
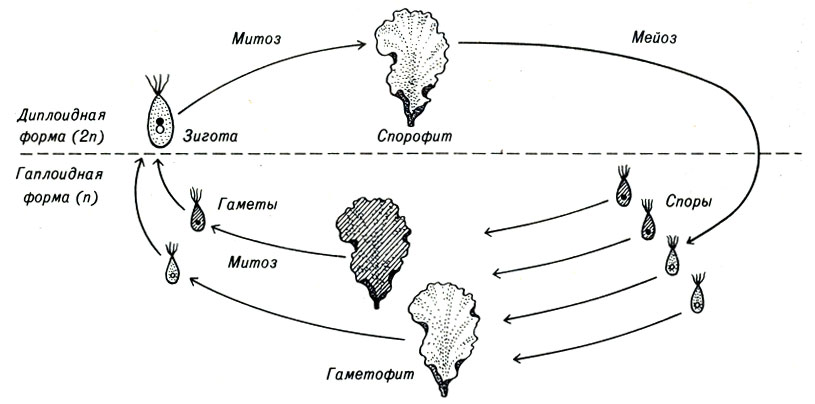
Рис. 3-1. Цикл развития Ulva (морского салата)
Если спора, опускаясь на дно, попадает на благоприятную почву, из нее развивается водоросль гаметофитного поколения, идентичная исходному спорофиту. Однако в ядрах клеток этого нового растения содержится гаплоидный набор хромосом. Со временем гаплоидные растения образуют изогаметы, каждая из которых имеет только два жгутика.
Если изогаметы двух разных растений объединяются, формируется зигота, которая может развиться в диплоидный спорофит.
3.2. У наземных растений чередование поколений более сложное, так как перемещение спор и гамет в воздухе более затруднено, чем в воде
Нецветущие растения, например лесные папоротники, являются диплоидными спорофитами. Споры образуются в результате мейоза в коричневых спорангиях на внутренних сторонах листьев папоротника. После того как созревшие споры падают на землю, они развиваются в маленькие, плоские, серцевидные гаметофиты (рис. 3-2). Гаметы образуются в специализированных структурах на нижней поверхности этих миниатюрных растений. Один тип гамет представляет собой маленькие подвижные микрогаметы (сперматозоиды). Другой - крупные неподвижные макрогаметы (яйцеклетки). У папоротников яйцеклетки и сперматозоиды образуются в одних и тех же гаметофитах.
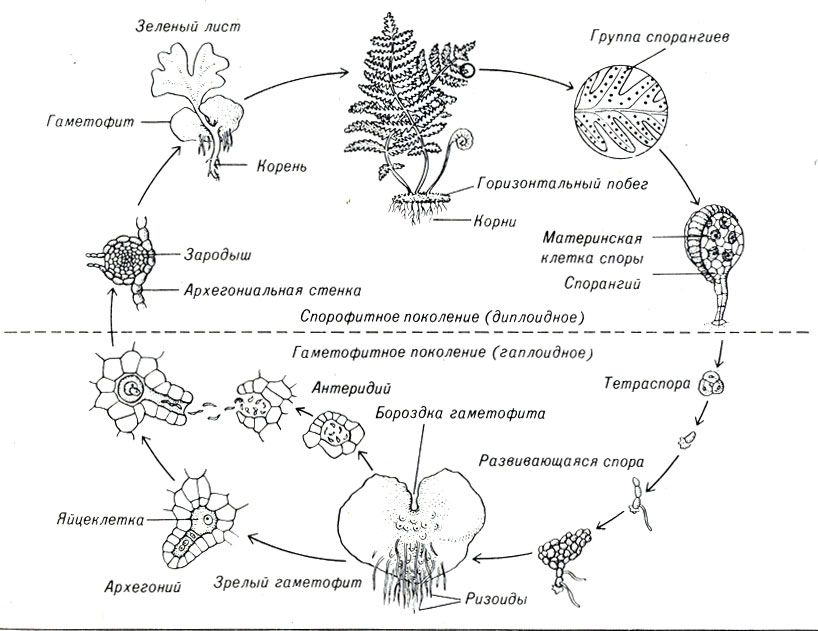
Рис. 3-2. Цикл развития папоротника
Когда происходит высвобождение сперматозоидов (во время дождливого периода), они передвигаются в воде, окружающей гаметофит. В свою очередь яйцеклетки образуют специфическое вещество, которое способствует передвижению сперматозоида к яйцеклетке и их слиянию. Образовавшиеся зиготы растут и разрушают гаметофит. Вновь сформированное растение является спорофитом.
3.3. У высших растений органы размножения находятся в цветках
Наземные растения часто подвержены воздействиям жары, холода, дождей, засухи, и, хотя взросльщ растения могут адаптироваться к таким условиям, большинство зигот не способно выжить. Поэтому у многих растений зиготы содержатся в тканях взрослого растения, которое их защищает. В этих условиях образуется меньшее количество гамет и зигот, однако каждая из них имеет больше шансов выжить.
Такими специализированными структурами, которые обеспечивают сохранность зародышей, являются цветки. Типичный цветок содержит все необходимые компоненты для полового размножения (рис. 3-3).
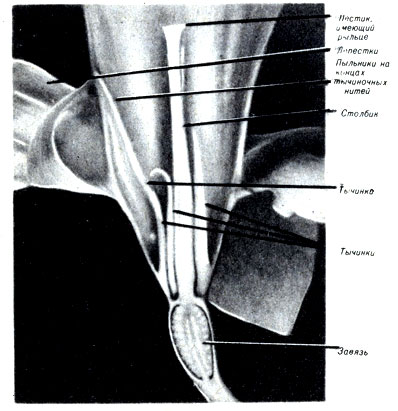
Рис. 3-3. Продольный разрез цветка нарцисса
Дополнительными частями цветка являются чашелистики, защищающие бутон во время его формирования, и лепестки, которые придают цветку особую привлекательность. Основными структурами размножения в цветке являются мужские тычинки, состоящие из тычиночных нитей, на концах которых расположены пыльники, и женский пестик, имеющий рыльце, столбики завязь.
Развитие микрогамет происходит в пыльниках. Макрогаметы формируются в завязи, где происходит оплодотворение и развитие зиготы.
У одних растений происходит самоопыление, т. е. микрогаметы из пыльника могут оплодотворять макрогаметы в завязи того же цветка. У других самоопылению препятствуют определенные физиологические барьеры. Властности, формирование микро- и макрогамет в период цветения происходит в различное время. У большой группы растений самоопыление невозможно, поскольку мужские b женские цветки образуются на разных растениях. У некоторых растений, размножающихся только вегетативно, развиваются стерильные цветы.
Независимо от того, происходит самоопыление или нет, гаметофитное поколение у цветущих растений представляет собой не отдельное растение, а часть цветка. Действительно, мужской гаметофит является пыльцевым зерном, а женский гаметофит - зародышевым мешком. Внутри пыльника каждая диплоидная пыльцевая материнская клетка, делясь мейотически, образует четыре гаплоидные пыльцевые клетки, или микроспоры. Таким образом формируется мужское гаметофитное поколение. Стенки пыльцевых клеток становятся тоньше, а митотически делящееся ядро образует зрелое пыльцевое семя с двумя ядрами. Когда пыльник созревает и открывается, происходит высвобождение семян, которые переносятся на другие цветки ветром, насекомыми или другим способом.
Женский гаметофит развивается в завязи растения и полностью обособлен от окружающей среды. В завязи обычно имеется несколько центров роста, где развиваются семяпочки. В дальнейшем в этих структурах произойдет образование ткани женского гаметофита. В центре семяпочки материнская клетка - макроспора - делится мейотически и образует четыре гаплоидных макроспоры. Выживает только одна из четырех макроспор, которая делится митозом и образует зародышевый мешок с восемью ядрами (рис. 3-4, Б). Цитоплазма зародышевого мешка распределяется между шестью ядрами, образуя структуры, подобные клеткам. Одна из этих новых клеток становится яйцеклеткой, которая располагается у одного из полюсов зародышевого мешка, вблизи отверстия в семяпочке, называемого микропиле. Клетки семяпочки, окружающие зародышевый мешок, формируют защитные оболочки, или интегументы. В отличие от мужских пыльцевых зерен зародышевый мешок неподвижен.
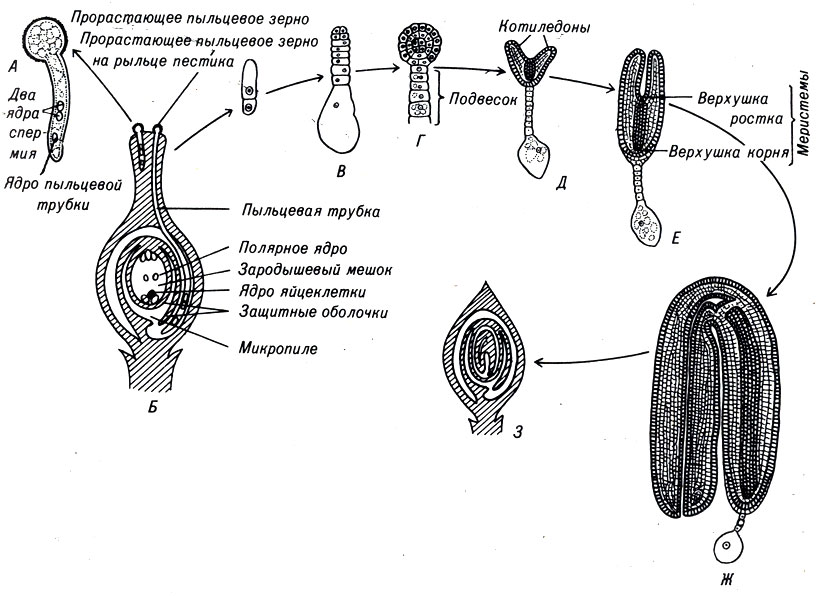
Рис. 3-4. Оплодотворение и раннее развитие цветковых растений. А. Проросшее пыльцевое зерно. Б. Пыльцевая трубка продвигается к завязи цветка, проходит микропиле и оплодотворяет семяпочку. В. - Ж. Стадии раннего развития зародыша растений. Показаны подвесок, котиледон и апикальная меристема. Все данные стадии развития проходят внутри семяпочки. Ж. Зрелый зародыш изогнут и повторяет форму семяпочки. 3. Зрелый зародыш внутри семяпочки
3.4. Опыление, перенос пыльцевых зерен от пыльника на рыльце пестика предшествует оплодотворению, в результате которого происходит объединение ядер яйцеклетки и сперматозоида
Вскоре после того, как пыльцевое зерно попадает на рыльце пестика, одно из двух ядер в пыльце делится и образует два ядра мужской половой клетки.
У пыльцевого зерна формируется труйса, которая растет внутрь пестика по направлению к семяпочке. Два ядра мужской половой клетки продвигаются по мере роста трубки. Этот процесс занимает от нескольких часов до нескольких дней, хотя у некоторых растений длится несколько месяцев (рис. 3-4, А).
Пыльцевая трубка входит в зародышевый мешок через микропиле семяпочки. При этом конец трубки разрывается и оба ядра мужской половой клетки попадают внутрь. Одно ядро сперматозоида сливается с поЯйрными ядрами семяпочки и формирует эндосперм, т.е. ткань, обеспечивающую питание зародыша. Эта ткань образуется в результате слияния трех ядер и поэтому содержит в три раза и более увеличенный гаплоидный набор хромосом. Другое ядро сперматозоида проникает в яйцеклетку и, объединяясь с ее ядром, образует диплоидную зиготу.
3.5. После оплодотворения начинается рост зародыша растения
Одно из отличительных свойств растений - их способность развиваться без помощи активно подвижных клеток. Увеличение массы клеток и морфогенез (становление формы растения) происходит исключительно за счет деления клеток. Становление же формы эмбрионального растения обусловлено неодинаковой скоростью деления клеток в отдельных участках зародыша и различиями в плоскости делений, которые по-разному ориентируют клетки относительно поверхности растения. (Рост зрелых растений также зависит от клеточных делений, которые в основном происходят в недифференцированных тканях, называемых меристемами. Поэтому они обычно считаются зонами, где происходит развитие эмбрионов.)
Сразу после оплодотворения ядра эндосперма начинают быстро делиться и образуют большое количество ядер внутри зародышевого мешка. Затем между ними возникают клеточные мембраны. Сформированные таким путем клетки эндосперма содержат большую часть цитоплазмы зародышевого мешка. Эндосперм является тканью, обеспечивающей питание эмбриона. Часто она представляет собой жидкость, например молоко кокосового ореха. После того как формирование эндосперма завершено, начинает делиться зигота. Образуются две структуры - собственно эмбрион (рис, 3-4, В-3) и подвесок (рис. 3-4, Д), который продвигает зародыш в эндосперм. Зародыш быстро развивается, и вскоре образуются клетки, которые в дальнейшем формируют главный стебель, или ось нового растения. На одном конце этой будущей оси находится зародышевый росток, или эпикотиль, на другом - зародышевый корень, первичный корешок. Первые эмбриональные листки, или семядоли (рис. 3-4, E), будут поглощать питательные вещества эндосперма и передавать их непосредственно в зародыш. Тем временем изменяются состав и строение наружных оболочек семяпочки. Она уплотняется и сохраняет зародыш до тех пор, пока он не прорастет и не образует молодое растение, или побег.
3.6. По мере развития эмбрионов растений их питательные потребности уменьшаются
Когда-то удавалось выращивать только относительно зрелые зародыши растений, поскольку культивирование ранних зародышей требует более сложной питательной среды. Позднее было обнаружено, что растворы, содержащие кокосовое молоко, способствуют развитию ранних зародышей, поскольку кокосовое молоко представляет собой жидкий эндосперм, в котором присутствуют растительные гормоны и органические вещества, необходимые зародышам. После того как эти гормоны были определены и очищены, появилась возможность выращивать очень ранние зародыши на полностью искусственных средах.
Эти эксперименты показывают, что способность к синтезу у ранних эмбрионов растений крайне ограничена и поэтому они зависят от наличия в окружающей среде как сложных, так и простых веществ. По мере развития зародышей увеличивается их способность синтезировать различные вещества, и в конечном счете они могут полностью удовлетворять свои потребности в сложных веществах, синтезируя их из простых.
3.7. Семена со зрелыми зародышами часто находятся в состоянии покоя
Зародыш, находящийся внутри семени, обычно способен выживать в таких условиях, в которых родительское растение погибает. Поэтому многие растения используют семена не только для размножения, но и для преодоления неблагоприятных условий окружающей среды. Некоторые семена могут находиться в состоянии покоя многие годы и сохранять способность к прорастанию. Так когда были высажены семена лотоса, имеющие возраст около 1000 лет, они проросли и развились в зрелые растения.
Состояние покоя предотвращает прорастание растений в неблагоприятное время. В зоне умеренного климата семена, созревшие в конце лета, падают на землю рядом с родительским растением и сохраняются в состоянии покоя. Они не прорастают до тех пор, пока не пройдет холодный период. Таким образом исключается прорастение семян осенью или во время короткой оттепели, которая, сменившись морозом, может погубить их.
Некоторые растения пустынь имеют очень короткий жизненный цикл и буйно растут только во время дождливого сезона. Семена этих растений не прорастают до тех пор, пока не наступит сезон дождей и не выпадет определенное количество осадков. После того как в почве накопится достаточное количество влаги, новое растение созревает и образует семена следующего поколения. Потребность семян во влаге регулируется растворимым веществом (ингибитором), от которого зависят сроки их прорастания. Только после того, как выпадет достаточное количество осадков, которые растворят и смоют этот ингибитор, может начаться прорастание.
3.8. После периода покоя семена прорастают, и у зародыша растения начинает развиваться корень
Прорастание семян включает в себя развитие корня, стебля (или ствола) и листьев. Однажды начавшись, развитие этих частей продолжается в течение всей жизни растения и может прерываться только на время сезонных периодов покоя. Каждый сезон у растения появляются новые стебли, листья и корни. У древесных пород растений увеличивается диаметр ствола. Вероятно, такой постоянный рост и развитие часто включают в себя те же процессы, которые происходят в зародыше.
Корни развиваются первыми. Для нормального развития молодого растения необходимо, чтобы корень пророс в почву, откуда он может извлекать воду и минеральные вещества. На кончике корня находятся молодые клетки, а ближе к основанию располагаются более старые (рис. 3-5). Корневой чехлик, расположенный на самом кончике корня, защищает более организованный и чувствительный участок корня от действия частиц почвы. Выше чехлика находится зона клеточного деления, корневая меристема, где формируются практически все новые клетки растущего корня. Хотя она является единственной зоной высокой митотической активности, наиболее заметный рост корня осуществляется не здесь. Сразу после деления клетки корневой меристемы приобретают кубическую форму, и, только когда последующие деления отодвинут их от кончика корня, начинается элонгация клеток, которая приводит к его удлинению. Элонгация клеток и поглощение воды обеспечивают продвижение корня в почве.
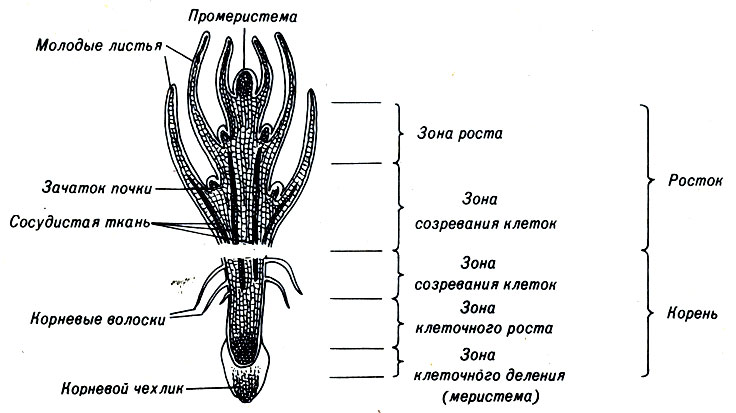
Рис. 3-5. Схема продольного разреза ростка и корня цветкового растения
Над зоной элонгации находится зона созревания, где клетки дифференцируются (рис. 3-5). Здесь проявляется дифференциальная активность генов, т. е. в процессе дифференцировки клетки используют только часть своей генетической информации. После того как клетки дифференцируются, они сохраняют свою специфичность в нормальных условиях в течение всей жизни растения.
Хотя корень взрослого растения состоит из дифференцированных клеток, меристема на кончике корня (апикальная меристема) сохраняет такой же эмбриональный характер, как и в прорастающем семени. Некоторые кольцевые слои клеток внутри зрелого корня также сохраняют меристематическую активность. Такой кольцевой слой называют камбием. Обычно существует пробковый камбий, образующий корневую кору, и сосудистый камбий, который постоянно увеличивает количество клеток в проводящей ткани. Таким образом корень может постоянно расти в длину и утолщаться.
3.9. Развитие проростка, как и развитие корня, происходит в апикальной меристеме
По мере роста верхней части развивающегося проростка формируются ткани стебля и листьев (рис. 3-5). Эту зону принято считать организующим центром растения. Основная организующая активная зона развивающегося проростка
находится в куполообразном выступе на самой верхушке и называется промеристемой. Типы клеток, составляющие промеристему, отличаются у разных групп растений.
Так же, как и у корней, в проростке различают ряд других зон роста, называемых камбиальными (рис. 3-6). В стволе деревьев и кустарников непосредственно под корой располагается пробковый камбий, из которого образуется кора. Глубже находится второй меристематический цилиндр, или сосудистый камбий. Обычно плоскость делеййй этих камбиальных клеток параллельна наружной поверхности ствола. Дочерние клетки, лежащие внутри кольца сосудистого камбия, образуют ткани ксилемы, по которым вода от корней перемещается вверх. Ежегодные приросты тканей ксилемы имеют кольцевую форму и увеличивают толщину ствола дерева. В зонах с умеренным климатом, где температура и количество осадков подвержены сезонным изменениям, годичные периоды роста отличаются друг от друга. На поперечном срезе ствола дерева ткани ксилемы располагаются в виде годичных колец. Дочерние клетки, лежащие снаружи сосудистого камбия, формируют ткань флоэмы, по которой перемещаются жидкости, в основном сок. Скорость клеточных делений в камбии зависит от температуры и влажности.
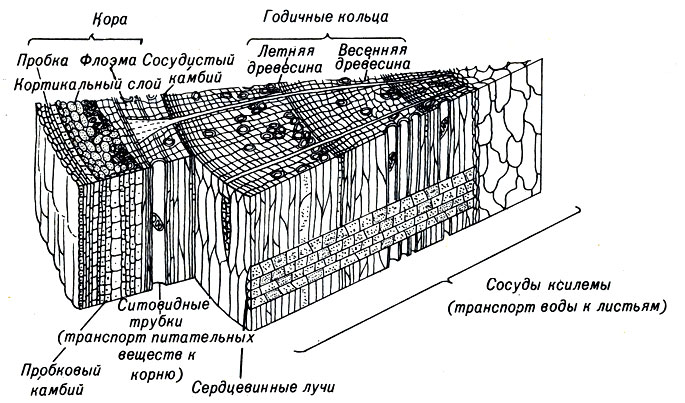
Рис. 3-6. Схема поперечного и радиального разрезов древесного ствола. Показаны слои меристемы, проводящие и опорные элементы, необходимые для роста растений
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'