Семейство кошек. Familia Felidae Gray, 1821
Высокоспециализированные хищники - наиболее специализированное семейство в подотряде и отряде.
Размеры мелкие, средние и крупные. Наиболее мелкие виды имеют длину тела около 50 см и вес 2-3 кг, более крупные (амурский тигр, Panthera tigris altaica) - длину тела до 300 см и вес до 390 кг (соотношение по длине тела 1 : 6, по весу 1 : 130-195). Тигр по размерам уступает только бурому и белому медведям. Все виды семейства, даже наиболее уклоняющиеся по внешнему виду (лев, гепард), имеют очень характерный, типично кошачий облик и в общем весьма однотипны. При довольно большом количестве видов (35) семейство несравненно более однородно; не только чем самые богатые видами семейства отряда - виверры (Viverridae - около 75 видов) и особенно куницы (Mustelidae - около 70 видов), но даже чем семейство волков (Canidae), содержащее несколько меньшее число видов (29).
Звери относительно легкого склада, очень гибкие, с удлиненным, у некоторых сильно вытянутым туловищем, обычно с длинным хвостом. Его длина не менее одной трети, иногда более половины длины тела, в виде исключения меньше трети. Ноги невысокие, сильные, и общие очертания тела вписываются в параллелограмм, иногда вытянутый. У немногих видов (рысь, каракал, особенно гепард) ноги высокие и общие очертания тела в профиль образуют квадрат или близкую фигуру. Грудная клетка небольшого объема, не выпуклая (малый объем легких и сердца), тело выглядит сжатым с боков и грудная часть по объему не больше брюшной. Передние ноги прямые, у большинства видов задние намного мощнее передних, хотя между их отделами стопа - голень - бедро - таз углы относительно невелики, крестцовая область приподнята и выше лопаточной и задняя часть тела выглядит особенно сильной - гораздо сильнее передней. Линия спины у этих видов (большинство) кзади повышается, иногда (значительно реже) почти прямая. Это особенно полное развитие структуры животного, способного к мощному прыжку с места (без разбега), свойственно как всем мелким видам, так и некоторым крупным (пума, ирбис). У немногих видов (большие кошки - лев, тигр, барс*, ягуар), передняя часть тела более мощная, спина повышается кпереди, область лопаток выше крестца или, реже, спина прямая (рис. 30).
* (Во всем тексте название барс употребляется, как это было искони, для вида Panthera pardus. В последнее время его часто именуют «леопардом» — названием, используемым в иностранных языках и русскому языку чуждым. «Барсом» в последнее время стали называть Uncia uncia. Для этого вида здесь применяется название снежный барс, или ирбис)
Шея умеренной длины или короткая. Голова относительно небольшая, с короткой лицевой частью, морда короткая и тупая. У немногих форм (некоторые кошки p. Panthera) лицевые части относительно увеличены и удлинены, но массивны и не бывают настолько вытянуты, как, например, у видов семейства волков. У многих лицевые части так укорочены, что форма головы приближается к шаровидной. Во всех случаях подбородочная часть выдвинута на уровень профиля оголенной части носа, что связано с положением резцов и сильным развитием и положением клыков.
Глаза большие, у мелких видов относительно очень большие и выпуклые, с округлым или щелевидным зрачком. Уши очень различны по величине и по форме - у некоторых (крупные виды) очень небольшие, иногда почти скрытые в мехе (лев, тигр и ирбис зимой), обычно же (мелкие виды) крупные, на вершине округленные или приостренные, поставленные к сагиттальной плоскости под небольшим углом, реже очень сильно наклоненные в стороны ("развешенные"). Иногда уши при значительной величине поставлены вертикально или почти вертикально, приострены и несут на вершине удлиненные волосы ("кисточку"; рысь, каракал). В некоторых случаях уши поставлены вертикально, округлены и очень велики (сервал). Общая площадь отверстий ушных раковин может быть больше площади проекции головы спереди. Ушная раковина с наружной стороны посредине вырезана, противокозелок хорошо развит, простой с валикообразным утолщением. Сильное развитие наружных частей органа слуха соответствует значительному развитию костных резонаторов и делает слух одним из главных органов чувств видов семейства. Губные ("усы") и надглазничные вибриссы развиты хорошо, особенно первые и особенно у мелких видов, нижнечелюстных нет.
Конечности пальцеходящие, на передней ноге 5 пальцев, причем I палец поставлен высоко вне общего ряда и очень мал, на задней - 4. Лапы передних и задних ног (особенно передних) сильно развиты, пальцы поставлены неплотно (лапа распущена - не в комке, как у волков); хорошо развиты пальцевые подушечки и большая подошвенная, имеющая треугольную форму. Карпальная мозоль одна, маленькая, метатарзальных нет. Лапа снизу опушена обычно не сильно и у большинства видов мозоли хорошо видны и отпечатываются на следу. Однако у некоторых видов, особенно в зимнее время (рысь) или весь год (барханная кошка, Felis margarita), волосы на нижней поверхности лапы столь густы и длинны, что совершенно скрывают подушечки. Все 4 пальца каждой конечности приблизительно равной мощности; III и IV длиннее, равной длины, II и V тоже равновелики, несколько короче средних. В целом у кошек лапа широкая и округлая. Все пальцы соединены эластичной кожной перепонкой, развитой в различной степени, но у большинства видов доходящей до пальцевых подушечек и до когтей (рис. 278). Пальцы весьма подвижны и могут широко раздвигаться в стороны. Это важная черта кошачьей лапы как хватательного органа.
Пальцы несут сильные когти. Они относительно не длинные, но круто изогнутые, резко сжатые с боков и очень острые. Когти у всех наших кошек, кроме гепарда, втяжные, т. е. в спокойном состоянии и при ходьбе они резко оттянуты назад и подняты. Острые концы их, хотя и направлены в общем вниз, но земли не касаются. Кошка идет на подушечках, находящихся в месте соединения I и II фаланг. Особые сочленовные поверхности фаланг позволяют очень широкое вертикальное движение их друг относительно друга, а система сильных связок осуществляет это движение-втягивание и выпускание когтей (рис. 17). Особую роль, кроме системы мелких связок, соединяющих I и II фаланги, играют две больших связки, идущих по верху и по низу всех фаланг и крепящихся спереди к I. В спокойном состоянии III фаланга поставлена под углом около 45°, а дистальный конец ее направлен вверх, II стоит по отношению к ней почти под прямым углом, направлена вниз. I фаланга плотно прижата ко II. Главная ось ее параллельна таковой II фаланги и направлена вверх и несколько назад. При таком положении коготь втянут. При выпускании когтя (волевой акт) работой связок, главным образом нижней, III и II фаланги ставятся в одну линию, I же становится по отношению ко II так, что ее вертикальный диаметр принимает вертикальное положение и несколько наклоняется вперед. Коготь не только выдвигается, но и конец его направляется вниз. Могут изгибаться вниз и пальцы целиком. Раздвигание пальцев, выпускание когтей и изгиб пальцев делает лапу кошек мощным и совершенным хватательным инструментом.
Хватательная способность лапы кошек в большей степени определяется формой когтей. Изгиб их бывает различен, но всегда крут, у некоторых же наружная поверхность когтя имеет правильные очертания дуги окружности малого радиуса. У ряда видов, особенно крупных кошек, коготь укреплен на фаланге тем, что его основание окружено особым, хорошо развитым костным "воротничком". В целом хватательный механизм совершеннее развит на передних конечностях, которые собственно и играют роль главного хватательного инструмента (рис. 17).
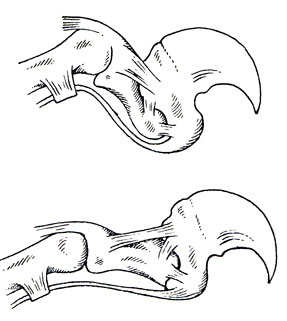
Рис. 17. Скелет и связки пальца тигра, определяющие положение и механизмы выпускания когтя. Верхний рисунок - положение покоя, коготь втянут, нижний - коготь выпущен. По В. Мазаку, 1965 (рис. Н. Н. Кондакова)
Во вполне типично развитой кошачьей лапе у каждого когтя имеется хорошо развитая, сильно растяжимая кожная складка, чехлом одевающая втянутый коготь с обеих сторон и сверху. Эти складки дополнительно защищают коготь и его конец. Они сильно растягиваются при выпускании когтя*. У видов с сильным развитием этих чехлов, как правило, особенно сильно развиты и межпальцевые перепонки и доходят до пальцевых подушечек и когтей. У некоторых видов эти складки (лопасти), одевающие когти - все или часть из них - уменьшены в размерах или развиты слабо. В разной степени сокращены бывают и межпальцевые перепонки. Сильнее всего уменьшены и те, и другие у гепарда (рис. 278). Лапа его имеет и некоторые другие особенности, в частности когти втянуты в меньшей степени и видны снаружи (см. характеристику рода гепардов).
* (Иногда встречающиеся в литературе указания на "роговые чехлики у основания когтей" (Сапоженков, 1957 и др.) неправильны. Это, по-видимому, ошибочные представления об описанных кожных складках.)
Когти играют роль только при хватании, удерживании и убивании добычи и при самообороне, в меньшей степени при лазании на деревья и никак не участвуют в ходьбе и рытье. Это обстоятельство и описанные механизмы сохраняют их концы всегда острыми и готовыми к выполнению основных функций. Вместе с тем, в таком строении лап, вероятно, одна из причин неспособности кошек к быстрому и длительному бегу. Гепард - единственная хорошо и быстро бегающая кошка, охотящаяся догоном. Это даже самый быстрый бегун среди млекопитающих.
Лапа с втяжными когтями - характерная черта семейства, и среди других хищников она имеется только в небольшой группе виверр (Prionodontinat; рода Prionodon, Pardictis).
Волосяной покров у большинства густой, мягкий и шелковистый. Его длина, густота и степень развития подшерстка значительно отличаются в зависимости от климатических условий мест обитания. В суровом климате хорошо выражены сезонные различия в характере меха. У большинства видов волосяной покров на всем теле более или менее одинаковой длины. Лишь у немногих видов (рыси) на щеках удлиненные волосы образуют "баки", у самцов львов на передней части туловища, на шее и на голове до ушей имеется густая и длинная грива и подвесы из удлиненных волос по груди и брюху, и у гепарда волосы по верху шеи и на холке удлинены.
Окраска весьма разнообразная, преимущественно пятнистая из темных пятен по светлому основному полю. Форма пятен очень разнообразна - простые сплошные мелкие пятна, более крупные кольцевые, составленные из отдельных пятен, сплошные округлые или вытянутые кольца и т. п. Нередко пятна образуют довольно правильные продольные или поперечные ряды, или часть из них сливается в поперечные или продольные полосы, и окраска представляет собой комбинацию из полос, вытянутых и округлых пятен. Встречается окраска без пятен на туловище, только из одних резко выраженных поперечных полос (тигр).
Основной тон окраски обычно светлый - от почти белого, светло-желтого и рыжеватого до ярко-рыжего и коричневого. Пятна обычно черные разной интенсивности и резкости очертаний, иногда рыжеватые. Немногие виды окрашены одноцветно (каракал, пума, лев) или почти одноцветно (барханный кот, Felis margarita), причем окраска обычно песчано-желтая или коричневато-желтая, или подобная. Иногда при одноцветной общей окраске имеется лишь слабо развитая пятнистость или только следы ее на конечностях (некоторые формы рыси) или узор на голове (одна из цветовых форм Felis temminckii). Обычно даже одноцветно окрашенные виды котятами или в молодом возрасте имеют более или менее развитую пятнистость. Пятнистый тип окраски у современных кошек следует считать исходным.
Индивидуальная изменчивость окраски выражена хорошо, у многих видов значительна, географическая также. Последняя проявляется в большинстве случаев в изменении общего тона окраски, частью характера пятнистости. У некоторых видов встречаются меланисты, альбиносы и хромисты, у которых намечается географическая локализация цветовых уклонений [черные "пантеры" (барсы) Индии, хромисты манулы Туркмении, и др.]. Немногие виды имеют полифазную окраску (ягуарунды, Felis yaguarundi; Америка). В одном случае (Felis temminckii) окраска, не будучи в прямом смысле полифазной, изменяется очень резко (пятнистая - одноцветная, темная - светлая), причем намечается географическая локализация цветовых типов. У льва половой диморфизм выражен в развитии гривы у самца. У всех кошек самцы крупнее самок.
Анальные железы обычно имеются, но развиты слабо. Penis лежит непосредственно у мошонки. Glans рудиментарен, несет роговые шипики. Сосков 2-4 пары.
Череп характеризован рядом семейственных признаков весьма четко. Однако в связи с очень большими различиями в абсолютных общих размерах разных видов и другими причинами общий облик и пропорции черепа могут меняться сильно. По общим очертаниям череп широкий, с широко расставленными скуловыми дугами. Самая передняя часть его - от переднего края глазниц ("морда") - короткая и широкая. Этмотурбинальные раковины велики, простираются вперед почти до отверстия носовой камеры и лежат поверх максиллотурбиналий. Алисфеноидного канала нет.
Костный слуховой барабан обычно высокий, округлый (вздутый), построен из ectotympanicum и entotympanicum и внутри соответственно разделен перегородкой на две камеры. Энтотимпальная (задне-внутренняя) часть всегда, хотя и в разной степени, больше эктотимпанальной, которая несет слуховое отверстие. Наружное разделение двух частей барабана обычно не заметно и лишь у некоторых форм оно резко. Слуховой трубки нет или она лишь намечена. Относительная величина слуховых барабанов очень различна - у мелких видов они велики и вздуты (у пустынных особенно сильно) и тонкостенны, у крупных относительно малы, часто плоски и толстостенны. Мастоидный и парокципитальный отростки разделены, причем последний уплощен, обычно плотно прилегает к задней стенке барабана, и вершина его у большинства не выдается за нижнюю точку барабана.
Главные общие изменения облика черепа имеют следующие направления. Череп более или менее нейтрального типа (рысь, каракал; Верещагин, 1967); общие очертания представляют собой слабовытянутый овал, поперечник которого составляет около 75% кондилобазальной длины. Межглазничное и заглазничное сужения широкие, мозговая коробка очень объемистая, слабо вытянутая, сильно вздутая, скуловые дуги имеют наибольшую ширину на уровне сочленовных поверхностей, однако, их общие очертания выпукло- дугообразные. Всякого рода гребни и бугры, служащие для крепления мышц, прежде всего жевательных, развиты слабо, сагиттальный выражен, но мал, затылочный развит хорошо.
Ряд, образуемый видами более крупных и самых крупных кошек и завершаемый тигром, характеризуется тем, что череп становится относительно гораздо более грубым, тяжелым и массивным, лицевая часть, в частности, морда, относительно увеличивается и удлиняется, мозговая часть становится относительно короче. Весь череп приобретает более вытянутую форму, хотя скуловые дуги в своей задней части могут быть разведены очень широко. Впереди их очертания не столь выпуклы и они образуют более угловатую фигуру. Межглазничное и особенно заглазничное сужения становятся относительно меньше, объем мозговой коробки уменьшается и она не столь вздута. Сильнее развиваются всякого рода бугры и гребни; особенно велики сагиттальный и затылочный, достигающие большой высоты - мозговая коробка сильно сжата мощной жевательной мускулатурой. Зубы, в частности клыки, мощные, с широким основанием, но относительно короткие. Слуховые барабаны относительно небольшие, в крайнем случае уплощенные, толстостенные. Нижняя челюсть тяжелая, относительно длинная, с выступающей вперед сильной передней частью, несущей большие клыки с длинными и массивными основаниями. Это череп животных, убивающих и разрывающих крупную добычу, которая часто превышает их собственный размер и вес - у самых крупных даже в несколько раз.
Череп мелких видов характеризуется, наоборот, легкостью общего строения, тонкостенностью мозговой коробки и, в сущности, отсутствием всякого рода гребней и бугров, кроме затылочного. Сагиттальный гребень не развивается совершенно или имеется маленький участок у затылочного, и лировидная фигура обычно широка. Лицевые части сокращаются еще больше, морда становится совсем короткой и тупой, глазницы сильно увеличиваются и все больше направляются вперед; скуловые дуги сильнее разводятся в средней части (не только сзади) и образуют более или менее правильную дугообразную фигуру. Главное же в том, что мозговая коробка, иногда умеренно вздутая и несколько вытянутая, становится все более объемистой, вздутой и короткой, и весь череп становится коротким и выпуклым с широким межглазничным и заглазничным пространством (см. рис. 19).
Общий абрис черепа, у некоторых форм образующий приблизительно такой же овал, как у рыси или даже несколько более вытянутый (удлиненная мозговая коробка), имеет тенденцию становиться более широким и коротким, и у крайней формы (манул, F. manul) приближается уже к окружности. В целом череп мелких кошек сравнительно с крупными выглядит инфантильным. Зубы относительно слабее, клыки относительно длиннее, но тоньше. Нижняя челюсть короткая, легкая, с относительно слабой - не массивной -передней частью. Это линия специализации мелких хищников, убивающих небольшую добычу, значительно меньшую собственных размеров (грызуны). Если кошек считают лучшим выражением хищника, то указанные две формы представляют собой высшее выражение типа кошки, хотя морфологически они противоположны.
Зубная система представляет собой одну из крайних степеней специализации в пределах отряда как по числу зубов, так и по некоторым особенностям их строения. Полная зубная формула 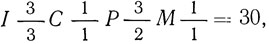 , однако у некоторых видов второй верхний предкоренной (первый в ряду - первого верхнего предкоренного у кошек вообще нет) иногда не развивается (теряется) или всегда отсутствует (манул, рысь) и она принимает вид
, однако у некоторых видов второй верхний предкоренной (первый в ряду - первого верхнего предкоренного у кошек вообще нет) иногда не развивается (теряется) или всегда отсутствует (манул, рысь) и она принимает вид 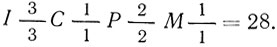
Диастема у большинства велика, особенно в нижней челюсти. Это связано с отсутствием первого предкоренного в верхней челюсти и первого и второго - в нижней, что в свою очередь зависит от сильного развития клыков при коротких челюстях.
Зубы мощные, резко выраженного секториального (режущего) типа. Хищнические зубы большие, сильно сжаты, относительно длинны, резцы небольшие и слабые, образуют прямые поперечные ряды, клыки большие. У крупных видов они более массивны и коротки, у мелких относительно тоньше и длиннее, иногда очень длинны и тонки (дымчатый леопард), у большинства с неглубокими продольными бороздками. Второй предкоренной (верхний) очень мал, конической формы, с одним корнем, третий верхний предкоренной хорошо развит, сжат с боков, обычно с высокой и острой средней вершиной и двумя маленькими, расположенными на ее основании впереди и сзади нее. Верхний хищнический зуб (четвертый предкоренной) очень большой, сжатый с боков, с тремя вершинами, лежащими на одной оси - первая вершина невысокая, коническая, вторая высокая, острая, третья - невысокая, удлиненная с режущим краем; в передне-внутренней части зуба у большинства имеется дополнительная лопасть с небольшой, обычно хорошо развитой тупой вершиной. Первый верхний коренной рудиментарен - он очень мал и поставлен поперек ряда (иногда теряется при жизни).
В нижней челюсти оба предкоренных (третий и четвертый) трехвершинные, подобные третьему предкоренному верхней. Хищнический зуб (первый коренной) сильный с двумя хорошо развитыми режущими вершинами. Молочный хищнический у некоторых видов сравнительно с постоянным усложнен.
Характерная особенность скелета - очень большая гибкость позвоночника и увеличенные размеры и особая форма сочленовных поверхностей фаланг и положение их друг относительно друга, допускающих втягивание и выпускание когтей (см. выше). Грудных позвонков 13. Os penis рудиментарна.
Относительная длина кишечника в связи с исключительно мясным питанием мала, относительный вес сердца мал и гораздо меньше, чем у видов семейства волков.
Обитатели самых разнообразных ландшафтов - от влажных тропических лесов до песчаных пустынь. Обычно в той или иной степени связаны с древесной или кустарниковой растительностью (укрытия). Живут в теплом и умеренно-теплом климате без постоянного или со слабым снеговым покровом. Лишь один вид (рысь) постоянно связан с областями со снежной зимой на равнине или в горах, один - со снежным высокогорьем (ирбис) и у некоторых часть ареала лежит в областях со снежной зимой (пума, амурский кот, лесной кот, тигр и некоторые другие).
Все виды - ярко выраженные хищники, добывающие живую добычу, главным образом теплокровных животных самой различной величины - от мелких грызунов и птиц до жираф и буйволов. Реже ловят рептилий, немногие в некоторой мере специализируются на добывании рыбы. Ловят, подстерегая на тропах, водопоях и солонцах, или скрадывая и настигая одним или немногими прыжками. К длительному бегу и преследованию неспособны. Лишь гепард может преследовать добычу на протяжении сотен метров и настигать даже газелей, развивая наибольшую для млекопитающих скорость*. Одиночные животные, не составляющие пар даже на время выкармливания детенышей. По-видимому, лишь один вид (лев) держится парами и семейными группами и охотится группами. Логово как открытое, так и в норах, расщелинах скал и пещерах и т. п. Главным образом ночные животные.
* (Обычно считается, что гепард может бежать со скоростью до 140 км в час. По новым сведениям, максимальная установленная скорость равна 56 миль (90,12 км) в час (Пурнелл, 1964).)
Ареал семейства очень обширен и занимает все зоогеографические области, кроме Австралийской (рис. 18). В Новом Свете кошки распространены от северного побережья материка до Патагонии и Магелланова пролива. На американских островах Берингова моря, кроме Нунивака (?), видов семейства нет, в Арктическом архипелаге граница ареала охватывает Баффинову землю и остров Саутгэмптон, а также мелкие острова Гудзонова залива, Нью-Фаундленд, острова залива Святого Лаврентия, острова западного побережья Северной Америки (Кадьяк, Ванкувер, архипелаг Александра и др.). На островах Вест-Индии, включая Багамские, Кубу и Тринидад, представителей семейства нет. У побережий Южной Америки в ареал входит Чилое; на Огненной Земле и Фолклэндских островах кошек нет.
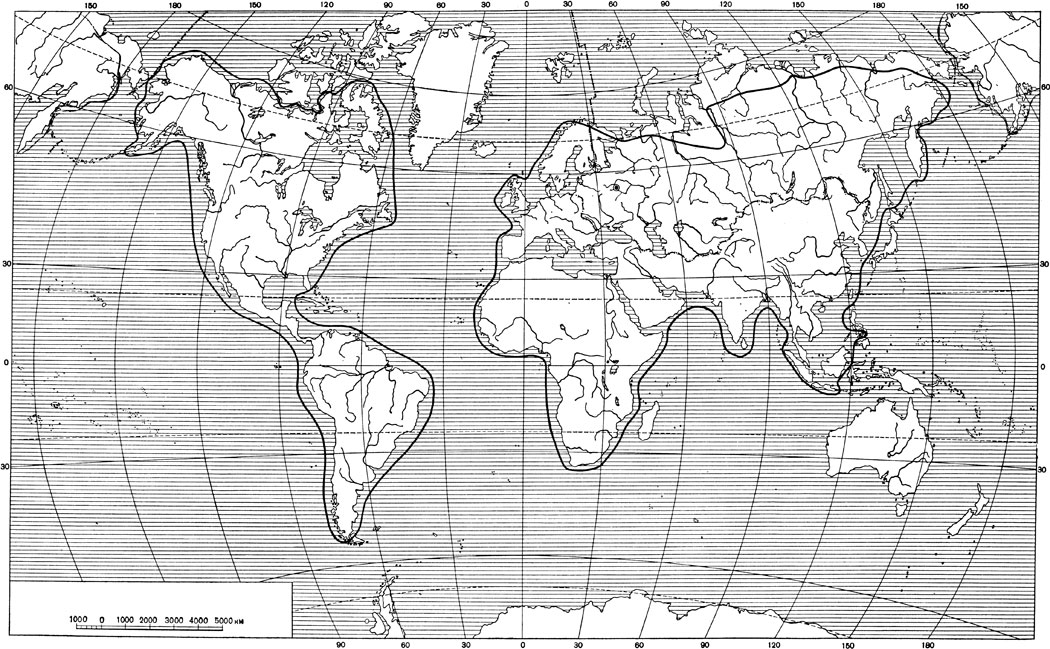
Рис. 18. Востановленный ареал семейства кошек, Felidae и рода мелких кошек, Felis L. В. Г. Гептнер
В Старом Свете ареал охватывает почти всю Европу на север приблизительно до границы леса. На западе ареал включает Англию, в Ирландии видов семейства нет и, очевидно, по крайней мере в историческое время, не было. В Средиземном море ареал занимает Корсику, Сардинию, Сицилию, Крит и Балеарские острова (на остальных островах, в том числе Родосе, Кипре и Мальте, кошек нет). На мелких островах и архипелагах Атлантического океана и на Исландии видов семейства нет и не было, как и на островах Баренцева моря.
В Азии северная граница ареала идет по северной границе леса или несколько южнее, на северо-востоке Сибири захватывая Камчатку. Южная граница охватывает Аравийский полуостров и Индию с Цейлоном, проходит по побережью Бенгальского залива, спускаясь на юг, охватывает Суматру и Яву и идет между Бали и Ломбоком, захватывает Калимантан (Борнео) и Палаван и некоторые из Филиппинских (по-видимому, только средние Негрос и Цеву); на архипелаге Ментавай (западнее Суматры) видов семейства нет, как и на о-вах Билитон, Банка и Мадура (Яванское море). Нет кошек и на мелких островах к югу от Сингапура, между Суматрой и Калимантаном (Борнео) и в Южно-Китайском море. Далее граница ареала охватывает Гайнань и Тайвань и, оставляя за пределами ареала о-ва Рю-Кю и Японские, но захватывая Цусиму и Чжечжудо (Квельпарт), охватывает Сахалин и, оставляя вне ареала Курильские острова, идет к Камчатке. На Карагинском и Командорских островах видов семейства нет; нет, по-видимому, и на Шантарских островах.
Ареал занимает весь материк Африки. Мадагаскар и острова Атлантического и Индийского океанов в ареал не входят. Кошки, особенно крупные, в некоторых местах истреблены, и ареал семейства сократился. С другой стороны, домашние кошки, которые в известных условиях, особенно в теплых странах, легко дичают, заселили некоторые острова, лежащие вне естественного ареала семейства. Так, описанная в качестве особого вида с Тимора (и Ротти) Felis megalotis, несмотря на некоторые настойчивые возражения (Вебер, 1928), признана одичавшей домашней кошкой*.
* (Эта форма вообще не значится ни в новейших каталогах семейства, ни в фаунистических сводках (Лори и Хилл, 1954, например). Не обсуждается и зоогеографический казус, связанный с возможным обитанием вида семейства в Австралийской зоогеографической области.)
Семейство кошек, после того как из него исключили мадагаскарскую Fossa (Viverridae), представляется весьма цельной естественной группой. Она хорошо отграничена от соседних семейств, прежде всего от ближайшего - виверр (Viverridae)*. Это относится к семейству во всем его объеме, т. е. включая ископаемые виды. Даже такие крайние формы, как саблезубые кошки, типичны для семейства. Попытка возвести в семейственный ранг некоторые ископаемые группы, главным образом в пределах саблезубых кошек (крайняя точка зрения - Кретцой, 1929), не нашли признания (Симпсон, 1945; Громова, 1962). Впрочем, отдельные современные авторы саблезубых кошек в особое семейство (Machairodontidae) все же выделяют (Тениус и Гофер, 1960). Совершенно не обоснованы и не обратили на себя никакого внимания предложения выделить из современных Felidae в отдельные семейства гепардов и рысей (Guepardidae Gray, 1869; Lyncidae Schulze, 1900).
* (Связь с ним, впрочем, достаточно ясна. Так, виверры родов Prionodon и Pardictis (линсанги Юго-Восточной Азии) имеют характерный для кошек короткий penis, прилежащий к scrotum, у них нет перинеальных желез и когти втяжные с характерными для кошек кожными когтевыми складками — чехлами.
Изолированное положение семейства подчеркивается и тем, что по серологическим признакам кошки отличаются от других хищных очень резко. Ранг этих отличий больше, чем отличий между некоторыми хищными и всеми ластоногими (В. И. Борисов).)
В настоящее время наиболее естественно принимать семейство в более широком объеме и рассматривать его как состоящее из пяти подсемейств - вымерших Proailurinae Zittel, 1893; Nimravinae Trouessart, 1885; Machairodontinae Gill, 1872 и Hyaenaelurinae Pilgrim, 1932 и современного Felinae Trouessart, 1885. При этом Machairodontinae (нижний олигоцен-плейстоцен; около 10 родов) рассматриваются как наиболее специализированная группа семейства, стоящая в ряду выше Felinae (Симпсон, 1945). Наиболее примитивные кошки, относящиеся к подсемейству Proailurinae (около 5 родов), существовали с позднего эоцена (с раннего олигоцена) по средний плиоцен в Старом Свете. В некоторых своих ранних формах они соединяют отдельные признаки кошек и виверр ("структурные предки"), указывая как на путь развития семейства, так и на его ближайшие связи.
Исходные формы ("предки") современных кошек находятся среди подсемейства Nimravinae (верхний эоцен - нижний плиоцен Евразии, Северной Америки и Африки; около 9 родов). Ископаемые формы современного подсемейства Felinae известны из Азии и в плейстоцене (может быть, плиоцене) Северной и Южной Америки. Кроме форм p. Felis описаны два ископаемых азиатских нижнеплиоценовых рода (Sivaelurus Pilgr, Dinofelis Zdansky) и один плейстоценовый (Sivapanthera Kretzoi).
Систематика современных кошек в некоторых частях не проста по существу, но, главное, она очень запутана и подчас все больше усложняется и в наши дни. Это относится к надвидовым группировкам (вопрос о видах кошек теперь уже ясен). Сейчас едва ли есть семейство современных млекопитающих, во взглядах на систему которого были бы столь разительные противоречия. Так, долго державшееся разделение современных кошек на 2 подсемейства-гепардов, Acinonychinae и остальных, Felinae, заменилось представлением о 3 подсемействах - из собственно Felinae были выделены Pantherinae - "большие" кошки (Покок, 1917; Огнев, 1935 и многие др.). Позже было предложено делить современных кошек на 4 подсемейства, выделяя из собственно кошек, Felinae - рысей, Lyncinae (Хальтенорт, 1957; Вейгель, 1961; Мазак, 1965 и др.). Крайняя точка зрения, не встретившая, однако, никакого интереса, выражается в разделении всех кошек на 15 подсемейств, причем среди современных в особое - пятое подсемейство, выделяются еще и дымчатые леопарды Neofelinae, с 1 родом и видом (Кретцой, 1929).
Взгляды разных современных авторов на разделение кошек на уровне подсемейства весьма различны. Большинство принимает 2 (гепарды, Acinonychinae и остальные кошки, Felinae) или 3 (гепарды, большие кошки, Pantherinae и остальные) подсемейства. Немногие признают 4 подсемейства- принимая еще отдельное подсемейство рысей (Lyncinae). При этом положение отдельных видов оказывается весьма различным: дымчатый леопард (F. nebulosa) оказывается то в Pantherinae, то вне этой группы, манул относится то к Felinae и стоит в соседстве с барханным котом (F. margarita), то оказывается в "подсемействе" рысей (Lyncinae).
Последнее - особенно наглядный пример формального подхода к системе, нарушающего ее естественность. Манул по всем своим краниологическим и другим признакам - непосредственно ближайшая форма к барханному коту (см. далее в систематической части) и представляет собой крайнюю степень специализации "мелких кошек" и современных кошек вообще. Однако у него нет переднего верхнего подкоренного (Р2) и по этому единственному формальному признаку он попадает в ближайшее родство и в одну группу с рысями, с которыми он, по существу, имеет мало общего.
Выделение подсемейств в других случаях обосновано еще меньше. Так, сравнение диагнозов подсемейства мелких кошек, Felinae, больших кошек, Pantherinae, и рысей, Lyncinae (см. например, Мазак, 1965) показывает, что в конечном счете единственным признаком Lyncinae оказываются кисточки на ушах. Совершенно очевидно, что этого признака недостаточно.
Выделение в отдельное подсемейство Pantherinae больших кошек - льва, тигра, барса, ягуара, обычно также и ирбиса и дымчатого леопарда (Neofelis)* на первый взгляд, естественное, тоже неприемлемо. Резкого отграничения от Felinae здесь нет. Что касается дымчатого леопарда, то он вообще не имеет признаков, характерных для Pantherinae, собственно, т. е. льва, тигра, барса ягуара, а у ирбиса признаки "Pantherinae" и "Felinae" комбинируются почти в равной пропорции (Хеммер, 1967; см. также характеристику рода ниже). Таким образом, ясной границы даже между наиболее типичными "большими" кошками ("Pantherinae" - лев, барс, ягуар, тигр) и "мелкими" ("Felinae") на подсемейственном уровне нет. В виде ирбиса есть даже настоящее "связующее звено", причем признаков "промежуточного положения" у этого вида больше, чем принято считать (см. описание его). Иногда разделение современных кошек на подсемейства вообще не проводится и все они относятся к одному, Felinae, противопоставляемому упомянутым вымершим (Эллермэн и Моррисон-Скотт, 1951, 1966; Эллермэн, Моррисон-Скотт, Хэймен, 1953). Эта точка зрения наиболее правильно отражает естественные отношения видов семейства, хотя и не может считаться самой распространенной. Она принимается и здесь, поскольку совершенно четкое разграничение даже родов современных кошек в сущности почти невозможно, (см. характеристики родов ниже) или затруднительно. Это относится даже к гепарду, особенности которого всегда обращают на себя внимание. Действительные отличия этого рода меньше традиционных представлений о них (см. ниже характеристику рода). Таким образом, в современных кошках можно видеть лишь одно подсемейство**.
* (Пума, несмотря на свои крупные размеры, как давно признается даже сторонниками крайних взглядов, к группе больших кошек (p. Panthera, «Pantherinae») отношения не имеет. Это, хотя и большого размера, но по всем морфологическим признакам настоящая «мелкая» кошка (Felis). В дальнейшем она здесь среди «больших» кошек (в любом толковании группы) не приводится и не подразумевается.)
** (Число работ, посвященных надвидовой систематике кошек, очень велико. Одна из первых и наиболее интересных, оказавшая большое влияние на всех позднейших авторов и имеющая значение и сейчас, принадлежит Н. А. Северцову (1858). Важны работы Покока (1917) и Хальтенорта (1936—1937). В наши дни вышел ряд работ, главным образом морфологических, но в последние годы и этологических (Вебер, 1928; Кретцой, 1929; Царапкин, 1932; Огнев, 1935; Покок, 1939; 1951: Симпсон, 1945; Лейхаузен, 1950, 1956; Хальтенорт, 1953; Тениус и Гофер, 1960; Вейгель, 1961; Хеммер, 1964; Мазак, 1965; Верещагин, 1967 и др.). Значительная часть материалов по крупным кошкам сведена и ревизована Хеммером (1966).)
Гораздо более запутана картина родовых группировок современных кошек. Это объясняется не столько сложностью проблемы самой по себе, но главным образом неясностью представлений о роде в современной таксономии - прежде всего своеобразным страхом некоторых систематиков перед "большими" родами и стремлением создавать роды с более или менее одинаковым числом видов в каждом*. В связи с этим широко развита тенденция далеко идущего дробления родов. Кошки оказались в этом отношении излюбленным материалом и, пожалуй, нет другой группы млекопитающих, в которой эта тенденция получила бы столь широкое развитие и привела бы к таким большим разноречиям и искажениям. Есть, впрочем, и среди современных систематиков авторы, рисующие, можно сказать, достаточно естественную картину надвидовых группировок кошек. Все они принимают очень малое число родов.
* (О «больших» родах в структуре систематических групп и их эволюционном значении см. Гептнер, 1965.)
В сущности, первый широкий и теоретически обоснованный анализ надвидовых объединений в семействе принадлежит Н. А. Северцову (1858). Он выделил всего 5 родов с 27 подродами: Tigris (лев и тигр), Panthera (ягуар,барс, ирбис, пума - 4 подрода), Cynailurus (гепард), Lynchus (рыси и каракал - 2 подрода) и Felis (все остальные кошки - 17 подродов). Система Северцова, в общем очень сдержанная и разумная, была превосходна и не только для своего времени. Ее взял за основу Покок (1917), от которого идет одна из современных распространенных точек зрения. Однако, она была им сильно изменена в ее существе в том смысле, что большинство северцовских подродов, прежде всего из рода Felis были возведены в родовой ранг. Тем самым естественный и с современной точки зрения "большой" северцовский род Felis был уничтожен. Всего Покок (1917) разбил семейство на 17 родов с подродами.
Некоторые позднейшие териологи пошли в направлении дробления родов еще дальше или так или иначе комбинировали тот же, или приблизительно тот же обширный набор родов. Так, некоторые принимают (при трех или четырех подсемействах) 14 (Хальтенорт, 1957), 19 (Вейгель, 1961), 20 (Мазак, 1965), и даже 24 рода (Огнев, 1935). Эта система сомнительна уже с теоретической стороны. При 20 родах 13, т. е. 65%, оказываются монотипическими, 1 содержит 2 вида, 3 - по 3 вида, 1-4 вида и 1-5 видов (Мазак, 1965). При 24 родах (Огнев, 1935) число монотипических родов становится еще больше. Таким образом, родовые характеристики, признаки и само понятие, в сущности, подменяются видовыми и совпадают с ними. Естественное понятие монотипического рода в данном случае в значительной степени теряет свой смысл.
Что же касается фактической стороны, то оказывается, что между некоторыми видами одного рода отличия оказываются несравненно больше и существеннее (Мазак, 1965), чем различия между родами, которые трудно уловимы (Felis - Prionailurus; Lynx - Caracal и т. п.) и фиктивны. У разных авторов далее, не только резко по-разному оценивается ранг отдельных форм (большие кошки - лев, тигр, барс, ягуар - 1 род или 4 монотипических рода и т. п.), но они занимают и очень разные места в ряду (эволюционном, специализации) форм семейства. Так, манул в виде особого рода находится то в непосредственном соседстве с родом Felis (Мазак, 1965), то между ними ставится 14 родов с 7 подродами и он относится к другому "подсемейству" - рысям (Вейгель, 1961) и т. п. Все это ясно указывает на искусственность далеко идущего дробления современной части семейства на рода.
Противоположная крайняя точка зрения сводится к признанию лишь двух родов - гепарда, Acinonyx, с одной стороны, и рода Felis, в который включаются все остальные современные кошки - с другой. В последнем обычно более или менее широко используется разделение на подроды. Эта система, во многом вполне разумная (см. далее), в настоящее время имеет ряд сторонников (Аллен, 1938; Новиков, 1956; Холл и Келсон, 1959 и др.).
Отмеченное сложное положение указывает на то, что, при всем разнообразии величины и облика отдельных видов, при больших краниологических различиях и при относительном большом числе видов (35), современные кошки представляют собой в морфологическом, систематическом и филогенетическом отношении весьма однородную группу. Практически имеется положение, при котором почти любая пара видов, даже очень несходных между собой, оказывается связанной через другие, образующие более или менее непрерывную морфологическую цепь. Разрывы этой цепи или мозаики форм на многочисленные самостоятельные роды почти неизбежно оказываются искусственными. Даже признаки гепарда (см. далее) менее резки и своеобразны, чем обычно представляется. Это относится и к признакам поведения (Лейхаузен, 1950, 1956; Хеммер, 1964, 1966 и др.).
Интересно, что у всех видов, насколько известно, одинаковое число хромосом 2n = 38, различия в кариотипе незначительны и лишь два (F. pardalis, F. wiedi) имеют 36 хромосом (В. Н. Орлов). В искусственных условиях (зоопарки) получены гибриды между всеми формами крупных кошек (лев, тигр, барс, ягуар), даже тригибриды; были удачны и обратные скрещивания. Пока нет лишь гибридов с ирбисом, который, однако, вообще почти не размножается в неволе и тигра с барсом (сведения о таком гибриде в природе мало определенны). Зато получены гибриды барса с пумой, не относящейся к большим кошкам (p. Panthera), т. е. межродовой гибрид (Хеммер, 1966).
Несмотря на указанные особенности семейства и разноречия в их толковании, известные естественные группы, которым соотносительно с другими возможными группировками видов можно придавать родовой ранг, среди современных кошек все же выделяются. Таких групп принимается 3 или 4. Согласно одной точке зрения (Симпсон, 1945), в семействе 3 рода - большие кошки, Panthera (лев, барс, тигр, ягуар, ирбис, дымчатый леопард), гепард, Acinonyx, и мелкие кошки, Felis, включающий все остальные виды семейства. Таким образом, предложенные Пококом (1917) подсемейства рассматриваются лишь как роды. Согласно близкой точке зрения (Эллермэн и Моррисон-Скотт, 1951, 1966) признаются те же роды и в том же составе, однако дымчатый леопард (F. nebulosa) выделяется в отдельный род (Neofelis), занимающий положение между pp. Panthera и Felis; всего принимается 4 рода (Палеарктика и Индия).
Как выясняется все больше (см. указанные выше новые работы, особенно Хеммер, 1964, 1966), Neofelis не имеет отношения к большим кошкам (Panthera). Не занимает он и промежуточного положения между ними и мелкими кошками (Felis). Это настоящая мелкая кошка, очевидно, не более своеобразная, чем некоторые другие, более или менее "периферические" виды этой группы. В качестве члена рода Felis (подрод в нем) он здесь и рассматривается. С другой стороны, ирбис для рода больших кошек (Panthera) не вполне типичен и довольно равномерно комбинирует в себе признаки как больших (Panthera), так и мелких (Felis) кошек*. Он, однако, отличается как от тех, так и от других, обладает и своеобразными признаками и заслуживает, несмотря на черты промежуточного положения, выделения в самостоятельный род**.
* (Это обстоятельство может дать основание для объединения рода крупных (Panthera) и рода мелких (Felis) кошек в один род. Предлагаемое решение паллиативное, однако, оно имеет смысл на фоне тех подродовых групп, которые можно выделить в пределах pp. Felis и Panthera и относительного таксономического веса их признаков.)
** (Подробности обо всех родах приведены далее в их характеристиках.)
Таким образом, здесь принимаются, как у названных выше авторов, тоже четыре рода, однако, несколько иные и в ином составе видов. Это бдлыние кошки, Panthera Oken, 1816, ирбисы, или снежные барсы, Uncia Gray, 1854, мелкие кошки, Felis Linnaeus, 1758, и гепарды, или пардусы, Acinonyx Brookes, 1828*.
* (По степени дробления современных кошек и по самому подходу к надвидовым группировкам в семействе — мало родов и широкое использование понятия подрода — изложенные взгляды некоторых современных авторов (Эллермэн и Моррисон-Скотт, Симпсон; Гептнер) очень близки к взглядам Н. А. Северцова (1858).)
Род гепардов, Acinonyx, своеобразен и отграничен очень наглядно, хотя и менее резко, чем обычно принято думать. Он близок к мелким кошкам, Felis, и представляет собой несомненное производное этой группы. Это специализированная кошка, особенности которой заключаются в адаптации к быстрому бегу, т. е. в совершенно необычном для кошек направлении, и к добыванию относительно крупной добычи. К большим кошкам, Panthera он не имеет отношения.
Границы между родами больших кошек, Panthera, и мелких кошек, Felis, гораздо менее определенны. Значительные черты промежуточного положения между обоими родами, как указано, имеет ирбис, род Uncia, довольно явственно связывающий их. Существуют также формы настоящих мелких кошек, представляющие собой как бы уклонение в сторону ирбиса (дымчатый леопард, пума). Это, видимо, черты конвергенции, обусловленной собственными крупными размерами и крупными размерами добычи. Таким образом, Panthera, Uncia и Felis представляют собой тесную группу - последовательную цепь родов.
Что касается относительной высоты организации ("примитивности" - "прогрессивности") этих родов, степени их морфологической специализации, то решение вопроса при однотипности видов семейства и тесной связи их довольно затруднительно. Это тем более, что палеонтология дает мало указаний - все типы современных кошек появляются в сущности более или менее одновременно. По-видимому, и радиация в разных адаптивных направлениях у современных Felinae шла тоже более или менее одновременно. Согласно традиционной и наиболее распространенной точке зрения, все мелкие кошки, Felis рассматриваются как наименее специализированная ("примитивная") группа, а большие кошки, pp. Panthera и Uncia, как более специализированные ("прогрессивные"), как и гепард. Последний обычно считается специализированным в особом смысле и в ином направлении - независимом от больших кошек.
При самой общей оценке структуры черепа мелких кошек p. Felis (кроме таких, как пума и дымчатый леопард) обращают на себя внимание такие черты, как очень большой объем и вздутая форма черепной коробки, очень небольшие лицевые части, отсутствие или слабое развитие разного рода гребней и бугров, служащих для прикрепления мышц, главным образом группы жевательных. Эти признаки обычно оцениваются как ювенильные (инфантильные) и, часто справедливо считаются показателем малой специализации группы, так сказать, более нейтрального и, тем самым, более примитивного строения. Противоположного рода признаки, характеризующие череп больших кошек, расцениваются как специализированные и, тем самым, прогрессивные.
Этот традиционный подход едва ли, однако, применим к современным кошкам. Дело в том, что череп больших кошек с сильно развитыми лицевыми частями, мощными гребнями и относительно небольшой удлиненной мозговой коробкой имеет облик и строение гораздо более сходные с черепом хищников других семейств, чем череп мелких кошек. Он носит гораздо более общехищнический, чем специфически-кошачий характер и строение (рис. 19). Наиболее резко специфические именно для современной кошки черты несут как раз представители мелких кошек p. Felis s. 1. Дифференциальная характеристика современных Felidae наиболее полно выражена в черепе именно мелких кошек. Таким образом, прогрессивная специализация в пределах современных кошек шла в направлении выработки высшего кошачьего типа в мануле, а не в тигре. При всей специализации крупных кошек как кошек, в них все- таки типичных кошачьих признаков меньше, чем в мелких, и они выражены не так ярко. Мелкие кошки представляют собой кошек в наиболее полном смысле.
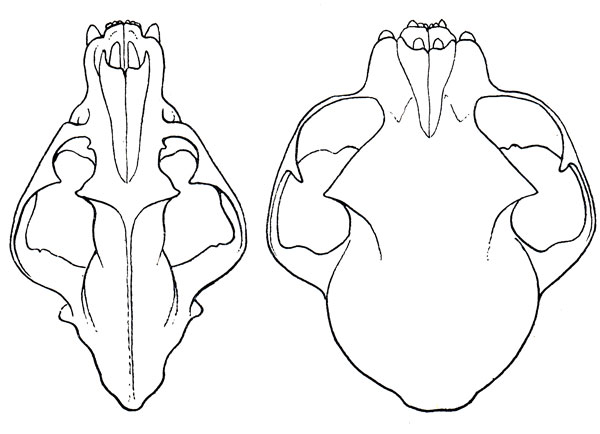
Рис. 19. Череп тигра (Panthera tigris L.). и манула (Felis manul Pall.), приведенные к одной величине; два крайних типа современных кошек. Колл. ЗММУ (рис. Н. Н. Кондакова)
При всей скудости палеонтологического материала, можно, однако, по- видимому, заключить, что у современных больших кошек (p. Panthera) черт сходства с ископаемыми формами, даже из целиком вымерших групп, больше, чему современных мелких. Ископаемые формы характерного типа Felis (silvestris, например, или lynx), не считая новейших, непосредственно связанных с современными, неизвестны или малочисленны. По-видимому, выработка вполне развитого типа современной мелкой кошки (p. Felis) - дело нового этапа эволюции семейства и подсемейства. Мысль об относительной примитивности крупных кошек, правда, без аргументации, высказали и палеонтологи (Тениус и Гофер, 1960), хотя обычно они держатся противоположного мнения (Симпсон, 1945).
Признание за современными кошками с "ювенильным" типом черепа положения наиболее прогрессивной части семейства необычно, однако это случай в группе хищных млекопитающих (Ferae) не единственный. Семейство настоящих тюленей (Phocidae), признанное всеми как наиболее специализированное и прогрессивное в отряде (подотряде), по строению черепа в такой же форме отличается от ушатых тюленей (Otariidae). Например, p. Eumetopias (сивучи) имеет особенно сильно выраженные "общехищнические" черты в черепе. Таким образом, путь развития семейства, по-видимому, есть путь цефализации.
Увеличение общих размеров в филогенетических рядах обычно рассматривается как признак специализации и прогресса. Едва ли, однако, ему можно придавать абсолютное и решающее значение; на тех же ластоногих видно, что он весьма относителен. На прогрессивность группы мелких кошек указывает и число видов в родах (Гептнер, 1965) - в роде больших кошек (Panthera) около 11,5% видов семейства, в родах гепардов (Acinonyx) и ирбисов (Uncia) по 3,5%, а в роде мелких кошек 81,5%.
Таким образом, есть достаточно оснований начинать филогенетический род современных кошек с больших кошек, Panthera, через ирбиса, Uncia, к мелким кошкам, Felis. Гепард, Acinonyx, представляет собой боковую специализированную ветвь линии мелких кошек. Названные группы характеризуют три главных направления специализации, связанных с размером добычи и способом охоты - добывание крупных зверей (большие кошки и ирбис), добывание мелких позвоночных (иногда более крупных; мелкие кошки) и добывание крупных животных догоном (гепард).
Семейство беднее видами, чем наиболее богатые в отряде виверры (Viverridae, около 75 видов) и куницы (Mustelidae около 70 видов), но все же в нем 35 видов, т. е. 14,5% от числа видов отряда и около 1 % всех млекопитающих, что больше, чем в семействе волков (Canidae, 29 видов). В роде больших кошек, Panthera, 4 вида, ирбисов, или снежных барсов (Uncia) - 1, мелких кошек (Felis) - 28 и гепардов (Acinonyx) - 1 вид.
Из четырех родов один (гепард) африкано-азиатский, один (Panthera) принадлежит главным образом Старому Свету и имеет лишь одного представителя (ягуар) в Южной и Центральной ( частично на крайнем юге Северной) Америке, один (Uncia) чисто азиатский и один (Felis) широко распространен по всему ареалу семейства как в Старом, так и в Новом Свете. Наибольшее число видов связано со Старым Светом, в частности, с Азией, наименьшее обитает в Северной Америке и довольно значительно число южноамериканских видов.
Практическое значение видов семейства в целом невелико и зависит от уровня хозяйства на данной территории. В нетронутых биоценозах и в областях мало населенных человеком виды семейства безразличны или имеют не-большое практическое значение как пушные или в торговле живыми животными для зоопарков. В более густо населенных областях кошки крупных видов нередко убивают скот или, добывая диких копытных, вредят охотничьему хозяйству. Отдельные звери (тигр, лев, иногда барс, ягуар) в отдельных местностях иногда при известных условиях становятся в большей или меньшей мере опасными для людей.
Некоторые крупные виды (лев, тигр, барс, пума, ягуар) представляют собой очень ценных охотничьих спортивных животных. Одновременно они осуществляют качественную селекцию, поддерживая благополучие популяций копытных. В культурной обстановке вредят охотничьему хозяйству и некоторые кошки мелких видов, уничтожая охотничьих птиц (фазан), зайцев или молодняк копытных. Многие виды питаются почти исключительно мелкими грызунами, однако роли существенных регуляторов их численности не играют. Некоторые виды местами находятся под охраной вообще или в заповедниках. Один (F. silvestris) существует также в виде настоящей домашней формы (домашняя кошка, "F. catus L.")*, два вида приручают и дрессируют для охоты (гепард - газели, каракал - птицы), но домашними считаться не могут.
* (Современное обозначение Felis silvestris L.— forma catus. О происхождении домашней кошки см. при описании степной кошки (F. s. caudata).)
В фауне СССР представлены все 4 рода и 11 видов, т. е. около 31-32% видов семейства. Они составляют около 10% видов нашей фауны.
Ареал покрывает почти всю территорию страны на север до границы леса. Большинство видов связано с южными областями страны, одни (рысь) занимают также широкие области ее северной части. Формы лесов, гор, степей и пустынь. Имеют сравнительно небольшое значение как пушные виды или животные для зоопарков. Численность некоторых в результате преследования очень низка, и они находятся на грани исчезновения. Один вид (тигр) особенно резко сократил свою численность и ареал и находится под охраной, два других (гепард, каракал) - под запретом. (В. Г.).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'