Отряд Прямокрылые (Orthoptera) (Ф. Н. Правдин)
Среди насекомых с неполным превращением прямокрылые занимают особое место. Это очень крупная группа, включающая свыше 20 000 видов, из которых более 700 встречаются в СССР. Прямокрылые широко распространены по всему земному шару - от тропиков до Заполярья -и отличаются большим разнообразием морфологических структур и физиологических приспособлений к весьма разнообразным условиям среды. Однако не только этим привлекают к себе внимание прямокрылые насекомые. С давних времен человек узнал среди них многих врагов, беспощадно уничтожающих плоды его труда. Слово "саранча" до сих пор вызывает представление о большой надвигающейся опасности.
К прямокрылым относят насекомых с удлиненным телом, грызущими ротовыми органами и характерным строением груди, летательного аппарата и задних конечностей. Голова у них с крупными, обычно овальными, сложными глазами и большей частью 3 глазками; находящиеся на ней усики могут быть длинными, превышающими длину тела (кузнечиковые, сверчковые), или короткими - короче половины тела (триперстовые, саранчовые). На этом различии в строении усиков основано деление прямокрылых на два подотряда - длинноусых и короткоусых.
Специфично строение груди прямокрылого: в ней сильно развита и подвижна переднегрудь, причем боковые части переднеспинки свешиваются вниз, образуя широкие лопасти, прикрывающие переднегрудь с боков. Остальные два отдела груди плотно слиты друг с другом. Крылья по большей части развиты нормально,хотя есть формы с укороченными крыльями и даже совсем бескрылые. Крылья передней пары более плотные и узкие и представляют собой надкрылья. Задние крылья, или просто крылья, широкие, перепончатые с хорошо развитым продольным жилкованием. При посадке насекомого они веерообразно складываются и прикрываются надкрыльями. Задние ноги прыгательного типа с утолщенными и удлиненными бедрами и длинными голенями. Поэтому прямокрылых иногда называют прыгающими насекомыми (Saltatoria). Брюшко 10-члениковое, удлиненное, с церками; снизу оно кажется 8- или 9-члениковым, так как один или два стернита оказываются редуцированными. Прямокрылые могут издавать и воспринимать звуки, так как имеют особые звуковые и слуховые аппараты, структура которых различна в разных подотрядах.
Подотряд Длинноусые прямокрылые (Dolichocera, или Ensifera)
Длинноусые прямокрылые отличаются прежде всего длинными щетинковидными антеннами, превышающими обычно длину тела; орган слуха, в тех случаях когда он имеется, расположен на голенях передних ног; яйцеклад всегда длинный. В подотряде различают два надсемейства - кузнечиковых и сверчковых.
Кузнечиковых легко отличить от остальных прямокрылых по строению их лапок и характерной форме яйцеклада самки: лапки на всех ногах у них 4-члениковые, а яйцеклад длинный, обычно сжатый с боков, изогнутый в виде серпа или сабли или же мечевидный. Надкрылья и крылья у кузнечиковых большей частью развиты хорошо и тогда достигают вершины брюшка или даже заходят за него. При этом в состоянии покоя надкрылья сложены по длине тела крышеобразно и прикрывают спрятанные под ними крылья. У некоторых видов надкрылья и крылья в разной степени укорочены, есть и совершенно бескрылые формы.
Звуковой аппарат у кузнечиковых располагается на надкрыльях. В основании правого надкрылья находится так называемое "зеркальце" в виде округлой тонкой прозрачной перепонки, окруженной толстой жилкой, образующей рамку. На левом надкрылье тоже имеется перепоночка в виде зеркальца, но оно обычно непрозрачное, матовое и довольно плотное (рис. 149). Окружающая его жилка толстая и снабжена на нижней стороне надкрылья мелкими зубчиками. Эта характерная жилка называется стридуляционной; она выполняет роль смычка, в то время как "зеркальце" служит при стрекотании резонатором. Стрекотание осуществляется следующим путем. Левое надкрылье в спокойном состоянии всегда лежит над правым. Во время стрекотания кузнечик несколько приподнимает и раздвигает надкрылья, а затем приводит их в вибрирующее движение из стороны в сторону, в результате чего зубчики "смычка" трутся о рамку "зеркальца", находящегося на правом надкрылье. Благодаря "зеркальцу" звук значительно усиливается и потому слышен издалека. У каждого вида кузнечиков издаваемые им звуки специфичны. В преобладающем большинстве случаев звуковой аппарат имеется только у самцов, однако есть виды кузнечиков, у которых стрекочут и самки.
Слуховой аппарат находится на голенях передних ног; снаружи его легко обнаружить благодаря двум овальным перепонкам, расположенным по обеим сторонам голени и выполняющим функцию барабанных перепонок (рис. 149). В одних случаях эти перепонки открыты, хотя и помещаются в небольшом углублении; у большинства же видов они прикрыты хитиновыми покрышками, так что снаружи видны только две продольные щели, ведущие к перепонкам. Внутренняя часть слухового аппарата имеет сложную структуру, состоящую из окончаний нервов, чувствительных клеток, мышц и двух ветвей трахей, из которых каждая подходит к своей барабанной перепонке. Благодаря давлению воздуха в трахеях перепонки всегда натянуты.
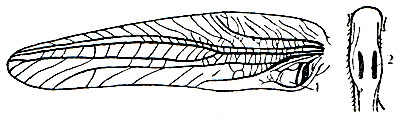
Рис. 149. Левое переднее крыло и верхняя часть голени передней ноги зеленого кузнечика: 1 - звуковой аппарат; 2 - щели, ведущие к слуховому органу
В летнее время стрекотание кузнечиков раздается повсюду: на лугах, опушках лесов, лесных полянах и в садах. Оно не смолкает даже вечером и ночью. Однако обнаружить самих стрекочущих кузнечиков далеко не такая уж легкая задача: слишком хорошо они оказываются замаскированными в зелени кустов или в толще густой травы. Эту маскировку обусловливает сочетание формы тела и его защитной окраски. Виды, живущие в траве, часто обладают вытянутым телом и длинными узкими надкрыльями. Распространенный на Малайском архипелаге кузнечик Elimaea poaefolia имеет настолько удлиненное тело, что напоминает стебель растения, на котором он сидит. Среди тропических кузнечиков большое количество видов обнаруживает сходство с листьями деревьев. Это сходство достигается благодаря сильному расширению надкрылий, а также их специфическому жилкованию. В зависимости от окраски они имитируют или здоровые, или отмирающие и мертвые листья (рис. 150). Так, например, на листоподобных крыльях Cycloptera elegans имеются бурые пятна, напоминающие повреждения листьев паразитическими грибами. Еще более своеобразна форма и окраска надкрылий у видов Tanusia: имеющаяся на них пятнистость имитирует начало разложения листа, а неровные края создают впечатление, что лист объеден или обломан. Сильно объеденными кажутся листоподобные крылья у акридоксены (Acridoxena hewaniana). Некоторые индо-малайские кузнечики, живущие на деревьях, весьма успешно имитируют лишайники. Так, например, яванская сатрофилия (Satrophyllia femorata), сидя неподвижно с вытянутыми вперед усиками и передними ногами на ветке дерева, совершенно сливается с общим фоном покрывающих эту ветку лишайников.
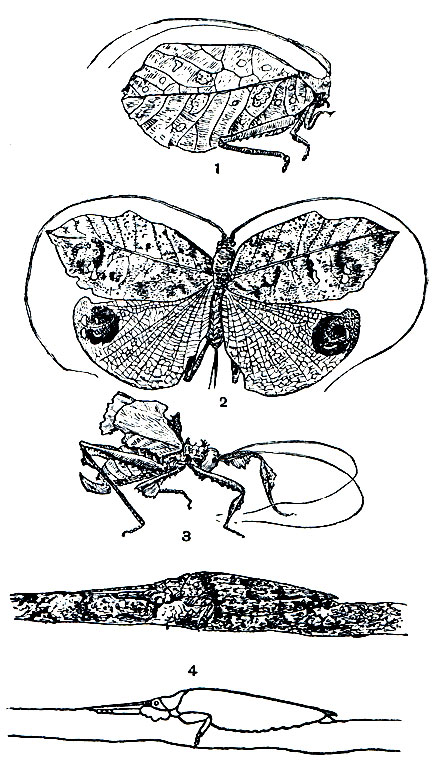
Рис. 150. Покровительственное сходство с частями растений у кузнечиков: 1 - Cycloptera elegans; 2 - Tanusia corrupta; 3 - Acridoxena hewaniana; 4 - Satrophyllia femorata (имитация лишайника)
Хотя у кузнечиков, так же как у всех прямокрылых, задние ноги прыгательного типа, т. е. с утолщенными бедрами, обычно они передвигаются по веткам или в траве, быстро ползая при помощи всех шести ног. Если же кузнечика потревожить, он упирается концами голеней задних ног, а затем быстро распрямляет ноги и делает прыжок вперед. Крылатые формы при этом могут распускать крылья, которые в данном случае служат парашютом и позволяют значительно увеличить прыжок.
Очень своеобразен способ оплодотворения у кузнечиковых: при спаривании самец подвешивает к концу брюшка самки сперматофор. Сперматофор имеет довольно сложное строение: он состоит из основной части - флакона и дополнительной - сперматофилакса (рис. 151). Флакон имеет узкую шейку и две укрепляющие лопасти; сверху он покрыт оболочкой, а его внутренняя полость, в которой содержатся сперматозоиды, разделена перегородкой на две самостоятельные части. Сперматофилакс представляет собой объемистуюлипкую массу. При спаривании самец вводит в половое отверстие самки шейку флакона; сам флакон и сперматофилакс остаются снаружи. После спаривания самка начинает поедать сперматофилакс; благодаря вязкой консистенции этого образования процесс поедания протекает медленно и может длиться несколько часов. За это время сперма постепенно перетекает из флакона в яйцевод, после чего самка поедает и флакон. Весь этот оригинальный процесс имеет большое биологическое значение. Подвешенный к брюшку самки сперматофор, снабженный утяжеляющим его сперматофилаксом, затрудняет ее движение и мешает откладке яиц и повторному спариванию. Поэтому удаление его совершенно необходимо. В то же время этот процесс должен протекать медленно, иначе сперма не успеет перелиться из флакона в яйцевод.
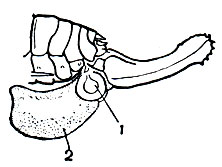
Рис. 151. Конец брюшка оплодотворенной самки кузнечика Isophya acuminata: 1 - флакон сперматофора; 2 - сперматофилакс
Характер откладки яиц во многом зависит от образа жизни того или иного вида кузнечиков. Большинство плотоядных видов, питающихся другими насекомыми, или видов, у которых пища смешанная - животная и растительная, откладывают яйца в землю, погружая в нее свой яйцеклад. В одних случаях яйца откладываются поодиночке, в других - небольшими кучками (по 5-10 шт.), скрепленными застывающими выделениями придаточных половых желез.
У растительноядных видов, более тесно связанных в своей жизни с растениями, яйца чаще всего откладываются или на поверхность растения, или внутрь какого-нибудь его органа. Так, например, самка четырехточечного пластинокрыла (Phaneroptera quadripunctata) откладывает яйца в паренхиму листа (табл. 29, 1). Приступая к откладке, она садится верхом на край листа, сжимая его с боков передними и средними ногами. Затем самка слегка надгрызает край листа, сильно подгибает брюшко и, придерживая челюстями основание яйцеклада, вводит его створки в паренхиму в том месте, где сделан надрез. Яйцеклад погружается почти полностью, и на дно образовавшегося в ткани кармашка откладывается яйцо.
Обыкновенный пластинохвост (Leptophyes albovittata, табл. 29, 2) откладывает яйца в щели старой древесины столбов и заборов, а другой вид этого рода (L. punctatissima) - в трещины коры деревьев и кустарников. Шиповатый пластинокрыл (Tylopsis lilifolia, табл. 29, 4) помещает яйцо на стебле злаков, погружая его за влагалище листа. Обитающий на сильно увлажненных, заливаемых водой лугах короткокрылый мечник (Conocephalus dorsalis) для откладки яиц выбирает стебли тростника или ситника с хорошо развитой сердцевиной, внутрь которых он вводит свой яйцеклад (табл. 29, 3).У наших видов кузнечиковых яйцекладка обычно происходит во второй половине лета, яйца имеют удлиненноовальную, цилиндрическую или уплощенную форму. Отложенные яйца перезимовывают, личинки вылупляются весной и в течение жизни линяют 4-6 раз. Зачатки крыльев появляются у личинок уже после первой линьки в виде оттянутых вниз и назад задних нижних углов среднеспинки и заднеспинки; после третьей линьки зачатки крыльев располагаются на спине, принимают треугольную форму и на них появляются продольные жилки. После последней линьки происходит окрыление.
Как правило, личинки и нимфы кузнечиковых похожи на взрослых представителей данного вида, отличаясь от них размерами и отсутствием нормально развитых крыльев. Однако известны случаи, когда личинки сильно отличаются по внешнему виду от взрослых. Эти отличия могут заключаться в структуре переднеспинки, наличии на теле шиловидных выростов, в окраске и ряде других признаков. Наиболее резкие отличия наблюдаются при трансформативной мимикрии. Так называют явление, при котором для личинки характерна мимикрия, отсутствующая у имаго. В этом отношении особенно интересны некоторые африканские и индо-малайские листоподобные кузнечики, личинки которых своей внешностью и повадками имитируют либо муравьев, либо жуков-скакунов. У суданского кузнечика (Eurycorypha fallax) личинки живут открыто на листьях и цветах кустарников, встречаясь совместно с имитируемыми ими видами муравьев. Эта имитация достигается характером распределения на теле темного и зеленого пигментов. Хотя плотное и толстое тело личинки и не похоже на тело муравья, однако на нем по светлому зеленому фону брюшка выведен темный рисунок, воспроизводящий узкую "талию" и вздутое брюшко, типичные для муравьев (рис. 152). У малайского кузнечика (Leptoderes ornatipennis) личинки младших возрастов своей металлически синей окраской и красными бедрами сходны с жуком-скакуном (Collyris tuberculata). Оба вида встречаются вместе, причем характер прыжков у них тоже очень сходен.
Рис. 152. Подражание муравью у личинки кузнечика Eurycorypha fallax
Кузнечиковые представляют собой группу древнего происхождения. Их предками, по-видимому, были так называемые первичные прямокрылые (Protorthoptera), существовавшие в каменноугольном периоде. Они имели много черт сходства с кузнечиковыми, отличаясь от них уплощенным телом и задними конечностями непрыгательного типа. В пермском периоде появляются уже типичные представители кузнечиковых (рис. 153). В третичном периоде формируются некоторые современные роды, такие, как зеленые кузнечики (Tettigonia), серые (Decticus) и скачки (Platycleis).
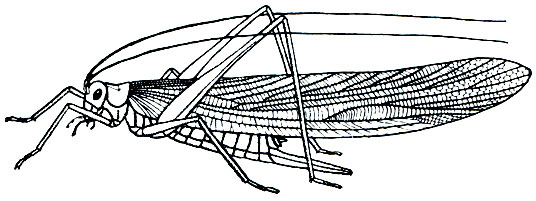
Рис. 153. Кузнечик Pinegia longipes из нижних отложений пермского периода (Урал)
В настоящее время около 70% кузнечиковых распространено в тропических и субтропических странах. На территории Советского Союза обитает свыше 170 видов, относящихся к 3 семействам.
К семейству рафидофорид (Rhaphidophoridae) относятся совершенно бескрылые кузнечики с тонкими и длинными усиками и длинными ногами. На голенях задних ног сидят сильно удлиненные шпоры, органы слуха отсутствуют. Преобладающее большинство рафидофорид обитает в тропиках. Типичным выходцем из тропических стран является оранжерейный кузнечик (Tachycines asynamorus) - небольшого разхмера (13-18 мм), буровато-серый в темных пятнах и в шелковистом налете из мельчайших густых волосков, с очень длинными тонкими усиками и длинными ногами. Родина этого интересного насекомого - Центральный Китай. Отсюда он с растениями завезен в города Европы и Северной Америки, где встречается теперь в оранжереях. Ведет преимущественно ночной образ жизни, на день прячась в укрытия. Может делать большие прыжки - до 40 см в высоту и 1,5 м в длину. Питается смешанной пищей - как частями растений, так и находящимися на них мелкими насекомыми. Вредит декоративным растениям: цикламенам, папоротникам и др. Самка в течение жизни откладывает в землю от 150 до 900 яиц, из которых через 3-4 месяца вылупляются личинки. Развитие последних протекает в течение 7 месяцев и притом с необычно большим для кузнечиков числом линек - до 11.

Рис. 154. Рафидофориды: 1 - оранжерейный кузнечик (Tachycines asynamorus); 2 - кавказский пещерник (Dolichopoda euxina)
На Черноморском побережье Кавказа и в Западном Закавказье в пещерах обитает кавказский пещерник (Dolichopoda euxina) - буровато-желтого цвета, с длиннейшими усиками, в четыре раза превышающими длину тела. Это представитель очень древней первичной тропической фауны Кавказа. Именно пещерный образ жизни позволил сохраниться этому кузнечику до наших дней, несмотря на те сильные изменения в природной обстановке, которые происходили на территории Кавказа.
Семейство брадипорид (Bradyporidae) - обитатели сухих районов Евразии и Северной Африки. Наиболее характерным признаком, позволяющим отличить, этих кузнечиков, является расположение усиков на шаровидно выпуклой: голове; они прикрепляются ниже глаз или на уровне их нижнего края. Надкрылья всегда сильно укорочены и часта бывают скрыты под переднеспинкой. На голенях передних ног всегда есть орган слуха.
Из наших представителей семейства весьма оригинален по своему виду и биологическим особенностям седлоносец, или виноградный эфиппигер (Ephippiger ephippiger, табл. 29, 10). Это сравнительно небольшой по размерам (20-30 мм) кузнечик, желтоватой или голубовато-зеленой окраски с сине-черным затылком. Свое название седлоносец получил из-за характерного строения переднеспинки: она резка седловидной формы с сильно приподнятой задней частью. Ржаво-рыжие надкрылья укорочены, почти совсем прикрыты переднеспинкой; крыльев нет. Но что особенно интересно - это наличие звукового аппарата у обоих полов. Самки, так же как: и самцы, способны стрекотать. Седловидная переднеспинка при стрекотании выполняет роль рупора, усиливая звук.
Еще сравнительно недавно в ковыльно-типчаковых степях Предкавказья, Украины и центральных районов России часто встречался степной толстун (Bradyporus multituberculatus) - крупный бескрылый кузнечик, длиной 6-7 см, с грузным телом, окрашенным в бронзово-черный цвет, с двумя продольными желтыми полосами на брюшке (табл. 29, 12). Распашка степей значительно сократила ареал этого вида. Сейчас это реликт степной фауны, сохранившийся только в Сальских степях и еще в некоторых местах Предкавказья, где уцелели целинные степные участки.
К семейству настоящих кузнечиков (Tettigoniidae) относится преобладающее большинство видов наших кузнечиков. От рафидофорид они отличаются наличием на голенях передних ног органа слуха, а от брадипорид - тем, что у них усики прикрепляются выше уровня нижнего края глаз.
Широкой известностью у нас пользуются зеленые кузнечики (Tettigonia)- довольно крупные, почти одноцветные с длинными усиками и длинными, узкими, довольно мягкими надкрыльями, под которыми в состоянии покоя спрятаны хорошо развитые прозрачные крылья; самки с длинным мечевидным яйцекладом, заостренным на вершине и немного загнутым вниз (табл. 29, 6-8). В Палеарктике обитает свыше 10 видов рода Tettigonia, из которых 4 встречаются в СССР. Наиболее широко у нас распространен зеленый кузнечик (T. viridissima): его можно найти во всех ландшафтных зонах, не заходит он только на север. В лесной зоне, начиная с конца июля и вплоть до поздней осени, зеленый кузнечик часто встречается по краям лугов в траве, а на опушках лесов и в садах - на кустах и деревьях, искусно маскируясь в зеленой листве. В пустынях он тяготеет к речным долинам и к опушкам тугайных лесов; в Средней Азии высоко проникает в горы, придерживаясь ущелий с сочной травянистой растительностью. Питается он мелкими насекомыми - различными двукрылыми, мелкими бабочками, их гусеницами и др. При содержании в неволе проявляет склонность к каннибализму, поедая более слабых особей и личинок своего вида. В то же время зеленый кузнечик может питаться и растительной пищей. Так, например, он иногда вредит виноградникам, выгрызая почки, объедая цветы и листья. Известны случаи повреждения им листьев чая, цитрусовых и других растений.
Стрекотание зеленого кузнечика можно слышать днем, в послеобеденное время и вечером, не смолкает оно и в темноте вплоть до 2-3 часов ночи. В утренние часы зеленый кузнечик принимает "солнечную ванну". Он лежит на боку, подставляя свое тело падающим на него солнечным лучам и время от времени переворачиваясь на другую сторону. Спаривание у зеленого кузнечика продолжается около 45 минут. Спустя 15 минут самец опять начинает стрекотать. Поедание самкой сперматофора может длиться 15 часов. Яйца она откладывает в землю по одному, располагая их так тесно, что они оказываются склеенными друг с другом по 2, 3 или 4. Процесс откладки яиц, по-видимому, довольно трудный, о чем можно судить по тому, что самка при этом порывисто дышит, а число откладываемых ею яиц достигает 70-100. Яйца лежат в почве до весны, когда из них начинают вылупляться личинки. Они тоже зеленого цвета с коричневой или черноватой полосой на спинной стороне.
Похожи на зеленого кузнечика как по внешнему виду, так и по образу жизни два других встречающихся в Европейской части вида этого рода - певчий кузнечик (Tettigonia cantans) и хвостатый кузнечик (T. caudata). Певчий кузнечик обитает в лесной зоне. Отличается он более короткими и широкими надкрыльями, не заходящими за вершины бедер задних ног. Хвостатый кузнечик встречается на юге Европейской части СССР, доходя до Западной Сибири. У него такие же длинные крылья, как у зеленого, заходящие за вершины бедер задних ног, но яйцеклад всегда выдается за вершину надкрылий. Четвертый встречающийся в СССР вид рода Tettigonia - уссурийский зеленый кузнечик (Т. ussuriana) распространен в Уссурийском крае, населяя область смешанных и широколиственных лесов Дальнего Востока.
Такое же широкое распространение имеет, как и зеленый кузнечик, и так же хорошо известен наш серый кузнечик (Decticus verrucivorus, табл. 29, 9). Это тоже крупный вид, окрашенный в светлый или темно-зеленый цвет с большим количеством бурых пятен, нередко сплошь бурый с еще более темными пятнами. Серый кузнечик не выносит затенения и поэтому обычно селится на ярко освещенных солнцем сухих лугах, лесных полянах и других открытых местах. Если его потревожить, он невысоко взлетает и затем снова прячется в траве. Серый кузнечик тоже всеяден и в неволе склонен к каннибализму. Под Москвой стрекотание начинается во второй половине июня. Самец приступает к нему спустя неделю после окрыления. Самка начинает откладку яиц через неделю после спаривания. Число откладываемых в землю яиц в среднем около 50.
Весьма своеобразна степная дыбка (Saga pedo, табл. 29, 11). Это один из самых крупных наших кузнечиков, длиной 6-8 см, почти или совсем бескрылый, с удлиненным, стройным телом зеленого или желтоватого цвета, с двумя светлыми полосами, проходящими по нижнему краю переднеспинки и по бокам всех брюшных сегментов. Дыбка - типичный представитель степной фауны. В СССР она встречается в степях Европейской части и Северного Казахстана и в прилегающих частях Западной Сибири. По своим повадкам степная дыбка скорее напоминает богомола, чем кузнечика. Она часами может неподвижно, с распростертыми ногами сидеть в траве или на кусте в ожидании своих жертв, которых она ловит передними ногами. Ее жертвами являются довольно крупные насекомые - саранчовые, сверчки, жуки и клопы. Любопытна еще одна биологическая особенность этого насекомого: самцы чрезвычайно редки, и поэтому размножение происходит, очевидно, партеногенетическим путем.
Сверчковые, в отличие от кузнечиковых, имеют 3-члениковые лапки, длинные гибкие церки и тонкий прямой яйцеклад (у самок), который на конце обычно копьевидно расширен. Только у медведки (сем. Gryllotalpidae) яйцеклад отсутствует.
Надкрылья у сверчковых в покое лежат плоско на спинной поверхности тела, причем левое всегда прикрыто правым. С этим связаны особенности звукового аппарата, который хотя и сходен по структуре с звуковым аппаратом кузнечиков, но отличается от последнего большей сложностью и иным расположением компонентов: стридуляционная жилка, выполняющая функцию смычка, располагается на правом надкрылье, а жилка, о которую она трется, воспроизводя звук,- на левом. Вообще весь этот аппарат развит сильнее и занимает значительно большую площадь на надкрыльях, чем у кузнечиков. Имеется он только у самцов, причем у каждого вида он отличен по строению и воспроизводимым звукам. Различают 6 типов стрекотания сверчков: призывное, брачное, стрекотание после спаривания, агрессивное стрекотание при встрече самцов, стрекотание, связанное с внезапным исчезновением самки, и у некоторых видов - "опознавательный" сигнал. Однако "репертуар" вида состоит обычно из меньшего количества песен. Наиболее древний тип песни, вероятно, брачное пение.
У некоторых видов сверчковых как надкрылья, так и крылья могут недоразвиваться или даже совсем отсутствовать. Нередки случаи, когда степень развития летательного аппарата неодинакова у различных особей одного и того же вида и пола. В тех случаях, когда задние крылья развиты нормально, они складываются веерообразно и в покое в виде жгутов выдаются из-под надкрылий.
Слуховой аппарат сверчковых такого же типа, как у кузнечиков, и также располагается на голенях передних ног.
В СССР встречается около 45 видов сверчковых, большинство которых относится к семейству сверчков (Gryllidae). Представителем этого семейства может служить широко распространенный в центральных и южных частях Европы, в Средней Азии и в Северной Африке полевой сверчок (Gryllus campestris). Это один из самых крупных наших видов, длиной 2-2,6 см. Он черного цвета с бурыми надкрыльями, при основании которых находится по одному оранжевому пятну; бедра задних ног внизу ярко-рыжие. На юге его стрекотание можно слышать уже в начале лета. Поющий самец сидит у входа в свою норку; если его потревожить, он сейчас же прячется в ней. Норка представляет собой наклонный ход, не шире пальца и примерно в палец длиной. Вход в норку закрыт пучком травы. Если к норке подходит другой самец, то между ними начинается драка. Оба бросаются друг на друга, ударяются своими толстыми головами, и каждый старается укусить своего противника. Побежденный противник поедается победителем, несмотря на то что обычно полевой сверчок питается растительной пищей.

Рис. 155. Полевой сверчок (Gryllus campestris)
Стрекотание самца привлекает самку. При спаривании самец подвешивает к брюшку самки сперматофор, который очень похож на сперматофор кузнечиков, но отличается отсутствием сперматофилакса. Через несколько дней после спаривания самка приступает к откладке яиц, для чего она погружает яйцеклад в землю, держа его совершенно отвесно. Одна самка может отложить до 500-600 яиц. Примерно через месяц из них вылупляются молодые сверчки, очень похожие на взрослых и отличающиеся в основном мелкими размерами и отсутствием крыльев. После второй линьки они начинают копать маленькие ямки; до начала зимы происходит еще одна или две линьки. Последняя линька, после которой формируется взрослый сверчок, происходит уже весной, приблизительно в мае.
В домах может встречаться домовый сверчок (Gryllus domesticus). Он меньшего размера, длиной 1,6-2 см, соломенно-палевый с коричневыми полосками. Днем он прячется в щелях и из этих убежищ выползает большей частью только ночью в поисках пищи, состоящей из различных отбросов растительного происхождения. Самцы стрекочут вечером и ночью. Развитие домового сверчка протекает в течение года. По-видимому, у него нет четкой приуроченности периода откладки яиц к какому-то определенному сезону, так как в любое время года можно найти разные стадии его развития. Вне поселений человека, на воле, домовый сверчок встречается только в пустынях.
Очень своеобразны стеблевые сверчки (семейство Oecanthidae) с желтоватым или зеленоватым нежным телом, плоскими широкими надкрыльями, почти сплошь занятыми органом стрекотания, и тонкими длинными ногами. Это южные формы, связанные с ландшафтами степей и пустынь. Большую часть времени стеблевые сверчки проводят на растениях, которыми и питаются. Днем они прячутся под листьями; громкое стрекотание их самцов раздается вечером и ночью. Самки откладывают яйца в стебли растений; здесь яйца остаются лежать на всю зиму, и только весной из них вылупляются молодые сверчки. Откладка яиц в стебли портит молодые побеги, которые после этого быстро засыхают. Встречаясь на культурных полях, стеблевые сверчки могут приносить вред сельскохозяйственным растениям. Так, в южной части Уссурийского края дальневосточный стеблевой сверчок (Oecanthus longicaudus) вредит сое и сахарной свекле; обыкновенный стеблевой сверчок (O. pellucens) повреждает виноградники и табачные плантации в степных районах, а туранский стеблевой сверчок (O. turanicus) в годы массового размножения вредит хлопчатнику, кунжуту и другим культурам в Закавказье и Средней Азии.
Совершенно лишены крыльев и звукового аппарата очень мелкие, длиной всего 2-5 мм, с яйцевидным или округлым телом и большими церками сверчки-муравъелюбы (семейство Myrmecophilidae), обитающие под камнями в гнездах муравьев. В СССР они представлены 5 видами. Наиболее хорошо изучен образ жизни у обыкновенного муравъелюба (Myrmecophilus acervorus). Этого сверчка можно встретить в подземных муравейниках различных муравьев. Питается он здесь не только остатками запасов, делаемых хозяевами, но может уничтожать их яйца и личинок. Характерно, что самки способны размножаться партеногенетически. Откладка яиц производится в землю и протекает с весны до осени. В течение сезона одна самка может отложить до 49 яиц.
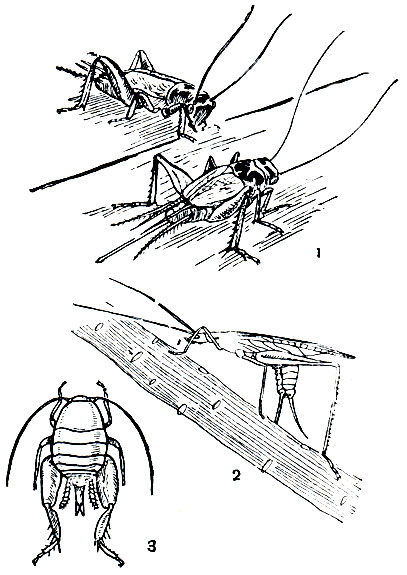
Рис. 156. Сверчки: 1 - домовый (Gryllus domesticus); 2 - обыкновенный стеблевой (Oecanthus pellucens); 3 - обыкновенный муравьелюб (Myrmecophilus acervorus)
Подземный образ жизни ведут медведки (семейство Gryllotalpidae), которые резко отличаются от всех остальных сверчковых очень большой переднеспинкой, сравнительно короткими усиками, лишь немного заходящими за переднеспинку, и сильно измененными передними ногами, отлично приспособленными к копанию и передвижению в земле. По совершенству приспособления передняя конечность медведки не уступает передней ноге крота; отсюда происходит латинское название этого животного: Gryllotalpa - "сверчок-крот". В копательной конечности медведки сильно расширены бедро и голень, лапка же укорочена и прикрепляется сбоку голени, на которой имеется 4 черных зубца. Из встречающихся в СССР 3 видов медведок широко распространена в Палеарктике обыкновенная медведка (Gryllotalpa glyllotalpa).
Ее крупное тело, длиной 3,5-5 см, сверху темно-бурого цвета, а снизу буро-желтое, кажется шелковистым благодаря густому покрову из мелких золотистых волосков; крылья развиты нормально и в спокойном состоянии выступают из-под укороченных надкрылий в виде загнутых книзу жгутов.
В естественных условиях медведка селится обычно в поймах рек, где почва всегда достаточно увлажнена. Однако ее часто можно находить на огородах и в садах, и тут она приносит большой вред, повреждая корневую систему многих культурных растений, в том числе кукурузы, картофеля, огурцов, а в Средней Азии - хлопка и риса.
Днем медведки держатся под землей, а вечером с наступлением темноты выходят на поверхность земли, причем иногда летят на свет. Самцы стрекочут в темноте, хотя могут издавать звуки и днем, находясь под землей, но эти звуки значительно более короткие и слабые по сравнению с длинными и резкими ночными призывными трелями. Характерно, что стрекотать могут и самки.
Пища у медведок смешанная, при содержании в неволе они охотно питаются как подземными органами растений, так и животными, например дождевыми червями, личинками божьих коровок и стрекозами.
В период размножения медведки устраивают сложную систему ходов, расположенных вблизи корневой системы диких и культурных растений. На образующихся в результате этой деятельности плешинах на глубине 5-10 см от поверхности медведка в середине лета устраивает крупные маточные камеры, имеющие вид шарообразного гнезда, диаметром 5-10 см. В середине лета в таком гнезде можно найти до 600 яиц или вылупившихся из них молодых медведок. В Европейской части СССР медведка распространена почти по всей территории, доходя на севере до Ленинградской области. Поэтому длительность постэмбрионального развития у нее неодинакова в разных районах. На юге она, по-видимому, развивается в течение года, тогда как на севере развитие затягивается до 2-2 1/2 лет. За это время может происходить до 8-9 линек. Личинки младших возрастов очень юрки и хорошо прыгают. В летний период медведки роют свои ходы неглубоко под землей, но на зиму как личинки, так и взрослые копают длинные ходы, расположенные под углом 45-60° к поверхности, и уходят на большую глубину - до 25 (личинки) и даже до 60 см (взрослые). Интересна еще одна биологическая особенность медведки: так как пойменные луга, где она обычно обитает, весной заливаются водой, насекомое хорошо приспособилось к плаванию, и может свободно переплывать залитые водой пространства.

Рис. 157. Обыкновенная медведка (Gryllotalpa gryllotalpa) (1) и обыкновенный триперст (Tridactylus variegatus) (2)
Подотряд Короткоусые прямокрылые (Brachycera, или Caelifera)
В отличие от длинноусых прямокрылых представители этого подотряда обладают относительно короткими антеннами, никогда не достигающими длины тела. В подотряде различают два надсемейства - триперстовых и саранчовых.
Триперстовыми называют мелких насекомых, длиной 4-9 мм, напоминающих по внешнему виду маленьких медведок. Это сходство выражается в однотипной конфигурации тела и в строении передних ног, которые, так же как у медведок, приспособлены к копанию. Однако достаточно посмотреть на этих насекомых немного повнимательней, чтобы убедиться в том, что триперсты относятся к другой систематической группе, а именно к короткоусым прямокрылым. Об этом свидетельствует строение головы, которая несет четковидные усики, состоящие всего из 10 члеников.
Надкрылья у триперстов короткие, роговые; крылья могут быть или длиннее, или короче надкрылий, с непрозрачным передним краем и многочисленными радиально расходящимися жилками. Весьма своеобразно строение задних ног: они прыгательные, с широкими уплощенными бедрами и тонкими, слегка изогнутыми голенями. По обеим сторонам голени располагаются шипики, имеющие вид лопастинок, а на ее вершине сидят две короткие и две длинные шпоры. Лапка задней ноги состоит всего из одного более или менее вытянутого пальцевидного членика, располагающегося между длинными шпорами голени, что и дало основание называть этих насекомых "триперстами". Брюшко 9-10-члениковое, несет на вершине две пары удлиненных придатков, из которых верхняя, 2-членистая, представляет собой церки. У триперстов нет ни звукового, ни слухового аппарата; у самок наших видов отсутствует также и яйцеклад.
В СССР встречается всего 4 вида, относящихся к одному роду (Tridactylus), распространенному преимущественно в субтропиках. Наши виды известны с Кавказа, из Средней Азии и Казахстана, а также из степных районов Европейской части СССР и Западной Сибири. Обитают они обычно на песчаных берегах рек и других водоемов. В солончаковых пустынях встречаются около западин, где долго сохраняется вода.
Примером может служить обыкновенный триперст (Tridactylus variegatus), широко распространенный в степях и пустынях Евразии. Его можно встретить на песчаных отмелях, где с помощью передних ног и челюстей он выкапывает в сыром песке норки. В таких условиях триперстов можно находить иногда в массе. Они быстро перепрыгивают с одного места на другое на освещенной солнцем поверхности песка или в разреженном травянистом покрове. Триперсты - хищники, питающиеся мелкими насекомыми и дождевыми червями.
Саранчовые составляют самую многочисленную группу среди прямокрылых. От кузнечиков и сверчков они легко отличаются короткими усиками, не превышающими длины половины тела, коротким яйцекладом самки и весьма специфичными органами звука и слуха. В отличие от триперстовых у саранчовых лапки задних ног 3-члениковые, число члеников усиков может доходить до 28, церки не членистые, в виде конических выступов, а у самки всегда имеется яйцеклад.
Подобно кузнечикам и сверчкам саранчовые - признанные музыканты в мире насекомых. Чтобы убедиться в этом, достаточно летом выйти на любой луг или опушку леса; в дневное время их стрекотание явно забивает "пение" кузнечиков в силу того, что одновременно может стрекотать сразу большое число экземпляров.
Звуковой аппарат саранчовых располагается на бедрах задних ног и надкрыльях. Обычно вдоль внутренней поверхности бедра тянется длинный ряд бугорков или головчатых шишечек, а одна из жилок надкрылья оказывается толще остальных. Быстро двигая бедром, насекомое проводит этими бугорками по жилке, в результате чего раздаются отрывистые стрекочущие звуки. Эти звуки можно воспроизвести даже на мертвом насекомом, если быстро двигать его бедра, прижимая их к надкрыльям. Расстояние между бугорками может быть неодинаковым: у некоторых видов бугорки у основания бедра располагаются чаще, чем в его верхней части. Этим достигается разная высота издаваемого звука. В ряде случаев, как это имеет место у трескучей огневки (Psophus stridulus), на бедре находится гладкий кант, а бугорки располагаются на передней жилке крыла. Этот вид и некоторые другие могут издавать на лету звуки иного характера. При полете они громко трещат, воспроизводя звук, напоминающий треск деревянной трещотки. Механизм этого звучания не совсем ясен, однако, очевидно, он связан с особенностями жилкования крыльев: обычно у "трещоток" сильно уплощены несколько продольных жилок крыльев.
Известно, что у одного и того же саранчового может быть несколько разных "песен". У короткокрылого конька (Chorthippus longicornis) обыкновенная "песня" состоит из 4-14 одинаковых слогов, разделенных паузами, и звучит примерно так: црэ-црэ-црэ-црэ-црэ. Ритм песни может несколько меняться в зависимости от прогрева воздуха: чем выше температура, тем больше частота ритма. Однако, помимо основной песни, у короткокрылого конька имеется еще три звуковых варианта, отличающихся частотой ритма, количеством и звучанием слогов. Можно различить "песню соперника", "призывную" и "копуляционную" песни. Первая из них возникает, когда самец слышит песню другого самца - "соперника", вторая - при встрече с самкой, а третья - при спаривании.
Тимпанальные органы помещаются у саранчовых по бокам первого брюшного сегмента, позади находящегося на нем дыхальца (рис. 158). Они представляют собой округлые или щелевидные отверстия, затянутые прозрачной барабанной перепонкой. Барабанная перепонка обрамлена хитиновым утолщением, слегка налегающим на нее спереди и сзади. Изнутри к перепонке прилегает большой воздушный мешок трахейной системы, который поддерживает ее в натянутом состоянии. По-видимому, в натяжении перепонки принимают участие также две специальные мышцы.
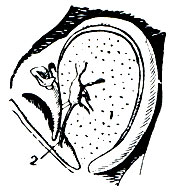
Рис. 158. Тимпанальный орган саранчового: 1 - перепонка; 2 - нерв
У саранчовых форма тела и окраска его покровов находится в очень тесном соответствии с поведением насекомого и отражает особенности той среды, в которой формировался и обитает в настоящее время тот или иной вид. Такое сочетание морфологических, биологических и физиологических свойств вида, находящееся в соответствии с окружающей средой, называют жизненной формой (табл. 30). Жизненные формы саранчовых могут быть объединены в два класса: обитатели растений, или фитофилы, и обитатели открытых участков на поверхности почвы, или геофилы.
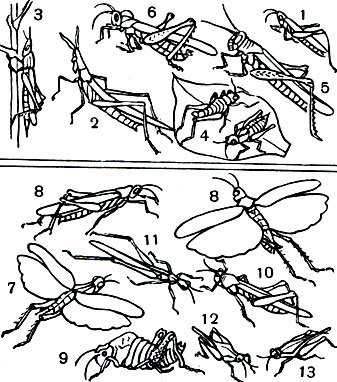
Таблица 30. Жизненные формы саранчовых: 1-3 настоящие хортобионты: 1 - степной конек (Euchorthippus pulvinatus), 2 - акрида (Acrida bicolor), 3 - тропидопола (Tropidopola longicornis); 4 - травоядный хортобионт конофима Дирша (Conophyma dirschi); 5, 6 - тамнобионты: 5 - египетская кобылка (Anacridium aegyptium), 6 - большая саксауловая горбатка (Dericorys albidula); 7, 8 - открытые геофилы: 7 - голубокрылая кобылка (Oedipoda coerulescens), 8 - пустынница (Sphingonotus octofasciatus); 9 - петробионт саксетания (Saxetania cultricollis); 10, 11- псаммобионты: 10 - обыкновенная песчанка (Hyalorrhipis clausi), 11 - стройный тонкошпор (Leptopternis gracilis); 12, 13 - гepпетобионты: 12 - Paratettix bolivari, 13 - Tetrix depressa
Среди фитофилов различают хортобионтов и тамнобионтов. Настоящие хортобионты (табл. 30, 1-3) обитают в толще травянистого покрова и питаются преимущественно злаками. У них стройное удлиненное тело, нередко со скошенным лбом; соотношение между шириной и высотой тела в наиболее широкой его части (индекс ш/в) всегда ниже единицы. В некоторых случаях тело может быть настолько сильно удлинено и к тому же лишено крыльев, что приобретает палочковидную форму, как это имеет место у южноамериканской палочковидной кобылки (Cephalocoema lineata). Покровы тела хортобионтов гладкие, лишены грубой скульптуры и окрашены под цвет зеленой или сухой травы, часто с продольными полосками вдоль тела. Крылья обычно бесцветные, реже окрашены в бледные розоватые или зеленоватые тона. Хортобионты широко распространены на лугах, в степях и саваннах, т. е. там, где преобладают травянистые растения. Типичным представителем хортобионтов может служить степной конек (Euchorthippus pulvinatus), широко распространенный в ковыльных степях Европы и Азии. Среди хортобионтов многие виды являются вредителями сельского хозяйства.
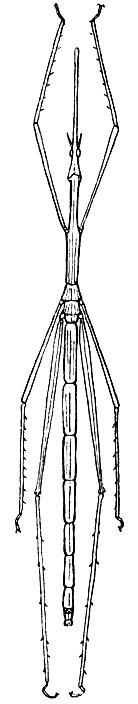
Рис. 159. Палочковидная кобылка (Cephalocoema lineata)
Некоторые хортобионты предпочитают питаться не злаками, а широколиственными травянистыми растениями; их так и называют травоядными хортобионтами. Сюда относятся виды бескрылых кобылок конофим (Соnophyma, табл. 30, 4), широко распространенных в горах Средней Азии, и целый ряд других форм. У травоядных хортобионтов туловище более коренастое, а голова с отвесным лбом. Характерно, что у них верхняя челюсть приспособлена к перетиранию и пережевыванию мягких листьев.
В ряде случаев встречаются виды, которые хотя и обитают в толще травы, но не избегают и открытых пространств на поверхности почвы, как это делают некоторые крестовички (Dociostaurus). Такие саранчовые называются факультативными хортобионтами. По своему поведению и форме тела они являются переходными к геофилам.
В отличие от хортобионтов, связанных с травянистой растительностью, тамнобионты (табл. 30, 5, 6) живут на кустарниках и деревьях. Приспособления к такому образу жизни прежде всего выражаются в специфичной структуре конечностей: у них наблюдается асимметрия в вооружении голеней задних ног, на внутренней стороне которых шипы длиннее шипов наружного ряда; асимметричными могут быть и находящиеся на лапках коготки, между которыми находится сильно развитая присоска. Некоторые тамнобионты тесно связаны с определенными видами растений. Например, большая саксауловая горбатка (Dericorys albidula), как и некоторые другие виды этого рода, связана с саксаулом. В песчаных пустынях Средней Азии в годы массового размножения этого насекомого оно может сильно повреждать саксаул.

Таблица 30. Жизненные формы саранчовых: 1 - 3 настоящие хортобионты: 1 - степной конек (Euchorthippus pulvinatus), 2 - акрида (Acrida bicolor), 3 - тропидопола (Tropidopola longicornis); 4 - травоядный хортобионт конофима Дирша (Conophyma dirschi); 5, 6 - тамнобионты: 5 - египетская кобылка (Anacridium aegyptium), 6 - большая саксауловая горбатка (Dericorys albidula)
Второй класс жизненных форм - геофилы - заключает в себе саранчовых, обитающих обычно на поверхности почвы, забирающихся на растения только во время питания. В нем различают открытых геофилов и скрытоживущих геофилов (герпетобионтов). Открытые геофилы (табл. 30, 7, 8), в отличие от хортобионтов, имеют более уплощенное тело с индексом ш/в, поднимающимся до единицы и выше; у них всегда слабо развиты, а иногда и совсем отсутствуют присоски между коготками. Покровы тела у открытых геофилов плотные, морщинистые, часто покрыты бугорками. Окраска их тела и надкрылий всегда защитная и соответствует тому общему фону, на котором они встречаются. Задние крылья нередко ярко окрашены, с темными, бросающимися в глаза перевязями. Такое сочетание окрасок надкрылий и крыльев стоит в тесной связи с поведением открытых геофилов. Можно подойти совсем близко к сидящему насекомому и не заметить его, так хорошо оно замаскировано. И вдруг совершенно неожиданно оно вылетает прямо из-под ног, привлекая к себе внимание яркими флагами раскрытых крыльев. Быстро перелетев на некоторое расстояние, насекомое так же внезапно садится на землю и вновь исчезает из глаз. Некоторые геофильные виды способны во время полета издавать трескучие звуки; так делают различные трещотки (Bryodema, Angara cris), трескучая огневка, некоторые пустынницы (Sphingonotus) и др. Летящие ярко окрашенные геофильные саранчовые привлекают к себе внимание движением, окраской и звуком, но, как только садятся, они исчезают из глаз преследователя, явно дезориентируя его.
Высокая криптичность окраски наружных покровов легко объясняется явлением гомохромии, выражающимся в том, что при развитии личинок одного и того же вида на различных фонах их окраска будет сильно варьировать и в конечном счете в каждом конкретном случае в той или иной мере соответствовать фону. Это доказано специальными экспериментами, проводившимися с разными саранчовыми, и в том числе с голубокрылой кобылкой (Oedipoda coerulescens). Воспитанные в условиях опыта экземпляры этого вида имитировали окраску почвы, на которой они выросли, приобретая соответственно черный, красноватый или серый оттенок.
Открытые геофилы преимущественно обитатели пустынь и полупустынь; представители этой группы, встречающиеся в степях или в лесной зоне, предпочитают открытые, хорошо прогреваемые места с разреженной растительностью на обнажениях горных пород, солончаках, галечниках по берегам рек и т. д.
Среди открытых геофилов особо отличают петробионтов ипсаммобионтов. Петробионты, или камнелюбы, обитают на каменистых участках, чаще всего на склонах гор. У них более резко, чем у остальных геофилов, выражена плотность и скульптурность покровов. Тело очень широкое и укороченное, вальковатое; переднеспинка обычно высокая, в ряде случаев с сильно приподнятым килем, как это имеет место, например, у саксетаний (Saxetania, табл. 30, 9), широко распространенных в северных горах Ирана и Афганистана, а у нас в Копет-Даге и некоторых других горных системах Средней Азии. У петробионтов обычно редуцирован летательный аппарат, а движения очень замедленны. Зато криптическая окраска выражена очень хорошо и всегда отражает специфику фона каменистых склонов.
Иные приспособления у псаммобионтов (табл. 30, 10, 11) - обитателей песчаных массивов, особенно ярко представленных в песчаных пустынях. К ним в первую очередь нужно отнести виды песчанок (Hyalorrhipis) и тонкошпоров (Leptopternis). Их окраска хорошо имитирует общий тон и специфичную зернистость песчаного субстрата. Своеобразны их приспособления к своему местообитанию: у псаммобионтов сильно удлинены шпоры на голенях задних ног, что облегчает отталкивание на сыпучем песке припрыжке.
В отличие от открытых геофилов герпетобионты (табл. 30, 12, 13) представляют собой формы, обитающие на поверхности почвы, занятой разреженным травянистым покровом, опадом листвы или другими растительными остатками. Их можно также встретить на увлажненном песке по берегам водоемов. Они очень требовательны к повышенному увлажнению. Обычно это мелкие формы с веретеновидным телом, суженным от заднегруди вперед и назад, окрашенным в защитные, часто сероватые тона. Типичными представителями герпетобионтов являются виды специфичного семейства прыгунчиков, или тетригид (Tetrigidae).

Таблица 30. Жизненные формы саранчовых: 7, 8 - открытые геофилы: 7 - голубокрылая кобылка (Oedipoda coerulescens), 8 - пустынница (Sphingonotus octofasciatus); 9 - петробионт саксетания (Saxetania cultricollis); 10, 11- псаммобионты: 10 - обыкновенная песчанка (Hyalorrhipis clausi), 11 - стройный тонкошпор (Leptopternis gracilis); 12, 13 - гepпетобионты: 12 - Paratettix bolivari, 13 - Tetrix depressa
Все распространенные в СССР саранчовые имеют в году одну генерацию, причем большинство из них зимует в фазе яйца. Однако существуют виды, дающие в течение года 2-3 поколения. Для саранчовых, так же как и для других прямокрылых, характерно сперматофорное оплодотворение; сперматофоры имеют вид или пузыревидных резервуаров с длинной выводной трубчатой частью, или округлых баллонов. При спаривании, которое продолжается в течение нескольких (до 20) часов, самец вводит сперматофоры со спермиями в половые пути самки.
Преобладающее большинство видов откладывает яйца в поверхностный слой почвы, прокапывая в нем с помощью короткого, состоящего из четырех створок яйцеклада ход. При этом самка глубоко погружает сильно растягивающееся брюшко в землю и выпускает порцию яиц, взвешенных в особой пенистой жидкости. Эти пенистые выделения, застывая, нередко цементируют частицы земли, окружающей кладку, вследствие чего формируется так называемая кубышка, представляющая собой капсулу, часто с твердыми землистыми стенками, внутри которой помещаются яйца (рис. 160). В некоторых случаях кубышки могут быть прикрыты особой крышечкой. Вообще структура кубышек, количество находящихся в них яиц, а также число откладываемых самкой кубышек различно у разных видов. Например, у многих видов травянок (Stenobothrus) стенки кубышки инкрустируются собственными экскрементами. Только некоторые хортобионты откладывают яйца на растения или внутрь их органов. Короткокрылый зеленчук (Euthystira brachyptera), например, помещает кубышку между листьями около корней растения или в опавшую листву; непарный зеленчук (Chrysochraon dispar) откладывает яйца в стебли малины и других растений.
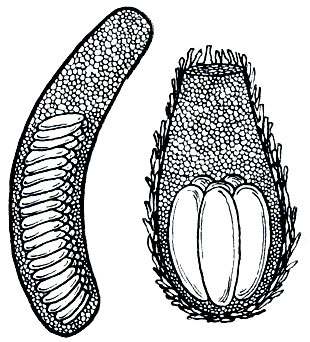
Рис. 160. Кубышки с яйцами: слева - перелетной саранчи (Locusta migratoria); справа - толстоголовой травянки (Stenobothrus lineatus)
Развитие зародыша начинается сразу после откладки яиц, но затем у большинства наших видов прекращается еще до наступления холодов и возобновляется весной после перезимовки; таким образом, имеется эмбриональная диапауза. Отрождение саранчовых начинается весной после стаивания снега и достаточного прогревания почвы. Выходящая из яйца личинка имеет червеобразную форму. У нее есть особый кратковременно функционирующий орган - пульсирующий пузырь, с помощью которого она раздвигает частицы почвы и, совершая червеобразные движения, выходит на ее поверхность, где сразу линяет и превращается в личинку первого возраста. Отродившиеся личинки сначала имеют молочно-белую окраску тела, а затем, часа через 2-3, темнеют. Теперь они уже похожи на взрослое саранчовое, отличаясь от него меньшими размерами, отсутствием крыльев и уменьшенным числом члеников усиков (не более 13). Иногда у них намечаются крыловые зачатки в виде слегка оттянутых нижних задних углов заднеспинки.
Личиночное развитие в зависимости от вида саранчового и условий среды длится 30-40 дней. За это время личинки линяют 4-5 раз, причем с каждой линькой у них увеличиваются размеры тела и крыловых зачатков, а также нарастает количество члеников усиков (рис. 161). У личинок второго возраста крыловые зачатки уже ясно заметны в нижних задних углах средне- и заднеспинки, но на них еще отсутствуют жилки; число члеников усиков увеличивается до 15-17, а иногда до 18-19. У личинок третьего возраста крыловые зачатки оттягиваются еще сильнее и на них ясно различаются жилки. Для личинок четвертого возраста характерно расположение крыловых зачатков на спине в виде треугольных, не превышающих по длине переднеспинку лопастинок, из которых внутренняя пара (зачатки надкрылий) короче и уже крыльев. Число члеников усиков продолжает нарастать, доходя в ряде случаев до 22. У личинки последнего (пятого) возраста обе пары зачатков достигают одинаковой длины, которая или равна длине переднеспинки, или уже превышает ее.
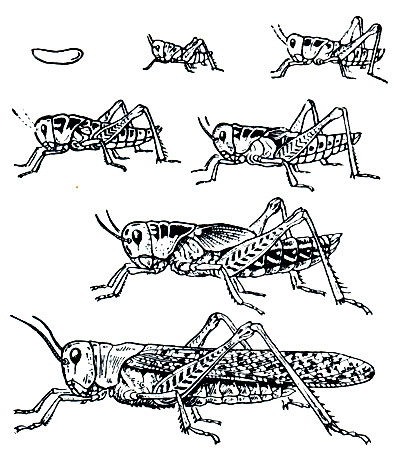
Рис. 161. Развитие перелетной саранчи
Среди саранчовых многие виды являются опасными вредителями сельского хозяйства, вследствие чего человек давно обратил на них внимание. На фресках и папирусах, написанных в древнем Египте еще за 3000 лет до нашей эры, уже изображалась саранча, а описания причиняемых ею бедствий в Египте, Ливии и Палестине датируются 1490-904 годами до нашей эры.
Саранчовый вопрос не потерял своей актуальности и в настоящее время, так как на всех континентах эти насекомые ежегодно наносят огромный ущерб сельскому хозяйству. Достаточно указать, что в 23 западных штатах США в период с 1925 по 1949 год ежегодные потери от саранчовых превышали 31 миллион долларов. Высокая вредоносность этих насекомых объясняется прежде всего тем, что некоторые виды саранчовых способны периодически давать вспышки массового размножения.
В 1928 году в Лондоне был создан специальный Противосаранчовый центр, который в настоящее время стал научной организацией международного значения. Здесь изучаются закономерности массового размножения важнейших видов вредных саранчовых и разрабатываются новые методы борьбы с ними. С 1961 года по соглашению с Продовольственной и сельскохозяйственной организацией Объединенных наций (ФАО ООН) в Противосаранчовом центре организована особая информационная служба по пустынной саранче. В ее обязанности входит научная обработка различных информации об этом вредителе, поступающих из всех стран мира.
Давно уже принято вредных саранчовых делить на стадных, иначе называемых саранчой, и нестадных, которых именуют кобылками. Стадные виды (перелетная саранча, мароккская саранча, пустынная саранча, или схистоцерка, красная саранча и др.) при массовом размножении живут скрыто, группируясь в кулиги (скопления личинок) или стаи (скопления взрослых).
Всем саранчовым в той или иной мере свойственны различного типа миграции, выражающиеся в виде переходов или перелетов. Хортобионты регулярно совершают вертикальные миграции, передвигаясь по растениям вверх и вниз в утренние и вечерние, а нередко и в дневные часы суток. Геофилы все время совершают днем горизонтальные миграции, перелетая с одного места на другое. Мигрируют саранчовые и с участков, где они кормятся, в места, удобные для откладки яиц. Однако миграционные инстинкты особенно развиты у стадных видов, у которых активные переходы и перелеты осуществляются кулигами и стаями на большие расстояния. Как личинки, так и взрослые саранчовые ночуют обычно на верхних частях растений, где они пребывают в ночное время в состоянии холодового оцепенения. После восхода солнца по мере прогрева воздуха личинки становятся активными и концентрируются на освещенных участках почвы или иногда на растениях, образуя так называемые "солнечные кулижки". Это плотные скопления неподвижно сидящих личинок, в которых каждая особь тесно прижимается к соседним. Первичные "солнечные кулижки" образуются личинками, отрождающимися из одной кубышки. Как только температура тела находящихся совместно особей поднимается выше 40°С, кулижки рассеиваются и начинается пешая миграция, которая прекращается вечером, когда личинки опять впадают в состояние покоя. В жаркое время дня, в случае если температура тела насекомого поднимается выше 50°С, наступает дневной период тепловой депрессии, в котором также наблюдается состояние относительного покоя.
В годы массового размножения образуются настолько крупные кулиги, что занимаемая ими площадь может достигать размеров сотен и тысяч гектаров. В период личиночного развития кулиги все время находятся в состоянии миграционной активности, передвигаясь с мест отрождения в сторону участков с более благоприятными условиями температуры, влажности и питания. В результате за период личиночной жизни стадные саранчовые могут проходить очень большие расстояния - до 30 км и больше, как это имеет место, например, у азиатской саранчи. Характерно, что движение кулиги имеет определенное направление и осуществляется обычно в сторону, противоположную направлению ветра. Оптимальные условия для движения создаются при небольшом ветре (3-5 м/сек); при порывистом или сильном ветре, превышающем 10 м/сек, движение кулиг прекращается.
После окрыления саранчи ее передвижения принимают вид миграционных полетов, когда стаи перелетают за день расстояние в несколько десятков километров. Характерно, что направление движения стаи стоит в очень тесной связи с направлением ветра. Так, при слабом ветре (до 1 м/сек) стаи азиатской саранчи летят обычно по ветру; при повышении силы ветра до 5 м/сек саранча взлетает с большим трудом, а сам полет осуществляется под острым углом к направлению ветра. Полеты могут осуществляться не только днем, но в некоторых случаях и ночью, однако скорость полета при этом резко снижается: днем азиатская саранча пролетает 30-40 км, а ночью не больше 15 км, За весь период движения стая может улететь от своего гнездилища за 200-300 и больше километров. Еще дальше залетает пустынная саранча: в 1954 году небольшие стаи этого вида из Северо-Западной Африки достигли Британских островов, пролетев более 2400 км над открытым морем. Значительно меньшая миграционная способность у итальянского пруса и мароккскои саранчи, которые отлетают от мест отрождения на более короткие расстояния, измеряемые десятками километров.
Как было установлено Б. П. Уваровым, у стадных саранчовых наблюдается весьма своеобразное явление так называемой фазовой изменчивости, сущность которой заключается в изменении внешнего облика и физиологических функций насекомого в зависимости от степени скученности и других условий в процессе развития. При высокой скученности развивающихся особей данного вида возникает стадная фаза, а при развитии в разреженной популяции - одиночная. Эти фазы прежде всего хорошо отличаются внешними признаками. Так, у особей перелетной саранчи стадной фазы срединный киль переднеспинки в профиль прямой или даже слегка вогнутый, тогда как у одиночной фазы того же вида он дугообразно выгнут. Личинки стадной фазы имеют пеструю окраску тела, в которой сочетаются черный и оранжевый цвета; у личинок одиночной фазы окраска тела гармонирует с окружающим фоном и чаще бывает одноцветной зеленой (табл. 31, 2, 3).
Считают, что более обычным является состояние одиночной фазы, при котором особи живут на местности рассеянно, и плотность популяции в этом случае очень низка. Переход в стадную фазу обусловливается резким увеличением численности популяции, приводящим к совместному существованию личинок в виде больших кулиг. Такие концентрации у одиночной фазы наблюдаются прежде всего в случае изменения условий существования, например при недостатке влаги, приводящем к высыханию растительности. При этом происходит миграция в места с более благоприятными условиями, где еще сохраняется зеленая растительность.
Такая вынужденная концентрация приводит к очень тесному контакту особей, формирующих кулигу. Этот контакт осуществляется не только через органы зрения и обоняния, но и, что особенно важно, через органы осязания. Личинки обмениваются прикосновениями антенн и толчками задних ног, что в конечном счете приводит к возникновению особого условного рефлекса - рефлекса стадности. Стадные привычки усиливаются с возрастом, и личинки обнаруживают тенденцию сохранять их, даже будучи изолированными от кулиги.
Стадный образ жизни повышает возбудимость нервной системы и изменяет направленность обмена веществ в организме. При большом скоплении саранчи самки возбуждаются другими особями не только механически, вследствие непосредственного соприкосновения, но и химически особым гормоном - феромоном, выделяемым клетками кожных покровов по всей поверхности тела самца. Тот же феромон у пустынной саранчи обусловливает характерную желтую окраску зрелых самцов. Интересно, что подобную окраску можно получить искусственно увеличением содержания в воздухе СО2 до 3-4% (по объему).
Характерные изменения происходят и в деятельности желез внутренней секреции, в том числе в так называемых прилежащих телах - особых парных железах, располагающихся в голове позади мозга и топографически связанных с аортой. Опыты с перелетной саранчой показали, что если личинкам четвертого или пятого возраста с ясно выраженными признаками стадной фазы пересадить прилежащие тела одиночной саранчи, то у них после линьки появляется характерная для одиночной фазы зеленая окраска. Однако при этом сохраняется повышенная интенсивность дыхания и подвижность, типичные для личинок и имаго стадной фазы. Следует заметить, что накопление темных пигментов в наружных покровах у особей, ведущих стадный образ жизни, способствует поглощению солнечной энергии и повышению температуры тела.
Скученность особей в кулигах и стаях влияет и на плодовитость саранчовых, понижая ее. В лабораторных условиях у потомства особей перелетной саранчи, воспитываемых в изоляции, плодовитость самок достигает 1000-1200 яиц, в то время как у потомства стадной фазы она составляет всего 300 яиц. В то же время потомство стадной фазы перелетной саранчи, схистоцерки и красной саранчи отличается более крупными размерами и большей способностью выживать при голодании. Это имеет огромное приспособительное значение, так как в районах перенаселения личинки попадают в более тяжелые условия жизни.
Таким образом, стадность саранчовых нужно рассматривать как комплекс полезных приспособлений, обеспечивающих существование вида в своеобразных условиях засушливого климата, где насекомые, не имеющие этих приспособлений, не могут сохранить большого потомства.
Виды саранчовых, имеющие широкий ареал, охватывающий различные ландшафтные зоны, в разных частях области своего распространения заселяют разные места обитания (стации). Выявлена следующая закономерность: такие виды в северной части своего ареала населяют более сухие (ксерофитные) стации, по мере продвижения на юг они последовательно перемещаются в более влажные места обитания - сначала в мезофитные (со средним увлажнением), а затем в гигрофитные (сильно увлажненные). Так, например, перелетная саранча в центрально-черноземной области и на юге Западной Сибири обитает на песчаных участках, а в пустынях Средней Азии и Казахстана - на побережьях рек, озер и морей в зарослях тростника. Итальянский прус на юге лесной зоны поселяется на меловых обнажениях с разреженным растительным покровом, а в Средней Азии - в долинах рек, оазисах и предгорьях, почему его и называют там оазисным прусом. Это явление получило название принципа зональной смены стаций (принцип Г. Я. Бей-Биенко). Оно объясняется в основном спецификой температурного режима и режима влажности в различных зонах.
Преобладающее большинство саранчовых - обитатели жарких стран: свыше 50% известных видов населяют тропики и субтропики. В СССР известно до 485 видов, относящихся к 5 семействам.
Тетригиды, или прыгунчики (семейство Tetrigidae),- одни из самых мелких саранчовых с темным телом, окрашенным под цвет земли, и характерной переднеспинкой, вытянутой сзади в длинный отросток, прикрывающий сверху брюшко. Надкрылья у них если и есть, то очень короткие, в виде небольших лопастей. Крылья же развиты нормально. Тетригиды не стрекочут и лишены органов слуха. В тропиках тетригиды представлены большим разнообразием видов, тогда как у нас их меньше 30. Биология этих саранчовых довольно своеобразна. Это все герпетобионты, обитающие на сырых лугах, опушках лесов, по берегам водоемов. Постоянно прыгая около воды, они могут попадать в нее, причем некоторые виды в этих случаях способны плавать и даже нырять.
Являясь выходцами из тропиков, тетригиды отличаются своеобразным ритмом развития. В наших условиях они зимуют в виде личинок или взрослых насекомых. У некоторых видов может быть 2-3 поколения в году.
Эумастациды (семейство Eumastacidae) характерны для лесных областей тропического пояса. У нас они встречаются только в Средней Азии, где представлены 16 видами, относящимися к 3 родам (Clinomastax, Phytomastax, Gomphomastax). Это небольшие или средней величины бескрылые саранчовые, длиной 1-2,5 см, с короткой головой и сжатым с боков телом; не стрекочут и лишены слухового аппарата. Обитают в условиях горного ландшафта и могут проникать высоко в горы - до 3000-4000 м. Преимущественно тамнобионты, предпочитающие держаться на кустарниках (карагана, шиповник, можжевельник) или кустарничках. Примером может служить Phytomastax robusta, живущий в горах Казахстана (Заилийский Алатау) и Киргизии (Кунгей-Алатау) на больших высотах, от 1800 м и выше. На щебнистых склонах держится на полыни и некоторых кустарничках.
Представители оригинального семейства памфагид (Pamphagidae) распространены по всей Африке, а также в пустынных и горных районах Южной Европы, Передней, Средней и Центральной Азии, достигая Дальнего Востока. В пределах СССР обитает около 50 видов.
Памфагиды - крупные саранчовые с очень плотными шероховатыми наружными покровами. Для них характерна специфичная структура бедер задних ног, наружная поверхность которых не имеет перистых площадок, расположенных между килями. Звуковой аппарат обычно имеется даже у бескрылых форм.
По образу жизни все встречающиеся у нас памфагиды или типичные геофилы, или специализированные обитатели каменистого субстрата - петробионты. Первые из них распространены в основном на широких равнинах пустынь и полупустынь. Они очень подвижны и имеют хорошо развитые крылья. В Средней Азии часто можно встретить тринхусов (Thrinchus) - характерных эндемиков среднеазиатских пустынь. Они обитают на глинисто-галечниковых и каменистых участках в долинах и в предгорьях. Тринхусы отличаются мощным, коренастым темноокрашенным телом и зеленоватыми или голубоватыми крыльями с темной перевязью посредине. Они очень подвижны: если спугнуть сидящее на земле насекомое, оно быстро перелетает на другое место, делая бросок на 5-10 м.
Обитающий в Кызылкуме пустынный тринхус (Thrinchus desertus) приспособился к условиям песчаной пустыни и имеет ряд черт, характерных для псаммобионтов. У него более стройное тело с темными продольными полосами по белому фону и удлиненные шпоры, помогающие отталкиваться от сыпучего субстрата при прыжке.
Петробионтные виды памфагид живут на каменистых склонах в горах Кавказа и Средней Азии. Большинство из них совсем лишено крыльев и имеет неуклюжее, вальковатое тело. Типичным петробионтом является саксетания (Saxetania cultricollis), распространенная в Иране (Северный и Центральный Хорасан), а также в некоторых горных хребтах Средней Азии (Копет-Даг, Кугитанг, Гиссарский хребет). Это бескрылый вид крупных размеров, длиной 2,5-6 см, с резко дуговидным срединным килем переднеспинки, Окраска тела, то серая, то темноватая, хорошо гармонирует с общим фоном каменистого субстрата; яркая, интенсивно синяя окраска выступает только на внутренней стороне голеней задних ног. По-видимому, это своеобразная "опознавательная метка", облегчающая встречу самца с самкой. Саксетания обитает на каменистых склонах гор с разреженной растительностью. Она очень медлительна, прыжки делает небольшие, и поэтому не способна к передвижениям на большие расстояния.
Пиргоморфиды (семейство Pyrgomorphidae) широко представлены в тропиках, тогда как в пределах СССР встречается всего 3 вида. От памфагид они легко отличаются конической головой с сильно скошенным лбом. Из наших представителей широко распространена пустынная пиргоморфа (Pyrgomorpha conica), обитающая в пустынях и полупустынях Казахстана, Средней Азии и Закавказья. Она относительно небольших размеров (1,5-2 см), с удлиненным телом, хорошо развитыми надкрыльями и крыльями и весьма характерной головой с мечевидными усиками и теменем, выступающим вперед наподобие утиного носа. Встречается в виде двух цветовых форм - серовато-желтой и зеленой. Держится в травянистом покрове, часто выбирая место вблизи рек и других водоемов, а в горах - днища и склоны ущелий, богатые ручьями и родниками.
Настоящих саранчовых (семейство Acrididae) легко отличить от всех рассмотренных выше семейств. От тетригид и эумастацид они прежде всего отличаются наличием звукового аппарата, помимо отсутствия тех признаков, которые специфичны для этих семейств. В то же время у них совершенно иная, чем у памфагид и пиргоморфид, скульптура наружной поверхности бедер задних ног - в виде правильно расположенных между килями перистых площадок. Это семейство богато видами: в фауне СССР на его долю приходится около 80% известных в настоящее время видов.
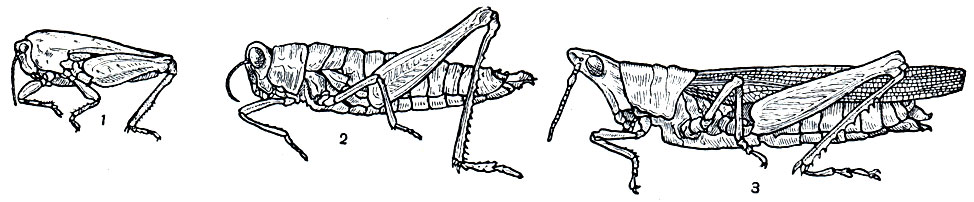
Рис. 162. Саранчовые; 1 - Tetrix tenuicornis; 2 - Phytomastax robusta; 3 - Pyrgomorpha conica
Форма тела у настоящих саранчовых сильно варьирует в зависимости от принадлежности вида к той или иной жизненной форме. По существу, в этом семействе выражены почти все жизненные формы, исключая только герпетобионтов и петробионтов.
Среди распространенных в СССР видов свыше 100 в той или иной мере могут вредить сельскохозяйственным культурам. Из них наиболее опасны стадные формы - перелетная саранча, мароккская саранча, прус итальянский и туранский, а также залетающая в некоторые годы из Ирана и Афганистана пустынная саранча (табл. 31, 1-6).
Перелетная саранча (Locusta migratoria) - крупное насекомое, до 6 см длиной; сероватая или оливково-бурая, в мелких пятнышках; переднеспинка с резким срединным килем; бедра задних ног изнутри в основной части синевато-черные; голени задних ног желтоватые или красные. О том, чем отличаются друг от друга стадная и одиночная фазы, уже говорилось выше. В СССР перелетная саранча встречается на юге Украины, в Предкавказье, в низовьях Волги, в южных районах Западной Сибири, в Казахстане и Средней Азии. На юг за пределами нашей страны она распространена до южной Африки, Австралии и Новой Зеландии.
На этой огромной территории саранча распадается на 7 подвидов, из которых в СССР встречаются два - среднерусская и азиатская перелетная саранча. Среднерусская саранча обитает в лесной и лесостепной зонах Европейской части СССР; в массе размножается в жаркие и сухие годы. Наибольшую опасность представляет азиатская саранча, постоянные гнездилища которой находятся в плавнях низовьев таких крупных рек, как Волга, Урал, Дон, Терек, Амударья, Сырдарья, а также по берегам крупных озер - Балхаша, Алаколя и Зайсана (рис. 163). Здесь она предпочитает заболоченные участки с густыми зарослями тростника. В этих условиях во второй половине лета азиатская саранча откладывает яйца на окраинах тростниковых массивов. Весной личинки отрождаются довольно поздно - в первой половине мая; развиваются они в течение 35-40 дней, после чего происходит окрыление. Скорость движения кулиги зависит от возраста личинок: у личинок третьего и четвертого возрастов она достигает 6-7 м в минуту, у пятого - до 10-12 м. Во время передвижения кулигам иногда приходится переправляться через водные пространства - болота, протоки, рукава рек. Известны случаи, когда личинки саранчи переправлялись через такие большие реки, как Кума и Сырдарья.
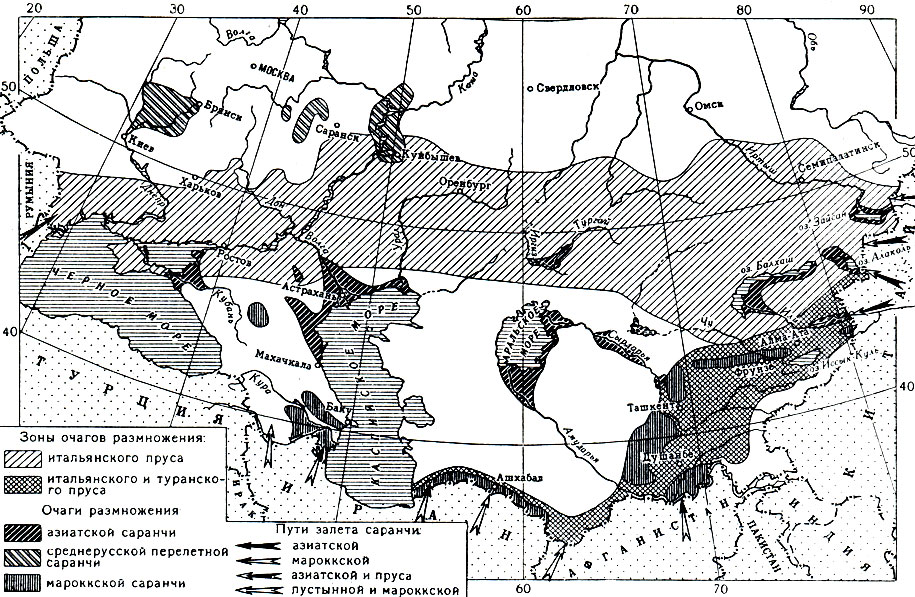
Рис. 163. Схематическая карта основных очагов массового размножения стадных саранчовых на территории СССР и путей залета саранчи в СССР из сопредельных стран
Стая взрослой саранчи летит со скоростью 10-15 км в час и за день пролетает 80-120 км. Поэтому, несмотря на ограниченность размеров постоянных гнездилищ, азиатская саранча может распространяться на большие расстояния и в течение нескольких лет подряд жить в новых заселенных ею местах, проходя здесь свойственное ей развитие. Из всех наших саранчовых азиатская саранча самая прожорливая. Подсчитано, что каждый экземпляр ее в течение своей жизни съедает 300 г зеленого корма. Потомство одной самки саранчи за лето уничтожает столько корма, сколько хватило бы для двух овец. Кулиги и стаи этого насекомого в течение 1 - 2 часов могут уничтожить сотни и тысячи гектаров посевов. Излюбленной пищей азиатской саранчи в местах ее отрождения является тростник. Во время движения она вредит многим растениям, предпочитая культурные и дикорастущие злаки. Чем старше саранча, тем она становится более многоядной и, помимо злаков, может наносить большой вред капусте, арбузам, подсолнечнику и другим растениям.
Мароккская саранча (Dociostaurus maroccanus) распространена в Средней Азии, Казахстане, на Кавказе, в Закавказье, Крыму, а за пределами СССР - в странах, окружающих Средиземное море, в Иране и Афганистане. Она значительно меньше размером, чем перелетная саранча, достигает в длину 22-38 мм; рыжевато-желтая, с темными пятнами, светлым крестообразным рисунком на переднеспинке, розоватыми или желтоватыми бедрами и красными голенями задних ног.
Места откладки яиц мароккской саранчи локализуются преимущественно в зоне пустынь - в предгорьях и нижней части склонов гор, не выше 1800 м. Это обычно сухие участки с суглинистыми и супесчаными сероземами, растительность которых составляют в основном эфемерные растения - живородящий мятлик, пустынные осоки и др. В таких условиях количество отложенных кубышек может доходить до 1500, а то и до 6000 штук на 1 м2. В Средней Азии личинки отрождаются в конце марта - начале апреля, а весь срок личиночного развития не превышает 25- 35 дней. Уже через 3-10 дней после окрыления начинается спаривание и откладка яиц. В это время мароккская саранча в течение суток совершает миграции, перелетая с мест откладки яиц на пониженные, хорошо увлажненные участки с обильной растительностью, где она питается. Мароккская саранча повреждает многие растения, но особенно сильно вредит хлопчатнику, хлебным злакам, люцерне, огородным и бахчевым культурам. Установлено, что основным условием, благоприятствующим ее массовому размножению, является выпадение весенних осадков в сумме около 100 мм в период развития перезимовавших яиц и отрождения личинок.
Прусами называют несколько видов саранчовых, относящихся к роду Calliptamus. Это средних размеров насекомые, длиной 14,5-48 мм, с коренастым телом, окрашенным в буровато-рыжие тона; надкрылья серые с темными пятнышками, крылья у большинства видов розовые. Наиболее опасны для сельского хозяйства итальянский прус (C. italicus) и туранский прус (C. turanicus). Они легко отличаются окраской задних ног: у итальянского пруса бедра с внутренней стороны розовые с 2 неполными черными перевязями, голени розовые или красные; бедра туранского пруса с внутренней стороны одноцветные, желтые или серые, без темных перевязей, а голени снаружи желтоватые, а изнутри бледно-оранжевые (табл. 31, 14,15).
Таблица 31. Вредные саранчовые: 1- перелетная саранча (Locusta migratoria); 2, 3 - ее личинки (2 - одиночная, 3 - стадная фазы); 4 - мароккская саранча (Dociostaurus maroccanus); 5 - итальянский прус (Calliptamus italicus); 6 - пустынная саранча (Schistocerca gregaria); 7 - атбасарка (Dociostaurus kraussi); 8-10 - сибирская кобылка (Gomphocerus si-biricus): 8, 9 - самец, 10 - самка; 11 - белополосая кобылка (Chorthippus albomarginatus); 12 - туркменская кобылка (Ramburiella turcomana); 13 - чернополосая кобылка (Oedaleus decorus); 14 - нога итальянского пруса; 15 - нога туранского пруса

Таблица 31. Вредные саранчовые: 1- перелетная саранча (Locusta migratoria); 2, 3 - ее личинки (2 - одиночная, 3 - стадная фазы); 4 - мароккская саранча (Dociostaurus maroccanus); 5 - итальянский прус (Calliptamus italicus); 6 - пустынная саранча (Schistocerca gregaria); 7 - атбасарка (Dociostaurus kraussi); 8-10 - сибирская кобылка (Gomphocerus sibiricus): 8,9 - самец, 10 - самка; 11 - белополосая кобылка (Chorthippus albomarginatus); 12 - туркменская кобылка (Ramburiella turcomana); 13 - чернополосая кобылка (Oedaleus decorus); 14 - нога итальянского пруса; 15 - нога туранского пруса
Туранский прус обитает в Средней Азии и Казахстане. Итальянский прус распространен значительно шире и в СССР вредит сельскохозяйственным культурам в среднем Поволжье, на юге Европейской части и в Западной Сибири, на Кавказе, в Средней Азии и Казахстане. Таким образом, в южных районах Казахстана и Средней Азии встречаются оба эти вида, однако их поведение здесь различно. Итальянский прус наиболее опасен в зоне поливного земледелия, где в основном вредит техническим культурам, туранский прус живет преимущественно на богаре и вредит злаковым культурам. Личинки прусов отрождаются в середине мая, а развитие их длится в течение 40-45 дней.
Пустынная саранча, или схистоцерка (Schistocerca gregaria), крупнее перелетной саранчи и достигает в длину 6 см. Только что окрылившиеся особи имеют розоватую окраску, позднее они становятся желтыми; надкрылья в темных пятнышках, а крылья совершенно бесцветные. Постоянные места обитания схистоцерки находятся в субтропической и тропической зонах Африки, Аравии, Индии и Пакистана. Благодаря своей огромной ПОДВИЖНОСТИ и способности к дальним перелетам она может выживать в жарких пустынных местностях, хотя казалось бы, что условия для ее размножения здесь неблагоприятны. Дело заключается в том, что самки откладывают яйца после дождей в песок или рыхлую почву. В течение короткого инкубационного периода, длящегося 12-15 дней, отрождаются молодые личинки, развитие которых продолжается 5-6 недель. В этот период они находят в пустыне сочную растительную пищу, благодаря тому что после дождей здесь идет вегетация растений. Взрослая схистоцерка лучше приспособлена к перенесению сухости воздуха и высоких температур. Однако для полового созревания она тоже нуждается в сочной зеленой пище.
Окрылившаяся стадная фаза пустынной саранчи может расселяться на тысячи километров. Так, в октябре 1954 года ее небольшие стаи, летящие из Северо-Западной Африки, достигли Британских островов, пролетев более 2400 км над морем. Средняя высота полета не превышает 600 м, однако с самолетов в некоторых случаях ее стаи регистрировались на высоте 2000 м. Известно, что отдельные экземпляры, поднимаясь до 6000 м, долетали до ледников Кении.
Из индийских и аравийских очагов схистоцерка долетает до Ирана и Афганистана, где образует временные очаги размножения. Из этих стран пустынная саранча может залетать в Советский Союз (рис. 163). В 1929 году огромные стаи схистоцерки проникли на территорию Средней Азии, где распространились на площади свыше 1,5 миллиона гектаров и нанесли большой ущерб сельскому хозяйству. В 1928 и 1930 годах она залетала в Закавказье. В 1962 году схистоцерка на стыке границ СССР, Ирана и Афганистана вторглась на территорию Туркмении, имея общий фронт залета протяженностью около 160 км. Залет продолжался с марта по май. При этом она отложила свои кубышки на площади около 40 тысяч гектаров. Характерно, что новое поколение после окрыления улетело обратно на юг.
Помимо стадных саранчовых, существенный вред сельскому хозяйству могут наносить и нестадные, причем в различных ландшафтных зонах вредят определенные виды кобылок. Так, в лесостепных и в северной части степных районов Сибири, Приуралья и Северного Казахстана существенный вред могут наносить сибирская (Gomphocerus sibiricus) и белополосая (Chorthippus albomarginatus) кобылки; в южных степях Казахстана - атбасарка (Dociostaurus kraussi); в условиях среднеазиатской богары - туркменская (Ramburiella turcomana) и чернополосая (Oedaleus decorus) кобылки и другие виды (табл. 31, 7-13).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'