Отряд Акариформные клещи (Acariformes)
Переходя к рассмотрению клещей, или, точнее, клещеобразных арахнид, напомним, что они представляют собой три неродственных отряда, в такой же мере самостоятельных и обособленных, как и другие отряды арахнид. Два из них, акариформные клещи и паразитиформные клещи, включают огромное число чрезвычайно разнообразных форм. Третий отряд - клещи-сенокосцы, напротив, мал и однотипен.
Как мы говорили, замечательная особенность больших отрядов клещей состоит в том, что их громадное и на первый взгляд хаотичное разнообразие в действительности представляет естественные ряды форм от примитивных исходных до сильно изменившихся в разных направлениях. Поэтому сравнительное изучение современного разнообразия каждого отряда позволяет представить пути и результаты его длительной эволюции и таким образом понять строение, индивидуальное развитие и образ жизни его многочисленных и подчас совсем непохожих друг на друга представителей. Кратко и популярно изложить данные о клещах в такой сравнительной форме нелегко, но другого пути для понимания природы клещей нет. При рассмотрении каждого отряда мы сначала остановимся на наиболее примитивных его представителях, служащих прототипами отрядов, а далее попробуем проследить, как изменялись образ жизни, строение и развитие в пределах отрядов. Конечно, мы сможем коснуться лишь некоторых основных группировок клещей в такой мере, в какой это необходимо для общей картины.
Акариформные клещи - это самый большой отряд клещеобразных, насчитывающий более 6000 видов, причем эта цифра далека от действительной. Благодаря интенсивному изучению клещей, особенно в последнее время, ежегодно появляются описания сотен новых видов. Разнообразие акариформных клещей очень велико. Отряд делится на два подотряда: саркоптиформные клещи (Sarcoptiformes) и тромбидиформные клещи (Trombidiformes). К первому относятся такие обширные и разнообразные группы, как панцирные, тироглифоидные клещи и ряд паразитических - перьевые, волосяные, чесоточные и др. Второй подотряд включает очень много группировок, в том числе паутинных, водяных клещей, краснотелок и ряд других. В этом отряде немало практически важных клещей - вредителей растений и сельскохозяйственных продуктов, паразитов животных и человека. Но переносчиков инфекционных болезней здесь сравнительно немного.
Классификация отряда сложна и находится в стадии разработки. В обоих подотрядах сейчас насчитывается более 170 семейств, а общее число родов трудно указать, настолько оно различно у разных авторов. Современная классификация, несомненно, есть результат чрезмерного дробления систематических группировок, в чем отражается аналитический этап изучения клещей, на котором находится наука о них - акарология. Одно из основных систематических подразделений- семейство обычно понимается акарологами в очень суженном смысле. Есть немало "семейств" акариформных клещей всего с одним-двумя десятками видов, а семейства по нескольку сотен видов составляют скорее исключение. Если в этом отношении сравнить клещей с насекомыми, то семействам последних более соответствуют надсемейства клещей, которыми мы и будем в основном оперировать в дальнейшем.
Наиболее примитивные акариформные клещи - это низшие представители подотряда Trombidiformes - небольшое надсемейство Endeostigmata (рис. 54, табл. 8, 1). К ним близки самые примитивные представители подотряда Sarcoptiformes - клещи-палеакариды надсемейства Palaeacaroidea (рис. 57, 1). Эти два надсемейства служат прототипом отряда в целом, а их близость указывает на единство его происхождения. Эти малоизвестные формы изучались французским акарологом Гранжаном, а у нас - А. А. Захваткиным и автором этого раздела. В последние годы удалось изучить биологию и эмбриональное развитие этих клещей.
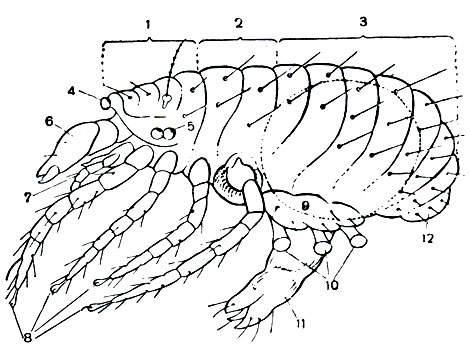
Рис. 54. Строение примитивного акариформного клеща надсемейства Endeostigmata (вид сбоку): 1 - протеросома (1-4-й сегменты); 2 - сегменты ног третьей и четвертой пар; 3 - брюшко (7-13-й сегменты); 4 - медиальный глаз; 5 - боковые глаза; 6 - хелицеры; 7 - педипальпы; 8 - ноги; 9 - половые крышки; 10 - коксальные органы; 11 - половой конус; 12 - анальные клапаны
Это очень древние клещи, найденные в ископаемом состоянии в среднедевонских отложениях и дожившие до нашего времени почти без изменений. Они живут в почвенном гумусе, и, вероятно, в этом одно из условий их эволюционного консерватизма. Почва - среда сравнительно мало менявшаяся на протяжении геологических эпох, и эти "живые ископаемые" нашли в ней такие условия, в которых смогли сохраниться.
По плану строения примитивные Acariformes соответствуют арахнидам вообще, чем опровергается ходячее мнение о том, что все клещи крайне видоизменились и деградировали. Более того, они сохранили ряд таких примитивных черт, которые давно утрачены другими арахнидами. Эти клещи очень малы, обычно всего 0,2-0,3 мм. Покровы тонкие, лишены щитков. Туловище хорошо расчленено (рис. 54), причем сегментировано не только брюшко, но и головогрудь. Сегменты хелицер, педипальп и двух передних пар ног разделены спинными бороздами и, как у кенений и сольпуг, образуют передний отдел, отвечающий голове трилобитов и называемый у клещей протеросомой. По сторонам этого отдела имеются боковые глаза, а спереди парный по происхождению медиальный глаз. Сегменты двух задних пар ног расчленены. Членистое брюшко состоит из шести сегментов и соединяется с головогрудью хорошо развитым предполовым сегментом. У других акариформных клещей сегментация обычно исчезает. Сохраняется борозда между протеросомой и задним отделом туловища, который образуется путем присоединения сегментов двух задних пар ног к брюшку и называется гистеросомой. Такой тип расчленения тела очень характерен для всего отряда.
Важную роль приобретают волоски, щетинки и другие кожные органы. Благодаря малым размерам клещей эти органы у них бывают относительно очень велики и обычно строго постоянны по числу и положению на сегментах туловища и придатках. Часть из них выполняет функции органов чувств - осязательных, воспринимающих колебания (трихоботрии на переднем отделе тела), обонятельных. Последние представлены лировидными органами и так называемыми соленидиями на ногах. Это тонкостенные замкнутые на конце трубочки со спиральной бороздой, пронизанной мельчайшими канальцами; внутрь соленидия входит отросток чувствительной нервной клетки. Щетинки нередко служат дополнительной защитой, особенно у тонкокожих форм, и бывают очень разнообразны по величине и форме - шиловидные, перистые, плоские, покрывающие клеща, как черепица, и т. п.
Хелицеры и педипальпы у низших Асаriformes свободные, не соединяются в "головку", что считалось характерным для всех клещей. Хелицеры обычно 3-члениковые, с клешней, но есть формы, у которых можно различить до шести члеников хелицер, чем последние очень похожи на ноги. Тазики педипальп подвижные и имеют жевательные лопасти. Ноги с одним когтевидным эмподием, еще лишены добавочных коготков, которые в дальнейшем образуются из щетинок.
Строение ротовых органов в деталях довольно разнообразно уже у низших форм. Как удалось выяснить, это связано с разнообразием питания. Одни из них жующие растительноядные, потребляют гифы и споры грибов, почвенные водоросли, дрожжи и т. п. Другие ловят ногохвосток, других клещей, прокалывают их хелицерами и высасывают. Третьи высасывают яйца ногохвосток и т. д.
Замечательно строение наружного полового аппарата - сложного образования, занимающего большую часть нижней поверхности брюшка и сходно устроенного у обоих полов. Как теперь выясняется, в его состав входят конечности трех передних сегментов брюшка. Половое отверстие открывается на конце крупного выпячивающегося полового конуса на восьмом сегменте. Ножки восьмого, девятого и десятого сегментов срастаются и образуют половые крышки, прикрывающие половой конус. Коксальные органы брюшных ножек, имеющие вид выпячивающихся мешочков, наполняемых гемолимфой, в числе трех пар расположены под крышками. Они называются у клещей половыми присосками. Это органы чувств, играющие определенную роль при сперматофорном оплодотворении и откладке яиц, по-видимому, воспринимающие влажность. Благодаря микроскопическим размерам низших клещей их сперматофоры и особенно яйца, будучи малы абсолютно, очень велики относительно взрослой особи, с чем и связана громоздкость и сложность наружного полового аппарата. Яйца созревают и откладываются по одному, так как в теле самки может уместиться обычно только одно готовое к откладке яйцо.
Примитивные Acariformes лишены специальных органов дыхания и дышат кожей. В сочетании с тем, что только что говорилось о судьбе брюшных ножек, это приводит нас к выводу, что предки этого отряда клещей никогда не имели легких: из воды они прямо перешли к жизни во влажной почве и кожному дыханию, в чем сходны с кенениями. В дальнейшем у акариформных клещей образовались трахеи, причем в разных группах независимо, в разных местах. Примитивные черты обнаруживаются во внутреннем строении низших форм, например, имеется три пары выделительных коксальных желез, функционирующих у взрослых клещей, сохраняется сегментация кишечника и т. п.
Индивидуальное развитие низших Acariformes представляет большой интерес. Оплодотворение сперматофорное. Самец оставляет на субстрате клейкие сперматофоры на ножке, а самка захватывает их наружными половыми органами. Но у некоторых видов даже нет еще сперматофора, а просто выделяются капли семенной жидкости, которые самка, идя за самцом, всасывает половым конусом. Яйца очень малы (0,1-0,2 мм) и бедны желтком. Благодаря этому индивидуальное развитие сохранило архаичные черты, утраченные другими арахнидами. Дробление яйца полное, равномерное, по некоторым признакам напоминает дробление яйца кольчатых червей. Зародыш проходит четырехсегментную стадию (рис. 55, А), сходную с личинкой трилобитов - протасписом (рис. 6).
Клещи этого отряда выходят из яйца на значительно более ранней стадии развития, чем все остальные арахниды. Образование кутикулы и подготовка к вылуплению начинается в то время, когда у зародыша еще не развита четвертая пара ног, нет зачатков брюшных ножек и недостает трех концевых сегментов брюшка. Поэтому вылупляется шестиногая личинка с коротким брюшком, лишенным трех последних сегментов и половых створок. Первая кутикула, одевающая зародыш, снабжена шипиками или упругими щетинками ("яйцевыми зубами"), которые служат для разрыва яйцевых оболочек. Это так называемая предличинка. У большинства акариформных клещей под этой первой кутикулой в яйце вскоре образуется кутикула собственно личинки, которая, вылупляясь, сбрасывает яйцевые оболочки и предличиночный покров. Но у низших представителей отряда предличинка вылупляется сама, хотя обычно бывает неподвижна. Недавно автору этого раздела удалось найти вид низших клещей, у которых предличинка подвижна и хорошо бегает, так что по существу сохранилось два активных личиночных возраста, разделенных линькой.
Личинка (рис. 55, В) превращается в нимфу, которая, трижды линяя, становится взрослым клещом. Соответственно различают три нимфальных возраста: протонимфу, дейтонимфу и тритонимфу. В течение нимфального периода появляются сегменты брюшка и конечности, недостававшие у личинки (рис. 55, Г-Е). Замечательно, что новые сегменты образуются один за другим вперед от анальной зоны роста и одновременно на лежащих впереди сегментах появляются конечности. Иначе говоря, здесь частично сохранился анаморфоз, т. е. тот первичный способ развития, который мы видели у предков хелицеровых - трилобитов. При превращении личинки в протонимфу появляются ноги четвертой пары, еще не вполне развитые и почти лишенные щетинок; на конце брюшка, перед створками, прикрывающими анальное отверстие, образуется новый, одиннадцатый по общему счету сегмент; на восьмом сегменте появляются видоизмененные ножки - зачатки половых клапанов с парой коксальных органов под ними. У дейто- и тритонимфы последовательно появляются двенадцатый и тринадцатый сегменты брюшка, а на девятом и десятом сегментах - видоизмененные ножки. Последние одни за другими прирастают сзади к ранее появившимся, образуя половые крышки с тремя парами коксальных органов под ними. При последней линьке и превращении тритонимфы во взрослого клеща новых сегментов и конечностей уже не появляется, а образуется только половой конус.
Жизненная форма низших Acariformes очень своеобразна и не меняется от личинки до взрослого клеща. Всю жизнь эти клещи обитают во влажной почве, в перегнивших растительных остатках, в пространствах между частичками, и не выработали каких-либо специальных расселительных фаз, свойственных другим клещам. Они чрезвычайно влаголюбивы и могут жить только в воздухе, насыщенном водяными парами. Извлеченные из почвы, они быстро высыхают и гибнут. Но не менее опасна и капельная влага. Если клещ прилипнет к поверхностной пленке, он намокает и гибнет. Условия жизни этих крошечных дышащих кожей существ лежат на грани воды и пара. С одной стороны, угроза высохнуть, с другой - безнадежно прилипнуть к воде и размокнуть. Эти клещи так малы, что силы поверхностного натяжения воды для них резко преобладают над земным тяготением. Их стихия - силы молекулярного сцепления. Отсюда понятны и некоторые приспособления - несмачивающиеся покровы и прыгательные задние ноги у целого ряда форм. Последнее требует пояснений. Наблюдая этих клещей под микроскопом на поверхности воды, можно видеть, как они беспомощно перебирают ногами, скользя и не двигаясь с места. А прорвать коготками поверхностную пленку опасно: мениски намертво схватят за ноги, и тогда конец. Но вот удар задними ногами - и клещ легко отрывается, перескакивая на новое место.
Заметим, что, вероятно, в том же состоит истинный смысл прыгательного аппарата низших бескрылых насекомых - отряда ногохвосток (Collembola), которые обитают совместно с низшими клещами и представляют ту же жизненную форму мелкого тонкокожего несмачиваемого прыгающего жителя влажных пористых субстратов.
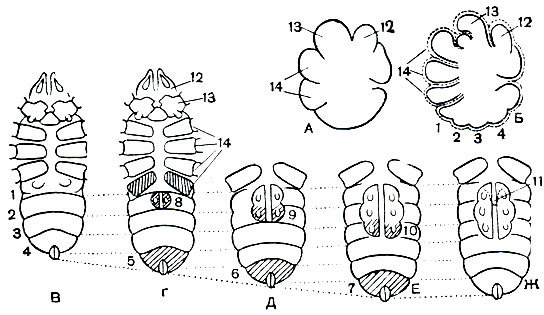
Рис. 55. Индивидуальное развитие акариформных клещей (исходный тип): А - зародыш на четырехсегментной стадии, вид сбоку; Б - шестиногий зародыш с неполным числом брюшных сегментов в момент выделения первой кутикулы (предличинка); В - личинка; Г-Е - прото-, дейто- и тритонимфа; Ж - взрослая фаза; 1-7- сегменты брюшка; 8-10 - половые крышки с коксальными органами под ними; 11 - половой конус; 12 - хелицеры; 13 - педипальпы; 14 - ноги (четвертая пара намечается уже у личинки). Послезародышевые фазы изображены с брюшной стороны, концевые отделы ног не показаны, сегменты и придатки, вновь образующиеся на каждой фазе, заштрихованы
Интересно, что некоторые виды тех и других выходят из почвы на поверхность стоячих водоемов. Низшие акариформные клещи и ногохвостки живут бок о бок с незапамятных времен, они найдены вместе в девонских отложениях, которым 300 миллионов лет, и, вероятно, были одними из первых членистоногих, заселивших почву на нашей планете.
Таково исходное состояние отряда Acariformes. Выше мы говорили о роли малых размеров в эволюции клещей. В судьбе данного отряда роль малых размеров особенно отчетлива и двояка. С одной стороны, благодаря малым размерам эти древнейшие клещи остановились на более низком эволюционном уровне, чем другие арахниды. Обогащение яиц желтком, столь характерное для хелицеровых вообще, здесь затормозилось малой величиной самки. В результате сохранился архаичный способ развития (шестиногая личинка, анаморфоз) и примитивные черты строения (полнота сегментации и др.), о чем свидетельствуют низшие представители отряда.
Но, с другой стороны, акариформные клещи в течение огромного периода времени их жизни на суше максимально использовали приспособительные возможности, которые открылись перед ними благодаря малым размерам. Они в изобилии заселили почву и всевозможные пористые органические субстраты, из почвы устремились на растения и животных в качестве паразитов, проникли даже в водоемы. При этом замечательно, что архаичный метаморфоз, о котором мы только что говорили, явился той основой, на которой возникли приспособления к новым условиям. По мере необходимости отдельные фазы метаморфоза брали на себя те или иные жизненные функции: одни стали питательными, другие расселительными или переживающими неблагоприятную обстановку; даже размножение, в зависимости от условий жизни, оказалось возможным и целесообразным на разных фазах индивидуального развития. Благодаря малым размерам взрослого клеща и отсутствию необходимости длительного роста возникли чрезвычайно быстро размножающиеся формы - тот скоростной тип развития, о котором мы упоминали.
Таким образом, огромное разнообразие акариформных клещей возникло на основе различных приспособительных изменений их жизненного цикла, причем эти приспособления складывались на более низком эволюционном уровне, чем тот, на котором находятся другие арахниды.
На столбовой дороге эволюции, по которой прошли хелицеровые животные, акариформные клещи раньше других остановились и рассыпались по боковым тропкам. Поскольку в основе эволюции клещей лежат приспособительные изменения индивидуального развития, мы составили сравнительную схему жизненных циклов для данного отряда в целом, к которой нам постоянно придется обращаться по ходу дальнейшего изложения материала (рис. 56). Следует сразу же отметить, что при сравнительном рассмотрении жизненных циклов клещей, помимо специализации отдельных фаз развития, усложнения метаморфоза, а также живорождения, мы столкнемся с такими несколько непривычными явлениями, как уподобление взрослой фазы молодым (педоморфоз), сдвиг половой зрелости на ранние фазы цикла и отпадение поздних (неотения), выпадение промежуточных фаз, сверхраннее вылупление из яйца (дезэмбрионизация) и размножение в полуэмбриональном состоянии и т. п.
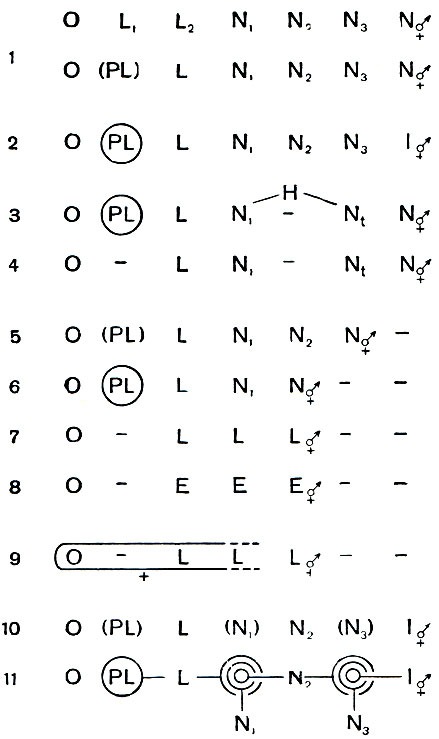
Рис. 56. Эволюция индивидуального развития клещей отряда Acariformes: 1 - исходные циклы (Endeostigmata, Palaeacaroidea); 2 - панцирные клещи (имагинизация взрослой фазы); 3 - тироглифоидные клещи (преобразование дейтонимфы в гипопуса); 4 - паразитические акаридии; 5 - почвенные простигматические клещи; 6 - тетраниховые клещи; 7 - шестиногие клещи; 8 - галлообразующие клещи; 5-8 - ряд нарастания неотении - размножения во все более раннем возрасте; 9 - тарсонемиды (утробное развитие, живорождение); 10 - краснотелки (повторный гистолитический метаморфоз); 11 - водяные клещи гидракарины (двойные линьки); О - яйцо; L - личинка; PL, взятое в скобки,- желточное питание предличинки; PI, в кругу - пред-личинка в яйце; N1-N3 - прото-, дейто- и тритонимфа; N1, N3, взятые в скобки,- покоящиеся нимфы; Н - гипопус; Е - пол у эмбриональное состояние; I - имаго; - - выпадение фаз; значком 'самка, самец' обозначены размножающиеся фазы
Подотряд Саркоптиформные клещи (Sarcoptiformes)
Представителей этого подотряда можно назвать грызущими или жующими акариформными клещами, так как большинство их обладает клешневидными хелицерами с крепкими клешнями жующего типа. Питаются они главным образом твердой растительной пищей, а переходя к паразитизму на позвоночных животных, питаются кожей, пером, волосом, выделениями кожных желез. В этот подотряд входят две большие серии семейств - панцирные клещи, или орибатиды (Oribatei), и акаридии (Acaridiae). В пределах каждой серии семейства объединяются в ряд надсемейств. Серии отражают два главных направления эволюции жизненного цикла в этом подотряде. У орибатид взрослая фаза развития становится панцирной и, помимо размножения, выполняет функцию расселения и переживания неблагоприятных условий. У свободноживущих акаридии такой расселительной и переживающей фазой становится дейтонимфа - так называемый гипопус, свойственный относящимся сюда тироглифоидным клещам (надсем. Tyroglyphoidea). При переходе акаридии к паразитизму переживающая дейтонимфа обычно выпадает. Таковы перьевые клещи (надсем. Analgesoidea), чесоточные клещи (надсем. Sarcoptoidea) и ряд других.
Панцирные клещи (Oribatei) - это самая обширная группа среди почвенных клещей, включающая, по разным авторам, от 40 до 95 семейств. Главная особенность орибатид, благодаря которой они распространились повсюду и достигли громадного видового разнообразия, состоит в том, что у взрослой фазы образовались твердый панцирь и трахеи, взрослый клещ стал устойчивым к внешним воздействиям, долговечным и способным активно переселяться. Предшествующая часть жизненного цикла сохранилась при этом почти без изменений (рис. 56,2). Личинка и нимфы лишены панциря и трахей и вынуждены жить во влажном воздухе в толще субстрата.
Заключенное в панцирь тело взрослого клеща разделено на протеро- и гистеросому, обычно сращенные неподвижно. Но в группе Ptyctima протеросома может подгибаться, под нее прячутся ноги и панцирь замыкается в виде гладкого шарика (рис. 57, 9, табл. 8, 10). Ротовые органы - короткие хелицеры с грызущими клешнями и небольшие педипальпы - спрятаны в углублении, образованном краями щита, и могут прикрываться снизу сросшимися в пластинку тазиками педипальп. Ноги крепкие, их членики иногда четковидные, на лапках 1-3 коготка. Тазики ног срастаются с панцирем. Снизу на гистеросоме два отверстия, герметически закрывающиеся парными створками. Заднее - анальное, из переднего выпячивается наружный половой аппарат. Половой конус самки обычно превращен в крупный яйцеклад, у самца половой конус служит для выведения сперматофоров. У основания конуса под створками имеются три пары пальчатых коксальных органов. Хорошо развиты покровные органы чувств (щетинки, соленидии на ногах и др.); глаз нет. Характерна пара видоизмененных, нередко булавовидных щетинок - трихоботрий на протеросоме. С их помощью воспринимаются колебания, причем орибатиды чутко реагируют на прикосновение и сотрясение субстрата, отвечая замиранием (танатозом). Дышат взрослые клещи трахеями, которые имеют вид тонких неветвящихся трубочек и открываются четырьмя парами стигм в местах причленения ног.
Взрослую фазу панцирных клещей обычно называют имаго - термином, который требует пояснений. Этот термин, в переводе означающий "истинный облик", возник в энтомологии для обозначения взрослого крылатого насекомого и подчеркивает его отличия от нелетающих предимагинальных фаз, особенно в случае превращения червеобразной личинки в крылатую форму. Но в дальнейшем понятие "имаго" стало попросту синонимом размножающегося насекомого и, проникнув в таком виде в акарологию, лишилось всякого смысла. Однако это понятие слишком укоренилось, чтобы можно было отказаться от него, как это пытаются делать некоторые акарологи. Полезнее его уточнить, и тогда оно приобретет важное значение не только для клещей, но и для членистоногих вообще. Способность размножаться не есть еще основание называть взрослую фазу имагинальной. Об имаго мы будем говорить в том случае, когда взрослая фаза наряду с размножением берет на себя функцию расселения и переживания, выходит из тех более постоянных условий, в которых развиваются предимагинальные фазы, становится способной переносить превратности открытой жизни, прежде всего сухость, вырабатывает в связи с этим специальные приспособления (панцирь, трахеи и др.) и становится отличной от предшествующих фаз развития. Превращение взрослой фазы в имаго (имагинизация) есть эволюционный процесс, связанный у членистоногих с освоением суши. На этом пути возникла крылатая форма насекомых. Панцирная имагинальная фаза формируется в ряде групп клещей. Заметим, что у клещей при переходе к паразитизму на животных и растениях, особенно к скрытой жизни в полостях и тканях, мы встречаем и обратный процесс - утрату имагинальных свойств взрослой фазы (дезимагинизацию), вплоть до ее исчезновения и размножения на более ранних фазах развития.
Формирование панцирного имаго хорошо прослеживается у орибатид (рис. 57). У более примитивных форм (сем. Нуроchthoniidae и др.), близких к упоминавшимся выше клещам-палеакаридам, панцирь еще не так прочен, желтоватый или розоватый и состоит из отдельных щитков, сохраняя следы сегментации. У большинства орибатид щиты сплошные, темно-бурые или почти черные, твердые, блестящие. В ряде случаев панцирь укрепляется и усложняется дополнительными образованиями. Он бывает чрезвычайно прочен и покрыт скульптурным орнаментом (сем. Carabodidae и др.). На протеросоме обычно имеются килевидные возвышения, а над основаниями ног - твердые выступы. Нередко по бокам тела образуются крыловидные выросты (птероморфы). Обычно они неподвижны, но у так называемых крылатых клещей (сем. Galumnidae) причленены подвижно и приводятся в движение мышцами (рис. 57, 7). На ходу клещ машет ими, как крыльями, а при замирании прикрывает поджатые ноги. Эти образования даже сравниваются с боковыми (паранотальными) выростами груди древних насекомых,от которых, как считают, произошли крылья.
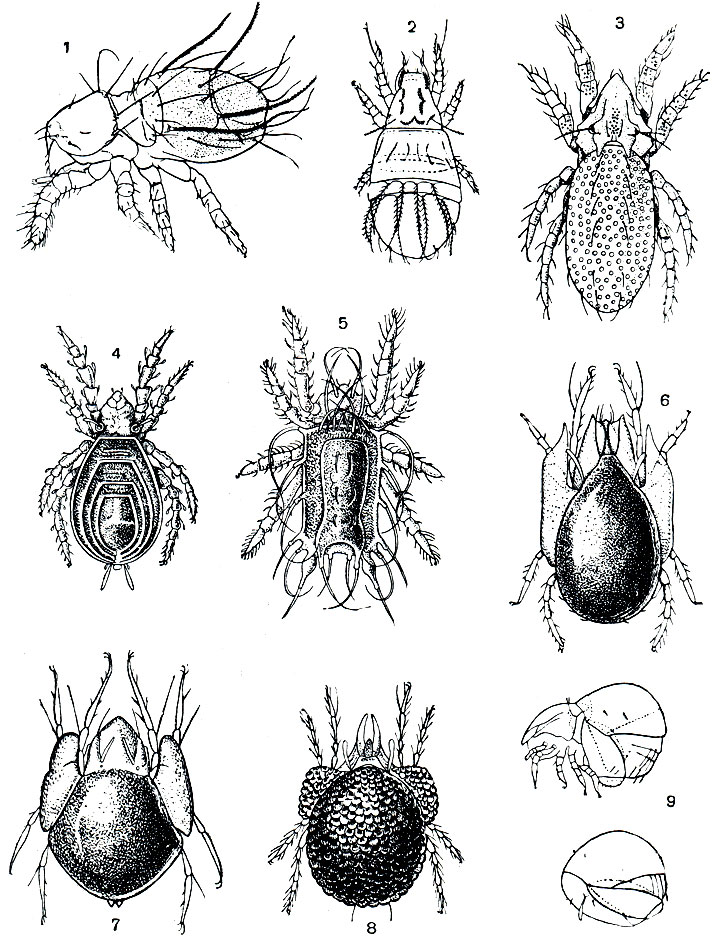
Рис. 57. Панцирные клещи: 1 - Beklemishevia galeodula - представитель примитивного надсемейства палеакарид; 2 - Cosniochthonius plumatus; 3 - Eulohmannia ribagai; 4 - Camisia spinifer; 5 - Platyliodes daderleinii; 6 - Notaspis nicoletii; 7 - Galumna mucronata; 8 - Phaenopelops variotosus; 9 - Aedoplophora glomerata; внизу - клещ с поджатой протеросомой
По мере развития имагинальных признаков взрослая фаза орибатид становится все более стойкой и долговечной. При высыхании субстрата взрослые орибатиды гораздо дольше остаются живыми, чем большинство других клещей, а продолжительность жизни имаго в благоприятных условиях достигает 15-18 месяцев и более. Различны характер размещения орибатид в толще субстрата и их расселительные способности. У видов со слаборазвитым панцирем взрослая фаза держится в толще влажного субстрата подобно личинкам и нимфам. У большинства орибатид взрослые формы преобладают в верхних слоях почвы и в растительных остатках, а при повышении влажности воздуха, в дождливое время, при туманах и росах, выходят на поверхность и поднимаются по растительности. Есть среди орибатид формы, хорошо приспособленные к активному расселению, например длинноногие клещи семейства Damaeidae, по жизненному облику удивительно похожие на мелких панцирных сенокосцев, в частности рода Nemastoma. Это такие, же закованные в панцирь, дышащие трахеями шагающие существа способные, долгое время проводить вне толщи влажных субстратов. Заметим, что все, что говорилось о биологическом значении имаго орибатид, вполне приложимо к панцирным сенокосцам, хотя в отношении последних это понятие до сих пор как-то не употреблялось.
Орибатиды встречаются во всех ландшафтно-климатических зонах, но наиболее обильны и разнообразны они в лесных почвах и гниющей подстилке, особенно во влажных районах. Их численность в таких местах бывает огромна - десятки тысяч экземпляров в одном кубическом дециметре субстрата. Орибатиды теснейшим образом связаны с микрофлорой. По типу питания это жующие сапрофаги, потребители гниющих растительных остатков с обильной микрофлорой. Вместе с детритом они поедают бактериальные налеты, дрожжи, гифы и споры грибов, почвенные водоросли и т. п. На туловище и ногах у некоторых видов орибатид нередко образуются целые сады из грибных мицелиев и бактериальных колоний (табл. 8, 7). По-видимому, эти обрастания полезны для клещей, особенно тонкокожих нимф, и защищают их от потери влаги.
Цикл развития длительный, обычно занимает более месяца, а общая продолжительность жизни, считая срок жизни имаго, составляет не меньше года, а иногда, по-видимому, несколько лет. Оплодотворение сперматофорное, причем строение сперматофоров довольно сложно (рис. 58, 1). На длинном стержне имеется чашевидное образование, в котором помещается мешочек со спермиями, нередко снабженный дополнительными придатками. Яйца откладываются с помощью яйцеклада (полового конуса) в толщу субстрата, но иногда самки носят яйца на себе (рис. 58, 2). У некоторых видов наблюдается очень своеобразное явление, получившее название посмертного живорождения. Самка погибает, не отложив яиц, которые затем развиваются в трупе матери. Вылупившиеся личинки выходят наружу, обычно раскрывая анальные створки или выгрызая и выталкивая из панциря ротовые органы. Плодовитость в этом случае невелика, но зато развивающиеся яйца надежно защищены материнским панцирем. Личинки и нимфы тонкокожие, лишены трахей и живут во влажных субстратах. У ряда видов линечные шкурки не сбрасываются полностью, а остаются прикрепленными на спине клеща и служат дополнительной защитой. В результате к моменту образования имаго на спине имеется целая горка шкурок: сверху личиночная, а ниже - прото-, дейто- и тритонимфальная (рис. 58, 5, табл. 8, 8).
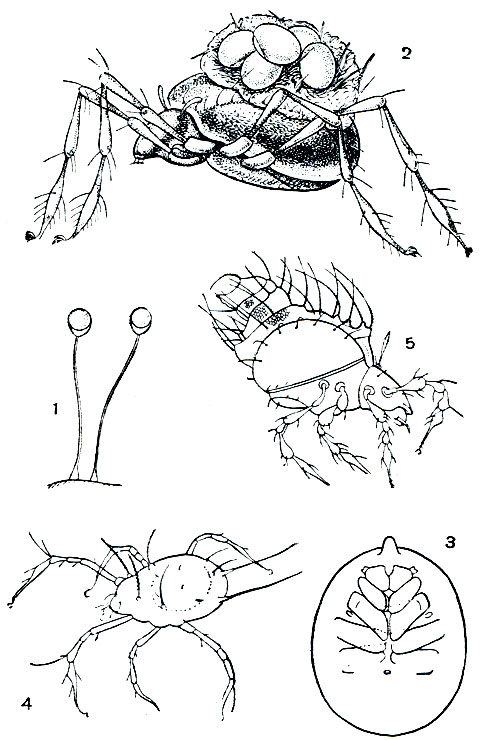
Рис. 58. Размножение и развитие панцирных клещей: 1 - сперматофоры; 2 - самка Damaeus с яйцами на спине; 3 - предличинка; 4 - личинка; 5 - Damaeus jacoti с линечными шкурками на спине
Практическое значение орибатид разнообразно. Будучи массовыми потребителями разлагающихся растительных остатков, они играют немалую роль в процессе почвообразования, что подтверждено наблюдениями и опытами. Тесная связь орибатид с микрофлорой и способность подниматься на растения заставляют подозревать в них один из факторов распространения фитопатогенных микроорганизмов и грибов, в частности поражающих древесину.
Но наиболее существенна роль орибатид как промежуточных хозяев ленточных глистов - цестод группы Anoplocephalata, поражающих скот и ценных промысловых животных. Среди этих гельминтозов наиболее распространен мониезиоз жвачных животных, главным образом молодняка, вызываемый паразитированием в кишечнике цестоды Moniezia expansa. Больные животные плохо растут, снижается удой и качество мяса, нередко молодняк гибнет. Цикл развития мониезии долгое время не могли расшифровать, и лишь в 1937 году американский гельминтолог Стэнкард установил, что промежуточными хозяевами этих глистов в природе служат орибатиды. Яйца мониезии выводятся с испражнениями животных и поедаются орибатидами, живущими в почве пастбищ. В кишечнике клеща из яиц выходят личинки - онкосферы, которые проникают в полость тела и превращаются в цистицеркоида - инвазионную стадию, способную заражать животное - окончательного хозяина (рис. 59). Животные поедают клещей с травой на пастбищах. В кишечнике жвачных панцири клещей разрушаются, из них выходят цистицеркоиды, которые проникают в тонкий кишечник и здесь присасываются к стенке, постепенно вырастая затем во взрослого лентеца. Имагинальная фаза орибатид словно самой природой предназначена быть промежуточным хозяином мониезии. Питание гниющими веществами и заглатывание с ними яиц гельминтов, долговечность, способность выходить из почвы на траву - все это обеспечивает протекание цикла гельминтов и стойкое сохранение очагов мониезиоза.
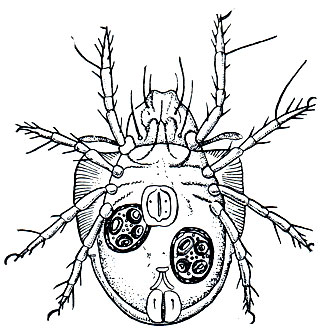
Рис. 59. Панцирный клещ Scheloribates laevigatas с цистицеркоидами мониезии в полости тела
Выяснение ветеринарного значения орибатид стимулировало изучение этих клещей за рубежом и у нас. Советскими гельминтологами расшифрованы циклы ряда аноплоцефалят, установлены виды орибатид - промежуточных хозяев этих гельминтов- и разработаны меры защиты сельскохозяйственных животных, в частности рациональная система выпаса с учетом размещения и численности клещей на пастбищах. Теперь известно более 50 видов орибатид, участвующих в распространении гельминтозов. Среди них крылатые клещи рода Galumna (сем. Galumnidae), виды рода Scheloribates (сем. Scheloribatidae) и др. Значительно продвинулось изучение фауны и разработка систематики орибатид. В нашей литературе появились руководства по этим клещам и их ветеринарному значению: В. А. Потемкиной "Мониезиозы жвачных животных" ("Колос", 1965); Е. М. Булановой-Захваткиной "Панцирные клещи-орибатиды" ("Высшая школа", 1966).
Тироглифоидные клещи (надсем. Тугоglyphoidea) называются также амбарными, так как среди них немало форм, повреждающих зерно в хранилищах, муку и другие пищевые продукты. Тироглифиды по количеству видов уступают орибатидам, но численность их в природе и в запасах продуктов бывает огромна. Заслуга всестороннего изучения тироглифид и разработки мер защиты пищевых запасов принадлежит коллективу советских ученых во главе с профессорами Московского университета З. С. Родионовым и А. А. Захваткиным. Последним написана монография "Тироглифоидные клещи" ("Фауна СССР", 1941).
Наиболее характерная особенность тироглифид - преобразование дейтонимфы в переживающую фазу - гипопус, чрезвычайно устойчивую к внешним воздействиям (рис. 60, 3). Благодаря этому выработался своеобразный и весьма совершенный ритм жизни. Тироглифиды легко заселяют всевозможные органические субстраты и благодаря многоядности и высокой размножаемости в короткий срок достигают колоссальной численности. При наступлении неблагоприятных условий все активные фазы погибают, но остаются гипопусы, которые не питаются, не боятся сухости и, попадая в подходящую обстановку, дают начало новой колонии клещей. Поэтому тироглифиды чрезвычайно жизнеспособны и широко распространены, буквально вездесущи. Где есть немного влаги и органического вещества, там обязательно окажутся те или иные виды этих клещей.
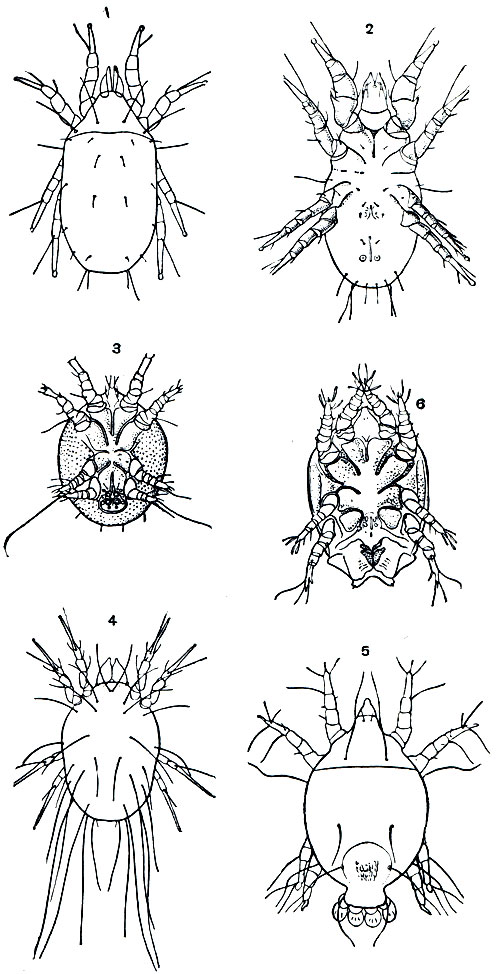
Рис. 60. Тироглифоидные клещи: 1 - мучной клещ (Tyroglyphus farinae), самка; 2 - то же, самец с брюшной стороны; 3 - то же, энтомохорный гипопус с брюшной стороны; 4 - Glycyphagus destructor; 5 - винный клещ (Histiogaster bacchus), самец; 6 - терохорный гипопус клеща Labidophorus desmanae из нор выхухоли
Строение питающихся фаз и гипопусов резко различно. Взрослые клещи обычно 0,3-0,5 мм длиной, туловище овальное, покровы тонкие, глянцевитые, несмачивающиеся благодаря выделениям жировых желез, открывающихся по бокам тела. Окраска белесоватая, сквозь покровы просвечивает запасная ткань, богатая жиром. Ноги и ротовые органы более темные. На теле сидят щетинки, постоянные по числу, но очень различные по длине. Глаз нет. Основные членики коротких педипальп сращены в пластинку, хелицеры с грызущими клешнями. Наружный половой аппарат видоизменен в связи со своеобразным способом копуляции. Под половыми клапанами у самки открывается яйцевыводное отверстие, а у самца расположен копулятивный орган. При спаривании самец прикрепляется к самке сзади под углом, сперма вводится в копулятивную пору, расположенную у самки позади анального отверстия и связанную канальцем с ее половыми путями.
Плодовитость сильно варьирует у разных видов, она зависит от количества поглощенной самкой пищи и обычно составляет 2-3 десятка яиц, но иногда более 600. При благоприятных условиях весь цикл развития занимает меньше недели. Очень мелкая (0,1-0,2 мм) личинка превращается в протонимфу, которая при высокой влажности и избытке пищи, обычно минуя фазу гипопуса, сразу превращается в тритонимфу, а последняя - во взрослого клеща. В связи с тем что фаза дейтонимфы (гипопус) в этом случае выпадает, тритонимфа у тироглифид получила особое название - телеонимфа (рис. 56, 3, 4).
Гипопусы тироглифид бывают покоящиеся и расселительные. Первые остаются в субстрате и "выжидают" изменения условий к лучшему. Такие гипопусы нередко почти лишены придатков и внешне похожи на яйцо или цисту в плотной оболочке. Расселительные гипопусы имеют щитообразное тело, твердый панцирь, короткие ноги, недоразвитые ротовые органы и прикрепительные приспособления. Различают энтомохорных гипопусов, т. е. расселяющихся на насекомых, и терохорных, расселяющихся на позвоночных животных, чаще на грызунах. У первых снизу имеется прикрепительный диск с несколькими парами присосок - видоизмененных щетинок. С их помощью гипопус прикрепляется к гладкой кутикуле насекомых, причем присоски действуют пневматически подобно присоскам выпускаемых теперь мыльниц и вешалок, прикрепляемых к кафельным плиткам. У терохорных гипопусов на месте диска имеются две пары хитиновых тисков, зажимающих волос животного-транспортера. Гипопусы расселяются на тех видах насекомых, которые развиваются в субстратах, заселенных клещами, и на случайных посетителях. Они поджидают транспортеров на поверхности субстрата и мгновенно прицепляются к подвижному предмету. Достаточно поднести иглу, как эти крошечные существа облепят ее конец. На грызунах расселяются виды, живущие в их гнездах и запасах. Гипопусы тироглифид, живущих в муравейниках, термитниках, пчелиных ульях, расселяются на соответствующих насекомых-хозяевах. При этом на теле насекомого нередко обнаруживается несколько сотен гипопусов.
Механизм появления гипопусов окончательно не разгадан и таит важные общебиологические закономерности. Явление это во многом напоминает наступление диапаузы у насекомых, но отличается тем, что у последних состояние диапаузы приурочено к обычным фазам цикла - яйцу, личинке, куколке или имаго, а у тироглифид выражается в образовании особой фазы - гипопуса. Сходны с диапаузой и варианты образования гипопусов. В одних случаях ряд поколений следует без гипопуса, а затем при ухудшении условий - сухости, недостатке пищи и т. п. - протонимфы начинают превращаться не в телеонимф, а в гипопусов, причем превращение последних в телеонимф в дальнейшем происходит довольно быстро, как только обстановка меняется к лучшему. У других видов гипопусы появляются заранее, как бы "зная", что в дальнейшем условия станут неблагоприятными и для их превращения в телеонимф требуется длительное время, уже независимо от того, есть подходящая для дальнейшего развития обстановка или нет. Наконец, есть виды, у которых гипопус образуется в каждом поколении.
Стойкость гипопусов поразительна и обусловлена, помимо защиты панцирным покровом, пониженным уровнем обмена. Гипопусы выносят высушивание и промораживание, действие ядовитых веществ, выдерживают высокие дозы ионизирующей радиации, смертельные для других организмов.
Сравнивая циклы развития тироглифид и орибатид, легко заметить, что те функции расселения и переживания, которые у орибатид взяла на себя взрослая имагинальная фаза, у тироглифид выполняет гипопус. Поэтому у тироглифид взрослая фаза не имеет характера имаго, это, по существу, размножающаяся нимфа, причем в тех случаях, когда гипопус выпадает, и число линек сокращается на одну по сравнению с исходным циклом (рис. 56, 3).
Тироглифиды распространяются не только в фазе гипопуса. Некоторые виды с длинными щетинками поднимаются токами воздуха на огромные высоты. Яйца тироглифид, проглоченные грызунами, не погибают и рассеиваются с экскрементами. Особенно способствует распространению этих клещей человек при перевозках продуктов.
Места обитания тироглифид очень разнообразны: почва, лесная подстилка, скопления всевозможных растительных остатков, гниющая древесина, вытекающий сок деревьев, грибы, лишайники, мхи, корни, клубни и зеленые части высших растений, гнезда млекопитающих, птиц, насекомых и т. п. Характерный набор видов заселяет зерно в элеваторах и складах, куда клещи заносятся с полей при уборке урожая. При влажности зерна около 17% клещи выедают зародыш, к которому проникают через повреждения оболочки, а при более высокой влажности начинается их массовое размножение в зерне, причем они питаются и эндоспермом. Помимо непосредственного повреждения, клещи снижают всхожесть зерна, засоряют его экскрементами и шкурками, приводят к склеиванию в комки и самосогреванию, заражают гнилостными микроорганизмами. Наиболее серьезные вредители зерна - мучной клещ (Tyroglyphus farinae) и удлиненные клещи (Tyrophagus noxius, T. perniciosus). Муке сильно вредит Aleuroglyphus ovatus.
Защита зерна от амбарных клещей включает ряд мероприятий. Расселительные способности тироглифид не позволяют полностью предотвратить их попадание в склады. Поэтому особое значение приобретает режим хранения, исключающий размножение клещей: охлаждение зерна и поддержание низкой его влажности, устранение факторов, вызывающих самоувлажнение и самосогревание, доступ сырого воздуха и т. п. Из химических мер борьбы применяют обработку зерна газообразным хлорпикрином и порошкообразными ядами (акарицидами), но необходимость этих "пожарных" мер обычно бывает результатом плохих условий хранения запасов.
Тироглифиды вредят не только зерну и мучным продуктам. Сырный клещ (Tyrolichus casei) размножается на сырах; клещ Carpoglyphus lactis живет на веществах, содержащих молочную, уксусную и янтарную кислоты,- сухих фруктах, кислом молоке, старом сыре, пиве, гнилых овощах и т. п. Интересен винный клещ (Histiogaster bacchus), который иногда в массе размножается на поверхности вина в чанах разливочных цехов винных заводов и портит вино. Виды этого рода живут в природе на вытекающем бродящем соке деревьев, особенно дуба. Как выяснилось, эти клещи попадают на винные заводы с дубовыми бочками, в которых транспортируется вино. Луковичный клещ (Rhizoglyphus echinopus) живет на корнях растений и сильно повреждает луковицы, корне- и клубнеплоды в хранилищах.
Тироглифиды патогенны для человека. При проглатывании с пищей они могут вызвать острые желудочно-кишечные заболевания, а при вдыхании - катар дыхательных путей и астматические явления. С тироглифидами связано и немало курьезных случаев в практике медицинских лабораторий. Живя повсюду, эти клещи обнаруживаются при самых неожиданных обстоятельствах. Автору этого раздела не раз приходилось определять тироглифид, доставленных из лабораторий в моче и крови человека, взятых для анализа, и даже "найденных" в тканях трупов при вскрытии. Все это, конечно, результат лабораторного заноса клещей. Однако есть некоторые виды, правда недостаточно изученные, которые встречаются при анализах мочи регулярно и, по-видимому, могут поселяться в мочеполовых путях человека.
Паразитические акаридии представлены несколькими надсемействами: перьевые клещи птиц (надсем. Analgesoidea), паразиты млекопитающих - волосяные клещи (надсем. Listrophoroidea) и чесоточные клещи (надсем. Sarcoptoidea), некоторые паразиты насекомых и др. Все они по происхождению связаны с тироглифоидными клещами и перешли к паразитизму через гнездовое сожительство с животными-хозяевами. Это постоянные наружные или внутрикожные паразиты с грызущими ротовыми органами, питающиеся не кровью, а пером, волосом, кожей, выделениями кожных желез и т. п. Переход к постоянному паразитизму сопровождался выработкой ряда приспособлений. Расселительная дейтонимфа (гипопус) исчезла за ненадобностью. У наружных паразитов (эктопаразитов), подвергающихся механическому воздействию хозяина - счесыванию и раздавливанию, возникли защитные и прицепные приспособления: уплощенное тело, покрытое щитками, крепкие ноги с крупными присосками на лапках, выросты и разнообразные прицепные щетинки, прикрепительный аппарат для захвата волоса у волосяных клещей и т. п.
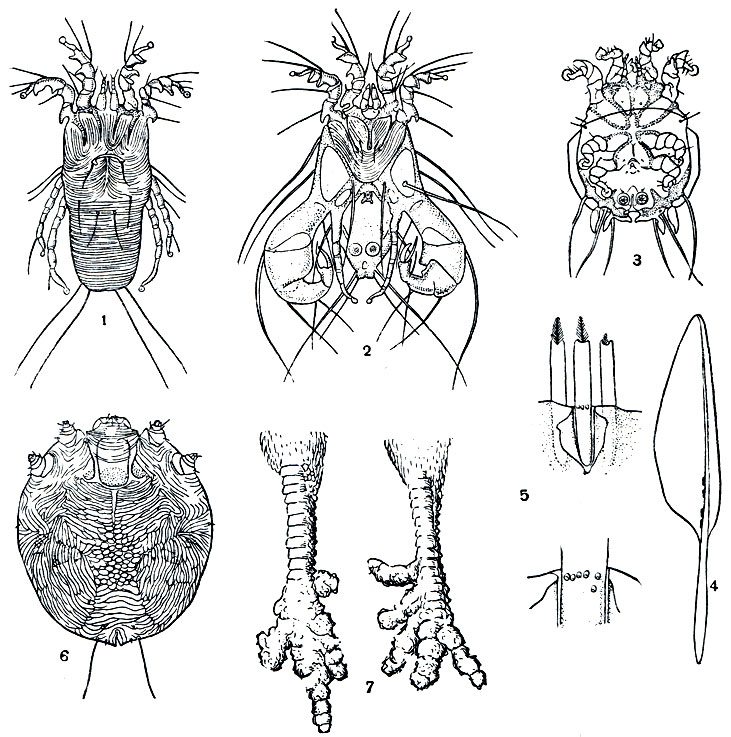
Рис. 61. Перьевые клещи: 1 - Analgopsis passerinus с перьев воробья, самка; 2 - то же, самец; 3, 4 - перьевой клещ уток Freyana anatina и его локализация на пере; 5 - клещи, спустившиеся в перовую сумку старого выпавшего при линьке пера и взобравшиеся на очин нового пера; 6 - ножной зудень Knemidocoptes mutans; 7 - тяжелое кнемидокоптозное поражение ног курицы с отпадением фаланг пальцев
Большой вклад в изучение паразитических акаридии внес наш известный акаролог В. Б. Дубинин, которому принадлежит монография по перьевым клещам (в нескольких томах "Фауны СССР", 1951, 1953, 1956) и книга "Чесоточные клещи" ("Советская наука", 1954).
Наиболее разнообразны перьевые клещи (надсем. Analgesoidea), которых только на птицах Палеарктики обнаружено около 1000 видов. Эти высокоспециализированные паразиты живут на бородках перьев и на коже птиц. На одном виде птиц обычно встречается несколько специфических видов перьевых клещей, причем каждый обитает на разных участках оперения, нередко на определенных перьях, с чем в свою очередь связана различная форма тела клещей и строение прицепных приспособлений. Перьевые клещи питаются отмершими частичками эпидермиса кожи и перьев, но главным образом жировой смазкой оперения птиц, выделяемой копчиковой железой. Самец спаривается с телеонимфой, у которой имеется копулятивный канал и которая после этого линяет, превращаясь в оплодотворенную самку. Яйца приклеиваются к бородкам пера. В большинстве случаев, кроме шестиногой личинки, имеется две нимфальные фазы - прото- и телеонимфа, но у некоторых видов бывает еще покоящаяся дейтонимфа, которая соответствует гипопусу и появляется при неблагоприятных условиях, например в период линьки птицы-хозяина. Течение жизненного цикла и все поведение перьевых клещей подчинены жизненному ритму птиц-хозяев и закономерно изменяются в зависимости от возраста птицы, наступления и характера линьки оперения, осенних перелетов и т. д.
Большинство перьевых клещей не причиняет птицам заметного вреда, но есть виды, вызывающие тяжелые поражения кожи. Таков ножной зудень (Knemidocoptes mutans), прежде причислявшийся к чесоточным клещам, который живет под чешуями неоперенной части ног кур и других домашних птиц и вызывает тяжелое заболевание "известковые ноги". При этом заболевании чешуи оттопыриваются, ноги покрываются белесыми бугристыми корками, под которыми происходит отмирание (некроз) тканей, и птица гибнет.
Чесоточные клещи (надсем. Sarcoptoidea) представлены немногими видами, паразитами млекопитающих животных и человека, возбудителями различных форм зудневой чесотки. У человека чесотку вызывает чесоточный зудень (Acarus siro). Именно этот паразит в трудах Аристотеля обозначается словом "клещ" (греческое ахара, латинское Acarus) и указывается в качестве причины чесотки.
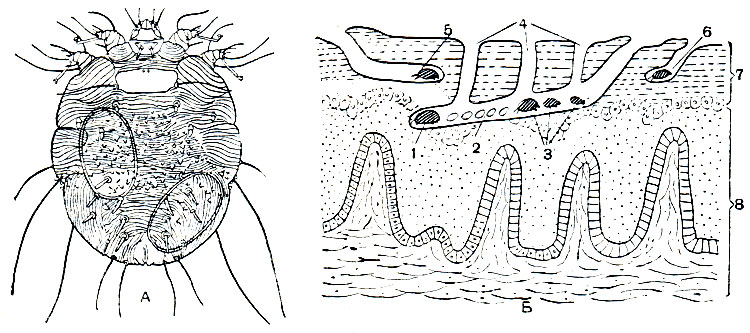
Рис. 62. Чесоточные клещи: А - чесоточный зудень (Acarus siro), самка; Б - схема расположения в коже маточного хода (в середине), хода самца (справа) и хода телеонимфы (слева) чесоточных клещей рода Acarus; 1 - самка; 2 - яйца; 3 - личинки и нимфы; 4 - вентиляционные отверстия; 5 - оплодотворенная телеонимфа; 6 - самец; 7 - роговой слой кожи; 8 - живые клеточные слои кожи
Самки чесоточного зудня 0,3 мм длиной, тело округлое с короткими ногами; самец вдвое мельче. Покровы кожистые, бороздчатые. Самка питается кожей, прогрызая в ее роговом слое извитые ходы до 15 мм длиной, которые различимы через поверхность кожи в виде сероватых линий. Яйца откладываются в ходах, над ними самка обычно выгрызает вентиляционные отверстия. Личинки и протонимфы живут в этих ходах, где питаются остатками изгрызенной самкой кожи и тканевой жидкостью. Сами они ходов не прогрызают. Протонимфы превращаются в телеонимф, которые выходят на поверхность кожи обычно ночью, когда больной спит. Здесь часть их превращается в самцов, которые спариваются с женскими телеонимфами - будущими самками. Самцы выгрызают в коже небольшие ходы, в которые прячутся. Оплодотворенные телеонимфы вгрызаются в кожу и превращаются в самок. Заражение происходит при соприкосновении с больным чесоткой, в редких случаях через платье больного. Человека могут заражать и другие чесоточные клещи, которые паразитируют на собаках, овцах, козах, свиньях, лошадях и других животных. Чесоточные зудни родов Acarus, Psoroptes, Chorioptes сильно вредят в животноводстве.
Лечат чесотку химическими препаратами в виде мазей, растворов; животных купают в ваннах или обрабатывают газообразными препаратами в специальных камерах.
Подотряд Тромбидиформные клещи (Trombidiformes)
Представителей этого подотряда, в отличие от предыдущего, можно назвать сосущими акариформными клещами, так как большинство их питается жидкой животной или растительной пищей и хелицеры приспособлены для прокалывания. Характерно развитие пары трахей, открывающихся стигмами близ хелицер. Классификация подотряда сложна, насчитывается до 20 надсемейств и 70 семейств. Многочисленные ряды форм ведут от почвенных клещей к разнообразным обитателям растений, паразитам животных, водяным клещам и др. Судя по характеру жизненных циклов, все это на первый взгляд необозримое многообразие представляет два главных эволюционных направления, ведущих начало от примитивных Endeostigmata. Одно из них характеризуется сокращением жизненного цикла, поздние фазы развития уподобляются ранним (педоморфоз), число линек сокращается, и размножение происходит во все более раннем возрасте (неотения), что обусловлено монотонностью условий и скрытым образом жизни этих форм на протяжении цикла развития. Таковы многие семейства почвенных Trombidiformes, обычно объединяемые в группу Prostigmata, у которых взрослые особи отвечают тритонимфам исходного цикла, и связанные с ними по происхождению постоянные обитатели растений - тетраниховые и галлообразующие клещи, а также ряд постоянных паразитов животных.
Другое направление представляют краснотелки и пресноводные клещи, у которых возник личиночный паразитизм, резкая смена образа жизни в течение цикла и связанный с этим сложный метаморфоз.
Почвенные Trombidiformes (табл. 8, 1-6) чрезвычайно разнообразны. Располагая микроскопом или лучше бинокуляром и несложным прибором для добывания клещей, можно увидеть эти формы. Клещей "выгоняют" из почвы с помощью термоэклектора. Это воронка с вложенным в нее ситом, на которое помещают почву или иной субстрат, подлежащий обработке. Над воронкой горит электрическая лампа, а под горлышко ставят чашечку с водой. По мере просушивания почвы лампой клещи и другие подвижные беспозвоночные уходят вниз, проваливаются сквозь сито и падают в чашечку. Часть их тонет, но через несколько часов поверхность воды покрывается бактериальной пленкой, по которой клещи свободно ходят. Если теперь поместить чашечку под бинокуляр, то глазам предстанет удивительный мир. Тут много уже знакомых нам орибатид и тироглифид, а также гамазоидных клещей, о которых речь ниже, но наиболее разнообразны тромбидиформные клещи. Здесь множество различных Prostigmata. По поверхности бегают крошечные беловатые клещи семейства Tydeidae, шустрые, подергивающие ногами клещи семейства Eupodidae. Вдруг среди этой мелюзги в поле зрения появляется крупный (в действительности всего 0,3 мм) лиловый клещ с острым "носом" - вытянутыми сомкнутыми хелицерами. Это бделла (сем. Bdellidae), хищница, пронзающая хелицерами, как копьем, ногохвосток и других клещей. Она очень прожорлива, ее нередко видишь с насаженной на хелицеры ногохвосткой, которую она высасывает на ходу. Вот другая хищница - рагидия (сем. Rhagidiidae), похожая на сольпугу в миниатюре, очень подвижная, нежная, но со здоровенными клешнями хелицер, жительница глубоких почвенных микропещер. Вот шествует бархатисто-красная краснотелка (сем. Trombidiidae), кажущаяся гигантом в сравнении с остальными. А вот удивительное нежнейшее существо с непомерно длинными ногами, подобие миниатюрного сенокосца - клещ Linopodes, живущий на грибах. Поверхность воды усеяна клещами-тарсонемидами (Tarsonemini), похожими на мельчайших черепашек, странными клещами-криптогнатидами (сем. Cryptognathidae) с капюшоном над ротовыми органами, которые выбрасываются на длинной "шейке", и множеством других форм. Здесь столько всего, что один перечень видов занял бы несколько страниц.
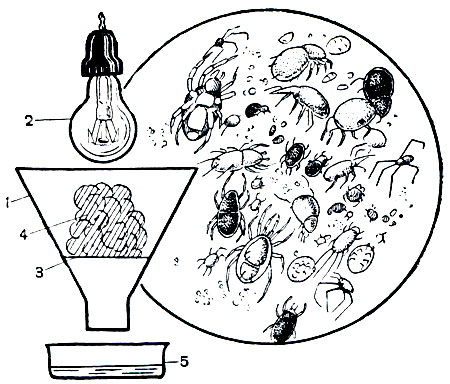
Рис. 63. Схема устройства термоэклектора: 1 - воронка; 2 - электрическая лампа; 3 - сито; 4 - почва; 5 - чашечка с водой; справа - клещи, извлеченные с помощью термоэклектора из почвенной пробы
Почвенные Trombidiformes дали начало множеству групп клещей, больших и малых по числу видов, из которых мы упомянем только некоторые. Своеобразны клещи-тарсонемиды (надсем. Tarsonemini), очень различные по образу жизни (рис. 64). Эти мельчайшие формы с сосущими ротовыми органами в виде "головки" и черепитчатыми щитками частью живут в почве, частью перешли на растения, насекомых и позвоночных в качестве паразитов. Размножение на ранних фазах (неотения) сочетается у них с утробным развитием молоди, вплоть до рождения взрослых клещей, иногда личинкоподобных. Злаковый клещ (Siteroptes graminum) повреждает злаки и переносит споры ржавчины. Пчелиный клещ (Acarapis woodi) паразитирует в трахеях пчел и сильно вредит в пчеловодстве. Клещи рода Pygmephorus живут в шерсти грызунов. Пузатый клещ (Pyemotes ventricosus) паразитирует на насекомых и может нападать на человека, вызывая уколами сильное раздражение кожи. Самка рождает взрослых клещей. В период размножения ее брюшко сильно вздуто и на нем держатся крошечные самцы, которые сосут гемолимфу матери и оплодотворяют рождаемых ею самок.
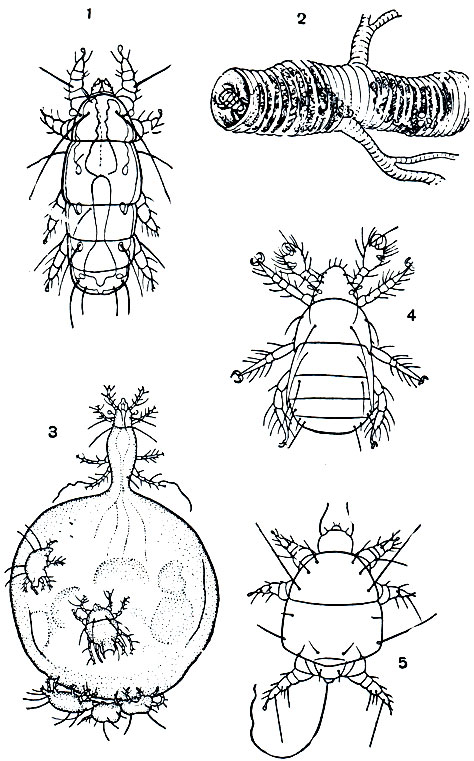
Рис. 64. Клещи-тарсонемиды: 1 - вредитель злаков Siteroptes graminum; 2 - трахея пчелы, больной акарозом, внутри клещи Acarapis woodi; 3 - пузатый клещ (Pyemotes ventricosus), беременная самка с раздутым брюшком, на котором держатся самцы; 4 - паразит грызунов Pygmephorus forcipatus; 5 - Locustacarus trachealis, живущий в трахеях саранчи, личинкоподобная самка
Другой пример перехода к паразитизму представляют простигматические клещи надсемейства Cheyletoidea (рис. 65). Хищные клещи (сем. Cheyletidae), с крупными хватательными педипальпами, живут в почве, растительных остатках, гнездах насекомых и позвоночных, нередко переходя на животных в погоне за клещами-паразитами. С этим семейством связано по происхождению несколько семейств своеобразных паразитов позвоночных. Клещи семейств Syringophilidae и Harpyrhynchidae живут в полостях очинов перьев, на коже и в подкожной клетчатке птиц. Волосяные клещи (сем. Myobiidae) паразитируют на мелких млекопитающих. Они держатся на волосе и высасывают содержимое волосяного фолликула, лимфу и плазму крови. От миобиид ведут начало железницы (сем. Demodicidae), которые живут в сальных железах и волосяных сумках млекопитающих и благодаря эндопаразитизму изменились до неузнаваемости. Они так малы, что их можно рассмотреть только при сильном увеличении микроскопа. Тело червеобразное, кутикула кольчатая, ноги короткие, как бы обрубленные, ротовые органы сосущие; они кладут яйца; личинки, нимфы и взрослые по облику сходны. Виды рода Demodex - паразиты разных млекопитающих, диких и домашних, некоторые сильно вредят в животноводстве, вызывая железничную чесотку (демодикоз). У человека это заболевание вызывает железница угревая (D. folliculorum), паразитирующая в сальных железах.
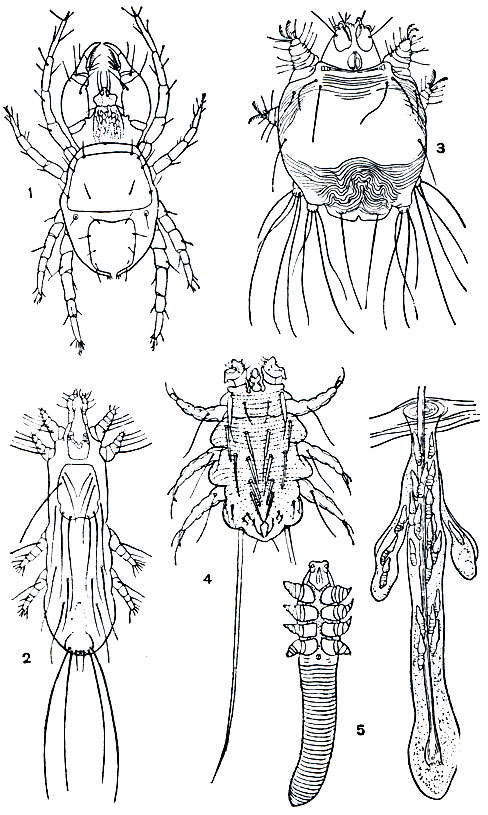
Рис. 65. Клещи хеилетоиды и железницы: 1 - хищный клещ Cheyletus eruditus; 2 - Syringophilus bipectinatus, живущий в очинах перьев птиц; 3 - Harpyrhynchus nidulans, паразитирующий во влагалищах перьев птиц; 4 - волосяной клещ грызунов Myobia muris-musculi; 5 - железница угревая (Demodex folliculorum), справа - железницы в волосяной сумке
Сосущие растительноядные клещи также связаны по происхождению с почвенными Prostigmata. Уже среди последних есть формы, высасывающие почвенные водоросли, соки мхов, но наиболее разнообразны заселившие высшие растения тетраниховые и галлообразующие клещи.
Тетраниховые клещи (надсем. Tetranychoidea) представлены несколькими семействами, среди которых наиболее распространены паутинные клещи (сем. Tetranychidae) и плоскотелки (сем. Tenuipalpidae, рис. 66, 67, табл. 9). Тело тетранихид овальное, цельное, у плоскотелок уплощено и разделено на протеро- и гистеросому. Это мелкие клещи 0,2-1 мм длиной. Кутикула тонкая, у плоскотелок уплотненная, зернистая, сетчатая. Характерна окраска различных оттенков зеленого, желтого, красного цвета. Щетинки обычно игольчатые, но бывают листовидные, перистые и др. Ротовые органы приспособлены для прокола эпидермиса и сосания сока. Основные членики хелицер сросшиеся, в них помещаются два выдвижных длинных стилета - измененные концевые членики хелицер. Внутри коротких, сросшихся в основании педипальп у паутинных клещей помещаются паутинные железы, протоки которых открываются на концевом членике. Близ хелицер открываются стигмы трахей. Ноги тонкие, на лапках имеется сложный коготковый аппарат, с помощью которого клещи удерживаются на листьях: средний коготок (эмподий), иногда с придатками, и пара боковых, с двумя булавчатыми волосками каждый. Наружный половой аппарат упрощен, нет ни клапанов, ни половых присосок. У самок половое отверстие прикрыто кожной складкой, у самцов имеется копулятивный орган на заднем конце тела.
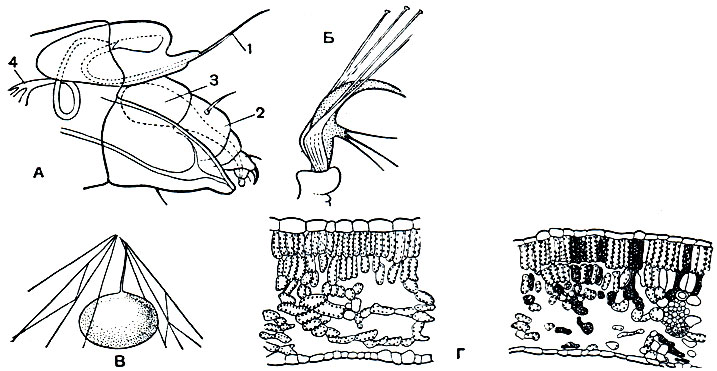
Рис. 66. Паутинные клещи: А - ротовой аппарат паутинного клеща; Б - ноготковый аппарат лапки; В - яйцо; Г - поперечные разрезы здорового и поврежденного листа; 1 - колющие стилеты хелицер; 2 - педипальпы; 3 - паутинная железа; 4 - трахеи
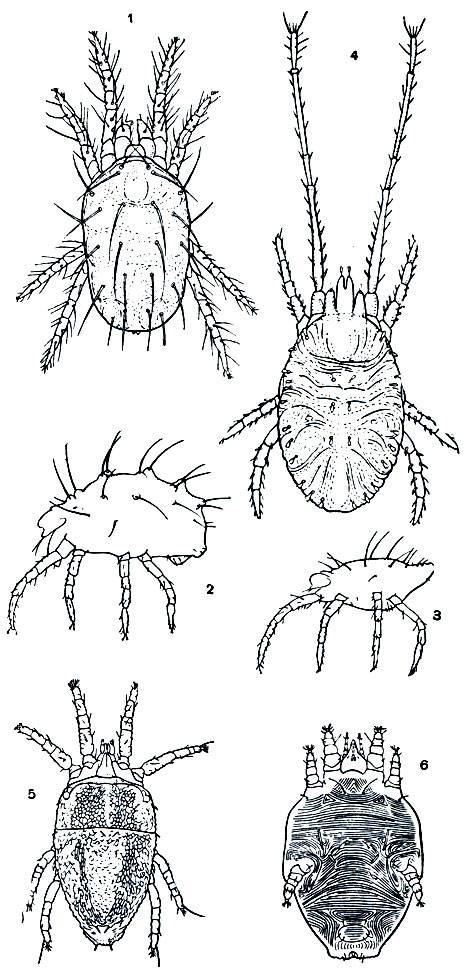
Рис. 67. Паутинные и плоские клещи: 1 - обыкновенный паутинный клещ, самка; 2, 3 - красный плодовый клещ (Panonychus ulmi), самка и самец, вид сбоку; 4 - Bryonia redikorzevi; 5 - плоский клещ Brevipalpus obovatus; 6 - шестиногий клещ Phytoptipalpus paradoxus

Таблица 9. Растительноядные и водяные клещи: 1-7 - клещи и вызываемые ими повреждения: 1 - лист хлопчатника, поврежденный обыкновенным паутинным клещом, 2 - колония обыкновенного паутинного клеща на листе огурца, самки (красная - зимовочная), самец, яйца, личинки, нимфы, 3 - скопления зимовочных самок обыкновенного паутинного клеща, спускающиеся на паутине с листьев огурца, справа - скопления самок при большем увеличении, 4 - сосочковидные галлы ольхового клеща (Eriophyes laevis) на листе ольхи, 5 - бляшковидные и краевые галлы грушевого клеща (E. piri) на листьях груши, в кругу сильно увеличенный грушевый клещ, 6 - разрастание побегов ясеня, вызванное ясеневым галловым клещом (E. fraxinivorus), 7 - осмоленные галлы соснового клеща (E. pini) на побегах сосны; 8-10 - водяные клещи: 8 - Axonopsis complanata, 9 - Piona coccinea, 10 - Brachypoda versicolor

Таблица 9. Растительноядные и водяные клещи: 1-7 - клещи и вызываемые ими повреждения: 1 - лист хлопчатника, поврежденный обыкновенным паутинным клещом, 2 - колония обыкновенного паутинного клеща на листе огурца, самки (красная - зимовочная), самец, яйца, личинки, нимфы, 3 - скопления зимовочных самок обыкновенного паутинного клеща, спускающиеся на паутине с листьев огурца, справа - скопления самок при большем увеличении, 4 - сосочковидные галлы ольхового клеща (Eriophyes laevis) на листе ольхи, 5 - бляшковидные и краевые галлы грушевого клеща (E. piri) на листьях груши, в кругу сильно увеличенный грушевый клещ, 6 - разрастание побегов ясеня, вызванное ясеневым галловым клещом (E. fraxinivorus), 7 - осмоленные галлы соснового клеща (E. pini) на побегах сосны; 8-10 - водяные клещи: 8 - Axonopsis complanata, 9 - Piona coccinea, 10 - Brachypoda versicolor
Оплодотворение внутреннее, без сперматофоров, в ряде случаев известно девственное развитие. Яйца округлые, иногда с нитевидным придатком, приклеиваются к поверхности листьев, зимующие яйца - в трещинах коры, в опавшей листве. Цикл развития (рис. 56, 6) включает сходные по образу жизни личинку, две нимфы и взрослую фазу, по облику подобную нимфам (неотеническую). При образовании самца у некоторых видов нимфа только одна. Каждой линьке предшествует более или менее длительный покой. Периодичность размножения различна. Многие тропические виды, а также оранжерейные в умеренном климате размножаются непрерывно, давая до 20 поколений в год. Виды умеренных широт размножаются в теплое время, одни успевают проделать несколько генераций, другие только одну. Зимуют либо яйца, либо оплодотворенные самки, обычно в состоянии диапаузы.
Круг кормовых растений тетраниховых клещей в целом широк. Паутинные клещи живут главным образом на лиственных деревьях, но немало видов обитает на травянистых растениях, в частности клещи-бриобии (подсем. Bryobiinae) в основном живут на травах. На хвойных встречаются немногие виды. Клещи-плоскотелки также в основном обитатели листопадных древесных растений, но немало обитателей хвойных, а также злаков. Большинство видов имеет более или менее ограниченный набор кормовых растений (олигофаги), некоторые живут только на одном виде растений (монофаги), но есть и такие, которые могут жить и размножаться на сотнях видов как древесных, так и травянистых растений (полифаги).
Большинство видов семейства Tetranychidae в том или ином количестве выделяет паутину, которой покрываются листья и другие части растений. Паутинный покров служит защитой клещам и особенно характерен для видов, живущих большими колониями. На паутинках клещи также расселяются, подхваченные токами воздуха.
Все виды в той или иной мере повреждают растения. Питаясь, клещи наносят многочисленные ранки, через которые высасывают содержимое клеток. Разрушаются хлоропласты, клетки ассимиляционной паренхимы буреют и сморщиваются. Внешне повреждения выглядят различно. Листья желтеют, становятся пятнистыми, светлые или бурые пятна сливаются, листья засыхают, иногда опадают. В других случаях листья краснеют, приобретают бронзовый или мраморный вид, иногда сильно деформируются. При массовом размножении клещей растения нередко гибнут. Особенно страдают хлопчатник, плодовые, бахчевые, огородные культуры, некоторые декоративные растения в открытом грунте и в оранжереях. Помимо непосредственного вреда, клещи переносят вирусные болезни растений.
До недавнего времени фауна тетранихид СССР была почти не изучена, главное внимание сельскохозяйственных энтомологов было направлено на борьбу с несколькими наиболее вредоносными видами, которые точно не диагностировались. В последние 10-15 лет этот пробел восполнен, в фауне СССР выявлено уже более 100 видов тетранихид, изучена биология ряда форм, опубликованы определители; Г. Ф. Рекк, Определитель тетраниховых клещей, Тбилиси, 1959; Б. А. Вайнштейн, Тетраниховые клещи Казахстана, Алма-Ата, 1960, и др.
Наиболее вредоносен обыкновенный паутинный клещ (табл. 9), которому разные специалисты до сих пор придают разные латинские названия из ранее употреблявшихся: Tetranychus telarius, T. urticae, T. turkestani и др. Этот вид очень широко распространен, в СССР наиболее обилен в южных районах, обычен в средней полосе, где в годы с жарким летом нередко размножается в массе, в оранжереях и на комнатных растениях встречается вплоть до Заполярья. Крайне многояден, известно несколько сотен видов кормовых растений, травянистых и древесных, кроме хвойных. Питающиеся клещи зеленые и желтоватые, живут колониями на нижней поверхности листьев, покрывая ее слоем паутины. Самка в течение двух-трех недель жизни при благоприятных условиях откладывает свыше 100 яиц. Длительность генерации - две-три недели, поколения накладываются во времени. В размножающейся колонии клещей можно найти все фазы - яйца, личинок, нимф, самок и самцов. Зимует оплодотворенная самка в состоянии диапаузы, которая наступает под влиянием сокращения длины светового дня. Во второй половине лета, когда условия еще подходящие для размножения клещей, в колониях начинают появляться готовящиеся к зимовке самки. Они перестают питаться, приобретают оранжево-красный цвет и уходят с растения, выделяя по пути много паутины. Зимуют они в растительных остатках, под комочками почвы, собираясь в плотные скопления.
Нередко самки еще на растении собираются в клубочки, которые свешиваются на паутинных тяжах. В южных районах и в оранжереях клещи размножаются непрерывно, давая в год до 20 поколений. Потенциальная размножаемость колоссальна. Приводят несложный подсчет: если бы в потомстве каждой самки выживало 50 особей, то при 15 поколениях в год от одной исходной самки получилось 6⋅1035 особей, которые покрыли бы земной шар сплошным слоем в 2,5 м. Поэтому вспышки массового размножения возникают внезапно при очень низкой первоначальной численности клещей.
Обыкновенный паутинный клещ на юге сильно вредит хлопчатнику, плодовым, бахчевым культурам, в более северных районах - оранжерейным и парниковым растениям, в особенности огурцам. Значителен экономический ущерб в хлопководстве. Листья хлопчатника покрываются багрово-красными сливающимися пятнами и опадают. Отмирают бутоны, урожай коробочек снижается на 30-50%. Характерную картину повреждения хлопчатника в Средней Азии называют "ургамчак", "канасы", в Закавказье - "чор".
Яблоням, сливам и другим плодовым вредит красный плодовый клещ (Panonychus ulmi), до 0,45 мм, темно-красный, спинные щетинки крупные, сидят на бугорках. Этот вид распространен в южной половине Европейской части СССР, в Крыму, на Кавказе. Зимуют яйца в трещинах коры, личинки появляются с распусканием листьев, на юге бывает несколько поколений в сезон. Помимо плодовых, этот клещ живет на различных лиственных породах, в особенности из розоцветных.
Меры защиты растений от клещей различны в зависимости от культуры, агротехники, вида вредителя и т. д. Химическая борьба не всегда приносит желаемый результат. Обработки садов ДДТ не раз приводили к последующим вспышкам размножения клещей, так как яд в большей степени действует на хищников, хищных клещей, клопов, личинок двукрылых и др., подавляющих численность паутинных клещей. На хлопчатнике хорошие результаты дает применение препаратов системного действия, делающих сок растений ядовитым для клещей.
Галлообракующие, или четырехногие, клещи (сем. Eriophyidae, надсем. Eriophyoidea, или Tetrapodili) - крайне специализированные паразиты растений (рис. 68). Эти мельчайшие существа (0,1-0,2 мм) высасывают содержимое растительных клеток и вызывают у растений появление тех или иных уродливых образований, наростов, уродливости роста побегов, цветков и плодов, ржавых пятен, волосистых подушек (эринеумов), разнообразных галлов (табл. 9). Часть видов живет на поверхности растений (подсем. Phyllocoptinae), большинство же - в галлах (подсем. Eriophyinae и Phytoptinae).
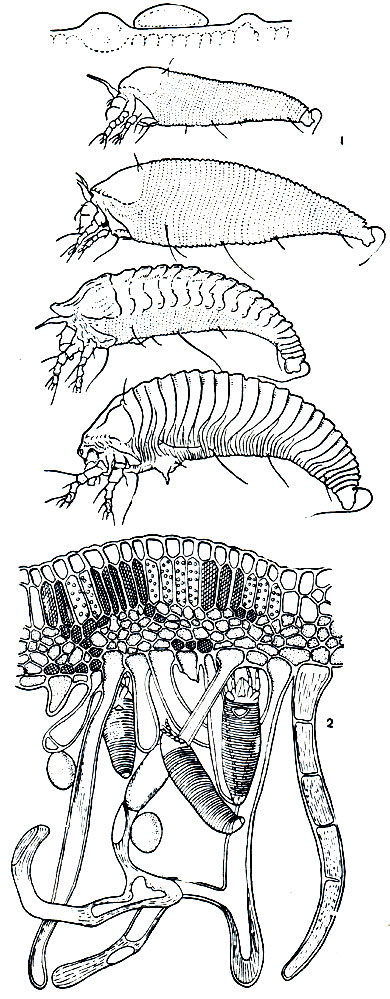
Рис. 68. Галлообразующие клещи: 1 - развитие четырехногого клеща Oxypleurites aesculifoliae; сверху вниз - яйцо, неполовозрелые фазы, называемые личинкой и протонимфой, летняя (протогинная) самка, зимующая (дейтогинная) самка; 2 - виноградный галловый клещ (Eriophyes vitis) среди разросшихся волосков листа
Тело эриофиид удлиненное, у галловых форм червеобразное, с мягким тонкокольчатым покровом, у открытоживущих более плоское, чешуйчатое. Ног только две передние пары, хелицеры колющие, глаз и трахей нет, половое отверстие сдвинуто вперед и прикрыто клапаном. Яйца очень крупные относительно самок. В послезародышевом развитии сохранилось только две линьки (рис. 56, 5), причем все фазы внешне очень сходны. Размножение в раннем возрасте (неотения), характерное для простигматических клещей, в частности тетранихид, доведено здесь до крайней степени. Строение фаз развития эриофиид таково, что их нельзя назвать ни личинками, ни нимфами в том смысле, как у других клещей. Перед нами, по существу, преждевременно вылупившиеся эмбрионы, у которых нет даже двух задних пар ног, одевшиеся кутикулой и способные размножаться. Структуры и процессы, свойственные послезародышевому развитию исходных форм, здесь в значительной степени утрачены, сохранилось только то, что необходимо для скрытой, эндопаразитической жизни при неограниченном источнике пищи. Это явление сверхраннего вылупления, называемое дезэмбрионизацией, встречается и у других эндопаразитов, живущих в живом пищевом субстрате, например у удивительных личинок наездников-яйцеедов, которые развиваются в чужом желтке. Только там личинка в дальнейшем окукливается и вылетает нормальное насекомое. Здесь же и размножение происходит в состоянии полуэмбриона, так как все развитие проходит в галлах и оборвано благодаря неотении. По происхождению эриофииды связаны с тетраниховыми клещами. Связующим звеном служат похожие на клещей-плоскотелок шестиногие клещи (сем. Phytoptipalpidae), живущие на растениях и размножающиеся в личиночной фазе. Эволюционный ряд от почвенных Prostigmata к тетранихидам, шестиногим и галловым клещам иллюстрируют рисунки 67, 68. Обращает на себя внимание внешнее сходство галловых клещей с железницами. Обе группы - конечные этапы паразитической специализации разных ветвей Prostigmata, одни эндопаразиты животных, другие - растений.
Жизненные циклы эриофиид строго согласованы с сезонными явлениями в жизни растений и в деталях разнообразны. Для ряда видов характерно образование двух типов самок - летних (протогинных), которые живут и размножаются в галлах, и зимних (дейтогинных), которые появляются к осени, уходят из галлов и зимуют в почках, трещинах коры, стерне и т. п. Весной они переходят на зеленые части растений и служат основательницами колоний клещей во вновь образуемых галлах.
Насчитывается несколько сотен видов эриофиид. Каждый вид, как правило, живет на одном или немногих видах растений-хозяев и вызывает характерные повреждения. Некоторые виды образуют по нескольку вариететов соответственно видам заселяемых растений, причем варьирует и характер повреждений. Наиболее многочисленны галловые клещи древесных пород. В справочнике "Вредители леса" (изд. АН СССР, 1955) только для Палеарктики указано 127 видов и экологических форм. Одни из них вызывают образование на листьях подушечек и в давлений, покрытых белесым войлочком из густых волосков, или загибание краев листовой пластинки, другие - различные бляшковидные, сосочковидные и иные галлы, третьи - уродливое разрастание почек, побегов и т. п. Почти каждая древесная порода имеет свой характерный набор галловых клещей. Особенно заметны красные или желтоватые галлы в виде небольших вытянутых сосочков. На листьях ольхи их вызывает ольховый галловый клещ (Eriophyes laevis), на листьях липы - липовый (E. tilae), на листьях черемухи - черемуховый (E. padi), на других породах - другие виды. Некоторые виды галловых клещей вызывают на деревьях образования, известные под названием "ведьминых метел", например E. triradiatus на ивах. Клещи живут в почках и под влиянием их сосания побеги вырастают укороченными, но появление новых побегов из пазух листьев ускоряется. В результате в короткий срок вырастает столько побегов, сколько при нормальных условиях может образоваться только в течение нескольких лет. Масса уродливых перепутанных побегов образует подобие метлы.
Многие виды галловых клещей вредят культурным растениям. Виноградный войлочный клещ (Eriophyes vitis) вызывает на нижней стороне листьев винограда волосистые пятна, сначала белые, затем буреющие. Вред ощутим в засушливые периоды. Грушевый клещ (E. piri) вызывает образование бляшковидных выпуклостей на листьях груши, яблони и других плодовых. Смородинный почковый клещ (E. ribis) вызывает вздутие почек смородины. Клещ Aceria tulipae живет под чешуйками луковиц лука, чеснока, тюльпана, вызывая сморщивание и высыхание луковиц. Галловые клещи переносят вирусы растений; сейчас их интенсивно изучают в этом отношении.
Клещи-краснотелки (надсем. Trombea) - формы со сложным метаморфозом, который возник в результате того, что одни фазы развития взяли на себя функцию питания, а другие за этот счет перестали питаться и превратились в формообразовательные покоящиеся фазы, подобные куколке насекомых (рис. 69). Но в отличие от последних, у которых куколка одна, у краснотелок фазы питания и формообразования чередуются (рис. 56,10). Из яиц, откладываемых в почву, вылупляются неподвижные предличинки, которые превращаются в личинок, потребляя запас желтка в кишечнике. Личинки паразитируют на насекомых или позвоночных. Они подстерегают животных в открытой природе, обычно на растительности (так называемый пастбищный паразитизм), присасываются к ним надолго, высасывают относительно большое количество гемолимфы или лимфы, сильно увеличиваются в размерах и отпадают. Дальнейшее превращение происходит уже вне связи с животными-хозяевами. Запасшаяся пищей личинка превращается в покоящуюся протонимфу, за которой следует активная дейтонимфа, непитающаяся тритонимфа и активная взрослая фаза. Дейтонимфы и взрослые краснотелки-хищники, высасывают мелких членистоногих, нередко их яйца, и также весьма прожорливы. Избыточное питание активных фаз, в первую очередь личиночный паразитизм, было той основой, на которой возник метаморфоз краснотелок. Личинки перешли к паразитизму от питания мелкими членистоногими (энтомофагии), начав присасываться к более крупным насекомым (клещи сем. Trombidiidae) и к позвоночным (сем. Trombiculidae).
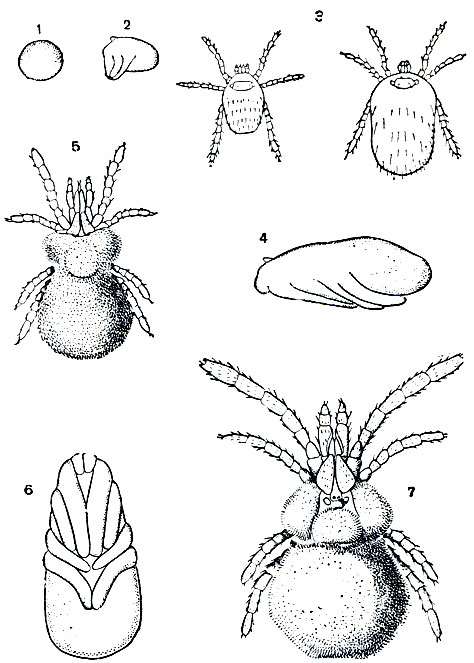
Рис. 69. Краснотелка Eutrombicula batatas: 1 - яйцо; 2 - предличинка; 3 - паразитическая личинка, голодная и напитавшаяся; 4 - покоящаяся протонимфа; 5 - хищная дейтонимфа; 6 - покоящаяся тритонимфа; 7 - хищная самка
Возникли и резкие различия в строении фаз развития. Неподвижные фазы имеют тонкие гладкие покровы, чехловидные придатки, нефункционирующие ротовые органы. На этих фазах происходит растворение (гистолиз) значительной части органов и тканей и из клеточных зачатков образуются новые органы и ткани. Паразитическая личинка имеет мешковидное тело, одетое эластичной растягивающейся кутикулой с редкими щетинками, прицепные коготки, ротовые органы в виде "головки", приспособленные для прокола п фиксации в покровах животного-хозяина. Дейтонимфа и особенно взрослые клещи гораздо крупнее голодных личинок (до 3-5 мм), туловище разделено на протеро- и гистеросому, густо опушено бархатистыми волосками и нередко ярко окрашено в различные оттенки красного цвета, откуда и пошло название краснотелок (табл. 8, 4). Ротовые органы обособлены, хелицеры с когтевидным пальцем, когтевидный вырост имеется и на педипальпах. Близ ротовых органов открываются дыхальца трахей. Хорошо развиты трихоботриии двойные боковые глаза на протеросоме. Крепкие бегательные ноги богато оснащены различными кожными органами чувств, особенно передние. Наружный половой аппарат сходен у обоих полов (половые крышки, выпячивающийся конус, три пары половых щупалец), что связано с сохранившимся у краснотелок сперматофорным оплодотворением без копуляции. Если отвлечься от личиночного паразитизма, сложного метаморфоза и укрупнения размеров краснотелок, то легко усмотреть их сходство с примитивными Endeostigmata, находящимися в истоках этого отряда клещей.
Ярко-красные крупные виды семейства Trombidiidae (род Trombidium и многие другие) нередко попадаются на влажной почве, а их мелкие личинки - присосавшимися к насекомым. Краснотелки семейства Trombiculidae мельче, их личинки часто в большом количестве встречаются на мелких млекопитающих и птицах, особенно на ушах грызунов. Личинки тромбикулид нападают на человека, вызывая раздражение кожи (тромбидиоз), особенно в теплых влажных районах.
Тромбикулиды передают человеку японскую речную лихорадку "цуцугамуси" - опасное заболевание, распространенное в Юго-Восточной Азии, на островах Тихого океана и недавно обнаруженное у нас на Дальнем Востоке. Болезнь вызывается особыми микроорганизмами - риккетсиями, которыми личинки заражаются от грызунов и насекомоядных. Личинка питается только раз, но риккетсии сохраняются в организме клеща в течение метаморфоза и передаются через яйцо (трансовариально) голодным личинкам следующего поколения, которые и заражают человека.
Водяные клещи представлены двумя неродственными группами: пресноводные клещи - гидрахнеллы (группа Hydrachnellae) и морские клещи - галакариды (Halacarae). Гидрахнеллы (рис. 70) обычны в пресных водоемах, особенно стоячих; их известно более 2000 видов, группируемых в 9 семейств и 3 надсемейства (по новейшей классификации Б. А. Вайнштейна)*. Это в большинстве хорошо плавающие формы, часть видов постоянно держится на водной растительности. Взрослые обычно 1-2 мм длиной, но немало и более крупных, до 3-5 мм и даже 8 мм. Это самые крупные представители отряда. Тело цельное, обычно округлое, покровы кожистые, плотные, нередко покрыты шипиками, сетчатой скульптурой или щитками различной величины и формы. Окраска разнообразна - ярко-красная, оранжевая, желтая, бурая, зеленая, синеватая, иногда пестрая (табл. 9). Обычно развита пара двойных глаз. Ноги снабжены плавательными волосками. Тазики ног крупные, сращены с телом в виде коксальных щитков. Ротовые органы объединены в "головку", хелицеры с когтевидным концевым члеником, когтевидный вырост на педипальпах иногда образует с их последним члеником род хватательной клешни. Близ ротовых органов открываются дыхальца хорошо развитых ветвящихся трахей. Половой конус прикрыт половыми створками, у самок выполняет роль яйцеклада, у самцов - органа, выделяющего сперматофоры; половые присоски варьируют в числе.
* (Прежде насчитывали до 20 семейств.)
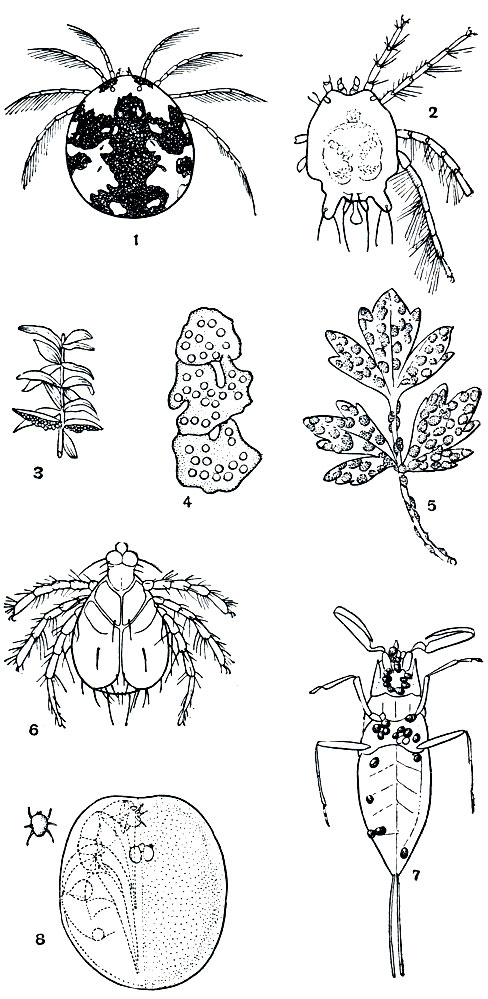
Рис. 70. Водяные клещи: 1 - Hydrarachna geographica; 2 - Arrhenurus neumani, самец; 3-5 - кладки яиц водяных клещей; 6 - паразитическая личинка Piona; 7 - личинки водяных клещей, присосавшиеся к клопу - водяному скорпиону; 8 - насосавшаяся личинка Eylais, под шкуркой которой образуется нимфа; слева показан относительный размер голодной личинки
Гидрахнеллы родственны краснотелкам, о чем свидетельствует их строение и особенно метаморфоз. Сперматофорное оплодотворение осложнено различными способами спаривания, при котором самец переносит слизистый сперматофор . с помощью третьей пары ног, подтягивает самку к сперматофору, прикрепленному к субстрату, и т. п. Гидрахнеллы кладут яйца на поверхность подводных предметов. Плодовитость самки может достигать нескольких сотен яиц. Яйца окружены набухающим секретом, нередко тысячи кладок сливаются в сплошной ослизненный слой. Предличиночная фаза проходит в яйце. Личинки гидрахнелл - паразиты, главным образом водных насекомых, по облику напоминают личинок краснотелок. Они либо отыскивают насекомых, плавая в толще воды, либо всплывают и подстерегают насекомых, плавающих по поверхности и вылетающих из воды. Насекомые-хозяева разнообразны: водные жуки, клопы, стрекозы, поденки, веснянки, ручейники, различные двукрылые. Клещи рода Unionicola не только в личиночной фазе, но и во всех последующих паразитируют в мантийной полости и на жабрах пластинчатожаберных моллюсков - беззубок и др. Личинки присасываются к насекомым надолго, как бы срастаясь ротовыми органами с покровом, сосут гемолимфу и постепенно очень сильно увеличиваются, по весу в сотни раз. Паразитизм, помимо питания, имеет расселительное значение, личинки заносятся насекомыми в новые водоемы и отпадают в воду. Дальнейшее развитие представляет гистолитический метаморфоз, как у краснотелок. Однако покоящиеся фазы - прото- и тритонимфа протекают здесь под покровом предшествующих. Линечные шкурки сбрасываются по две. Под покровом личинки образуется кутикула протонимфы (аподерма), под ней кутикула дейтонимфы, и уже затем шкурки личинки и протонимфы сразу сбрасываются (рис. 56, 11). То же происходит при превращении дейтонимфы во взрослого клеща. Поэтому внешне линек только две. В периоды метаморфоза клещ прицепляется к подводному предмету и переходит в покоящееся состояние - окукливается.
Дейтонимфы и взрослые клещи-хищники, питаются мелкими рачками, личинками водных насекомых. Взрослую фазу гидрахнелл нередко называют имагинальной. Действительно, плотный покров, развитые трахеи, укрупнение размеров, сложные формы спаривания, долговечность (многие виды живут более года), способность активно расселяться - все это признаки имаго, сходные с теми, которые мы видели у орибатид, с тем различием, что здесь имаго водное и способ его формирования иной. Имагинальность гидрахнелл выступает особенно отчетливо, если сравнить взрослую фазу этих клещей с тем, что мы видели у примитивных Acariformes и в особенности у неотенических представителей отряда.
В водоемах разных типов видовой состав гидрахнелл различен. Наиболее обильны и разнообразны они среди зарослей водной растительности в прудах и озерах, где обычны многие виды родов Eylais, Hydrachna, Piona, Arrhenurus и др. Беднее фауна рек, ручьев, временных водоемов. Специфичные виды найдены в подземных водах и горячих источниках.
Галакариды (рис. 71) в большинстве морские формы, но есть и пресноводные. По происхождению они связаны с Prostigmata. Это мелкие клещи (0,2-1,7 мм). На спине обычно четыре щитка, на брюшной стороне - два и по бокам сросшиеся пластинчатые тазики ног. Ноги не приспособлены для плавания, плоско расставлены, клещи держатся на поверхности подводных предметов. Ротовые органы в виде "головки", хелицеры колющие. Половой конус самки превращен в яйцеклад, у самца - в копулятивный орган. Яйца откладывают на подводные предметы, иногда носят на задних ногах. Развитие - как у большинства Prostigmata: яйцо, личинка, прото-, дейтонимфа, размножающаяся фаза. Галакариды высасывают сок водорослей, разлагающиеся остатки животных, мелких беспозвоночных и их яйца. Паразитизм редок. Известны паразиты морских ежей, хитонов, речного рака. Галакариды широко распространены в морях, до полярных широт, живут главным образом в прибрежной зоне до 30-40 м, особенно на участках, богатых водорослями. Известно около 200 видов семейства Halacaridae.
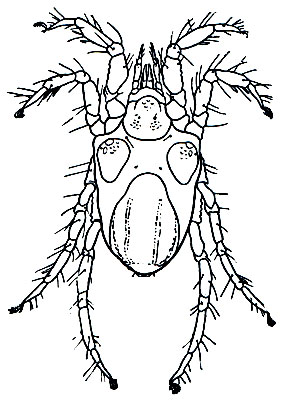
Рис. 71. Морской клещ Copidognathus fabricii
Данные о водяных клещах имеются в двух выпусках "Фауны СССР" (1940 и 1952), написанных И. И. Соколовым.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'