Отряд Веслоногие ракообразные (Copepoda)
При изучении жизни каждого водоема - от маленькой лужи до океана - исследователь встречается с этими рачками. Планктонная сеть, протянутая на любой глубине океана, в жарких тропиках или в холодных приполярных водах, приносит улов, состоящий преимущественно из веслоногих рачков. В ручьях и озерах глубоких темных пещер также обитают эти ракообразные. Они массами плавают во всех обычных пресноводных водоемах, часто не уступая по численности ветвистоусым. Некоторые из них паразитируют на рыбах и других водных животных, причем нередко под влиянием паразитизма меняются настолько, что не сразу можно угадать их истинную природу. Веслоногие массами поедаются рыбами. Сельдь, иваси и многие другие рыбы питаются почти исключительно веслоногими. Большинство этих ракообразных - животные микроскопические, длина их тела от 1 мм и меньше до 5 мм; лишь отдельные глубоководные и паразитические виды достигают в длину 1-3 см.
Тело свободноживущих веслоногих разделяется на головогрудь, грудь и брюшко (рис. 202). Голова слитная, без всяких следов сегментации, срастается с первым грудным сегментом, образуя головогрудь. Передний конец головы часто вытянут в загнутый вниз клюв, или рострум. Очень характерно отсутствие парных фасеточных глаз; на лобной части головы расположен только науплиальный глазок. Именно это обстоятельство позволило датскому натуралисту Мюллеру в свое время назвать обычных пресноводных веслоногих "циклопами" в честь одноглазого гиганта греческой мифологии.
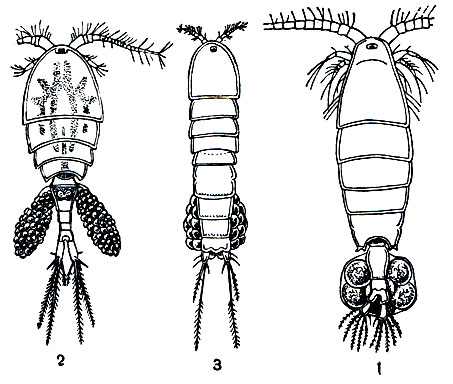
Рис. 202. Общий вид самок трех подотрядов веслоногих: 1 - Calanoida; 2 - Cyclopoida; 3 - Harpacticoida
Голова снабжена 5 парами придатков. Передние антенны часто очень длинные, иногда длиннее тела, и участвуют в плавании и парении рачков. Кроме того, они выполняют и функции органов чувств: на них сидят чувствительные щетинки и цилиндрические чувствительные придатки. Задние антенны короткие, обычно двуветвистые. Жвалы мощные и имеют двуветвистый щупик. Их жевательная сильно хитинизированная часть обладает острыми зубцами, помогающими раздроблять пищу. При внимательном рассмотрении зубцов жвал некоторых морских веслоногих выяснилось, что эти зубцы покрыты кремневыми коронками, увеличивающими их прочность (рис. 203). Открытие кремневых коронок интересно в двух отношениях. Во-первых, оно указывает на способность веслоногих ракообразных усваивать и концентрировать кремнии; такой способности лишены почти все высшие беспозвоночные - черви, моллюски, а также другие членистоногие.
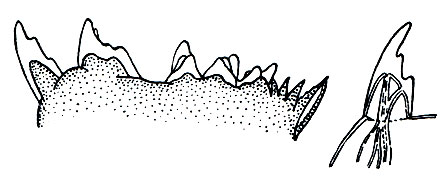
Рис. 203. Край жующей пластинки Eucalanus bungii и один зубец при большом увеличении
Во-вторых, можно надеяться найти в геологических отложениях кремневые коронки древних веслоногих, почти совсем не сохранившихся в ископаемом состоянии.
Передние челюсти веслоногих устроены очень сложно, так как снабжены внутренними и наружными лопастями и многочисленными перистыми щетинками. Задние челюсти имеют только внутренние лопасти и также многочисленные щетинки. К головным придаткам присоединяется пара одноветвистых ногочелюстей, принадлежащих переднему слившемуся с головой грудному сегменту.
Задние антенны, щупики жвал и передние челюсти фильтрующих веслоногих совершают частые и беспрерывные взмахи, создающие круговороты воды, которые приносят взвешенные пищевые частицы. Эти частицы отфильтровываются главным образом щетинками задних челюстей.
Грудной отдел состоит из 5 сегментов с ясно видимыми границами между ними. Все 5 пар грудных ножек у примитивных веслоногих устроены одинаково. Каждая ножка состоит из 2-члениковой основной части и двух обычно 3-члениковых ветвей, вооруженных шипами и щетинками. Эти ножки совершают одновременные взмахи, действуя как весла и отталкивая тело рачка от воды. У многих более специализированных видов пятая пара ножек самца преобразована в аппарат, приспособленный для удерживания самки во время спаривания и прикрепления сперматофоров к ее половым отверстиям. Нередко пятая пара ног редуцирована.
Брюшной отдел состоит из 4 сегментов, но у самок часто их число меньше, так как некоторые из них сливаются между собой. На переднем брюшном сегменте открывается парное или непарное половое отверстие, и у самки этот сегмент часто крупнее остальных. Брюшко заканчивается тельсоном, с которым сочленены фуркальные ветви. Каждая из них вооружена несколькими очень длинными, иногда перистыми щетинками. Эти щетинки особенно сильно развиты у планктонных видов, у которых они приспособлены для парения в воде, так как препятствуют погружению рачка.
Строение паразитических видов сильно отклоняется от описанного. Их головные придатки превращаются в органы прикрепления к хозяину и в связи с этим часто имеют вид хитиновых крючков, погруженных в кожу рыб или других водных животных. Плавательные ножки иногда исчезают совсем, или число их сокращается. В большей или меньшей степени исчезает сегментация, покровы становятся мягкими. Специализированные паразиты приобретают червеобразную форму тела и теряют всякое сходство с ракообразными. Однако можно подобрать полную серию переходов между такими предельно упрощенными видами и нормальными свободноживущими. Кроме того, яйца прикрепляются к телу паразитических видов совершенно так же, как к телу свободноживущих, и из яиц выходят такие же личинки.
Дыхание веслоногих осуществляется всей поверхностью тела, жабры отсутствуют. С этим, возможно, связано и слабое развитие или даже отсутствие кровеносной системы. Сердце есть только у представителей подотряда Calanoida, да и у них оно невелико, хотя и бьется очень часто: например, у морского рачка Labidocera оно совершает более 150 ударов в минуту. У остальных веслоногих полостная жидкость приводится в движение сокращениями кишечника.
Все веслоногие раздельнополы. Самцы обычно меньше самок и у свободноживущих видов отличаются от самок искривленными (одной или обеими) передними антеннами, а также иным строением пятой пары грудных ножек. Особенно резко различны самцы и самки многих паразитических видов. У некоторых из них прикрепленные к хозяину самки полностью теряют сегментацию и конечности, а маленькие самцы сохраняют облик поздней личинки и свободно плавают. У других (семейство Lernaeopodidae) маленькие самцы, также сходные с поздней личинкой, своими клешневидными челюстями и ногочелюстями прикрепляются к крупным червеобразным паразитическим самкам.
При спаривании самец удерживает самку пятой парой грудных ножек и первыми антеннами и при помощи той же пятой пары ног приклеивает колбасовидный сперматофор близ ее половых отверстий, т. е. к нижней стороне первого брюшного сегмента. У некоторых видов одна из ветвей пятой пары ног самца снабжена на конце щипчиками, захватывающими сперматофор и переносящими его на нужное место (рис. 204). Из сперматофора сперма попадает в семеприемник самки. При выметывании яиц они оплодотворяются.
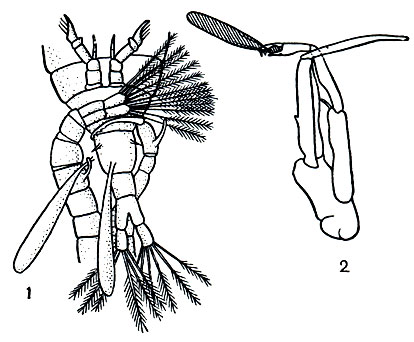
Рис. 204. Спаривание Calanoida: 1 - прикрепление сперматофора к генитальному сегменту самки у Diaptomus; 2 - пятая пара ног Pareuchaeta glacialis; последний членик левой ноги с 'щипчиками', удерживающими сперматофор
Большинство морских планктонных видов откладывает яйца прямо в воду, но у всех пресноводных и у всех паразитических, а также у морских донных и прибрежных видов яйца склеиваются между собой специальным секретом и прикрепляются близ половых отверстий самки. У одних веслоногих таким образом формируется один, у других - два яйцевых мешка, которые самка носит на себе до выхода из яиц личинок.
Из яйца выходит личинка науплиус. Личинка многократно линяет и постепенно приближается по своим признакам к взрослому рачку. Различают 12 личиночных стадий веслоногих. Первые две стадии - ортонауплиуса - характеризуются присутствием только обеих пар антенн и пары жвал, у следующих четырех стадий - метанауплиуса - закладываются и развиваются остальные ротовые придатки, но тело остается несегментированным. Последние 6 стадий называются копеподитными и отличаются сегментацией заднего конца тела и постепенным развитием грудных ножек. Для завершения метаморфоза разным веслоногим требуется различное время, да и биология личинок далеко не у всех видов одинакова.
Образ жизни, способ питания и среда обитания веслоногих рачков настолько разнообразны, что лучше рассмотреть этот отряд не целиком, а каждый из входящих в него подотрядов в отдельности.
В пределах отряда веслоногих принято различать 5 подотрядов, из которых 2 объединяют исключительно паразитические виды*. Свободноживущие веслоногие относятся к подотрядам Calanoida, Cyclopoida и Harpacticoida (рис. 202).
* (Некоторые исследователи разделяют паразитических веслоногих на 5 или 6 подотрядов. )
Calanoida - исключительно планктонные животные. Их голова и грудь значительно длиннее узкого брюшка, передние антенны очень длинные, превосходят голову и грудь, а нередко и все тело рачка, если есть яйцевой мешок, то один.
Cyclopoida обитают преимущественно на дне или в придонных слоях воды. Среди них есть и паразиты. Их голова и грудь относительно всего тела гораздо короче, чем у Calanoida, передние антенны короткие, не достигают и середины головогрудного отдела, брюшко уже груди. Самки всегда носят два яйцевых мешка.
Harpacticoida, за единичными исключениями, живут на дне и больше ползают, чем плавают. Тело их червеобразно благодаря тому, что брюшной отдел почти не отличается по ширине от грудного. Передние антенны очень короткие, самки большинства видов образуют один яйцевой мешок. Представители всех трех подотрядов населяют как моря, так и пресные воды.
Подотряд Каланиды (Calanoida)
Вся организация Calanoida превосходно приспособлена к жизни в толще воды. Длинные антенны и перистые щетинки фуркальных ветвей позволяют морскому Calanus или пресноводному Diaptomus неподвижно парить в воде, лишь очень медленно погружаясь. Этому способствуют находящиеся в полости тела рачков капли жира, уменьшающие их удельный вес. Во время парения тело рачка располагается вертикально или наклонно, причем передний конец тела расположен выше заднего. Опустившись на несколько сантиметров вниз, рачок делает резкий взмах всеми грудными ножками и брюшком и возвращается на прежний уровень, после чего все повторяется сначала. Таким образом, путь рачка в воде рисуется зигзагообразной линией (рис. 205, 1). Некоторые морские Calanoida, как например приповерхностный вид ярко-синего цвета Pontellina mediterranea, совершают настолько резкие скачки, что выпрыгивают из воды и пролетают какое-то пространство по воздуху наподобие летучих рыб.
Если грудные ножки действуют время от времени, то задние антенны, щупики жвал и передние челюсти вибрируют беспрерывно с очень большой частотой, совершая до 600-1000 ударов ежеминутно. Их взмахи вызывают мощные круговороты воды с каждой стороны тела рачка (рис. 205, 2). Эти токи проходят через образованный щетинками челюстей фильтрационный аппарат, и отфильтрованные взвешенные частицы проталкиваются вперед к жвалам. Жвалы размельчают пищу, после чего она поступает в кишечник.
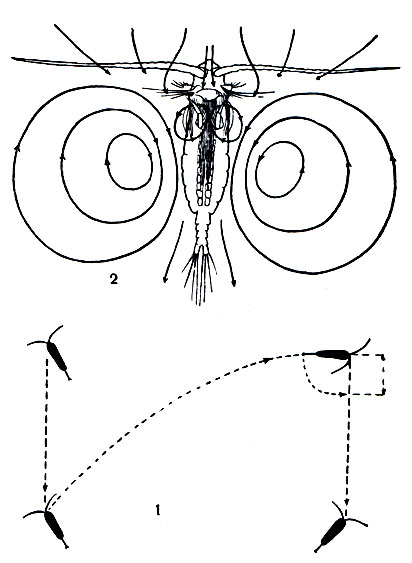
Рис. 205. Движения Calanoida: 1 - движение Diaptomus в воде; 2 - токи воды, образуемые конечностями калануса
Пищей фильтрующим Calanoida служат все организмы и их остатки, взвешенные в воде. Рачки не заглатывают только сравнительно крупные частицы, отталкивая их ногочелюстями. Основой питания Calanoida следует считать планктонные водоросли, потребляемые рачками в огромном количестве. Eurytemora hirundoides в период массового развития водоросли Nitzschia closterium поедала до 120 000 особей этих диатомовых в день, причем вес пищи почти достигал половины веса рачка. В случаях такого избыточного питания рачки не успевают усвоить все органическое вещество пищи, но все-таки продолжают ее заглатывать.
Для определения интенсивности фильтрации Calanus применялись водоросли, меченные радиоактивными изотопами углерода и фосфора. Оказалось, что один рачок за сутки пропускает через свой фильтрационный аппарат до 40 и даже до 70 см3 воды, причем питается он преимущественно ночью.
Питание водорослями необходимо для многих Calanoida. Так, например, половые продукты Calanus finmarchicus созревают только при достаточном потреблении рачком диатомовых водорослей.
Помимо фильтраторов, среди Calanoida есть и хищные виды, большинство которых обитает на значительных или больших океанических глубинах, где планктонные водоросли не могут существовать из-за отсутствия света. Задние челюсти и ногочелюсти таких видов снабжены крепкими острыми шипами и приспособлены для схватывания жертв. Особенно интересны приспособления для добывания пищи у некоторых глубоководных видов. Винкстед наблюдал, как глубоководная Pareuchaeta неподвижно висит в воде, расставив в стороны свои удлиненные ногочелюсти, образующие нечто вроде капкана (рис. 206). Как только между ними оказывается жертва, ногочелюсти смыкаются, капкан захлопывается. При крайней разреженности организмов на больших океанических глубинах такой способ охоты оказывается наиболее целесообразным, так как затрата энергии на активные поиски жертв не окупается их поеданием.
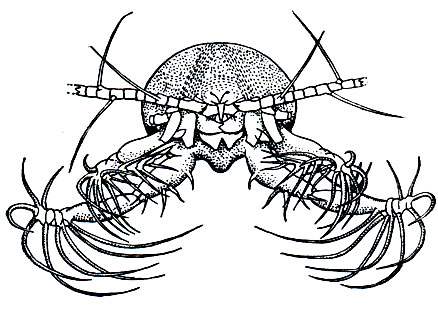
Рис. 206. Раскрытый 'ловчий капкан' Pareuchaeta
С особенностями движения и питания Calanoida связана сложная проблема их суточных вертикальных миграций. Уже давно было замечено, что во всех водоемах, как пресных, так и морских, огромные массы Calanoida (и многих других планктонных животных) ночью поднимаются ближе к поверхности воды, а днем опускаются вглубь. Размах этих суточных вертикальных миграций различен не только у разных видов, но даже у одного вида в разных местах его обитания, в разные сезоны года и у разных возрастных стадий одного вида. Нередко науплиусы и младшие копеподитные стадии держатся всегда в поверхностном слое, а старшие копеподитные стадии и взрослые рачки мигрируют. В северной части Атлантического океана диапазон вертикальных миграций Calanus finmarchicus составляет 300-500 м. Такие же громадные расстояния ежесуточно преодолевают дальневосточные Metridia pacifica и M. ochotensis. В то же время другие массовые дальневосточные Calanoida - Calanus plumchrus, C. cristatus, Eucalanus bungii - мигрируют не более чем на 50-100 м.
Скорость передвижений рачков при их вертикальных миграциях измеряется величинами порядка 10-30 см в минуту. Если принять во внимание длину их тела (для Calanus finmarchicus, например, около 2 мм), то такую скорость приходится считать значительной. При этом не только подъем кверху, но и опускание вниз осуществляется за счет активных движений рачков, а не за счет их пассивного погружения.
Не следует думать, что, совершая вертикальные миграции, все рачки одновременно двигаются в каком-нибудь определенном направлении. Английский ученый Бэйнбридж опускался под воду и проводил наблюдения над мигрирующими веслоногими.
Он видел, как в одном и том же слое воды часть рачков движется вверх, а другая - вниз. В зависимости от того, какое движение преобладает, перемещается вверх или вниз вся масса животных.
Вопрос о причинах вертикальных миграций до сих пор окончательно не выяснен. Совершенно очевидно, что стремление рачков подняться в поверхностные слои объясняется изобилием там планктонных водорослей, которыми питаются веслоногие-фильтраторы. Менее понятны причины, заставляющие рачков покидать эти богатые пищей слои. Многие исследователи считают, что свет вредно влияет на рачков и, избегая его, они утром начинают уходить вниз. Важное значение света подтверждается наблюдениями В. Г. Богорова над вертикальным распределением веслоногих в Баренцевом море летом, т. е. в условиях круглосуточного освещения. Оказалось, что в это время Calanus finmarchicus неизменно находится на одной глубине, там, где условия освещения для него наиболее благоприятны. В этом районе моря в толще воды наблюдаются внутренние волны, которые должны то несколько приподнимать, то несколько опускать рачков. Очевидно, рачки активно передвигаются в противоположном направлении, так как они в течение круглых суток не выходят за пределы определенного горизонта. Осенью, когда восстанавливается смена дня и ночи, возобновляются обычные вертикальные миграции (рис. 207). Не только солнечный, но и лунный свет заставляет рачков уходить из поверхностных слоев воды в более глубокие.
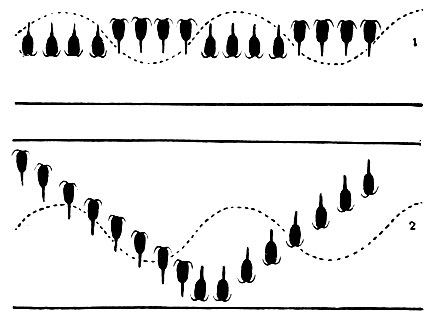
Рис. 207. Вертикальные миграции Calanus в Баренцевом море в условиях круглосуточного дня (1) и смены дня и ночи (2)
Однако далеко не во всех случаях вертикальные миграции удается связать непосредственно с действием света. Существуют наблюдения, показывающие, что рачки начинают опускаться вниз задолго до восхода солнца. Эстерли содержал веслоногих рачков Acartia tonsa и A. clausi в полной темноте, и, несмотря на это, они продолжали совершать регулярные вертикальные миграции.
По мнению некоторых ученых, уход рачков утром из освещенного слоя воды следует считать защитной реакцией, помогающей избегать выедания рыбами. Доказано, что рыбы видят каждого рачка, на которого они нападают. Опустившись в глубокие темные слои воды, рачки оказываются в безопасности, а в богатых водорослями поверхностных слоях ночью рыбы их также не могут рассмотреть. Эти представления не могут объяснить многие хорошо известные факты. Так, например, целый ряд веслоногих рачков совершает регулярные миграции небольшой протяженности, не выходя из освещенной зоны и оставаясь, следовательно, доступными для планктоноядных рыб.
Помимо суточных вертикальных миграций, морские веслоногие совершают и сезонные миграции. В Черном море летом температура поверхностного слоя повышается, и обитающий там Calanus helgolandicus опускается приблизительно на 50 м, а зимой снова возвращается на меньшую глубину. В Баренцевом море молодые стадии C. finmarchicus весной и летом держатся в поверхностных слоях. После того как они подрастут, осенью и зимой рачки опускаются вниз, и перед весной достигающие половозрелости особи начинают подниматься к поверхности, где и отрождается новое поколение. Особенно многочисленны в поверхностных слоях рачки, находящиеся на IV-V копеподитных стадиях и известные под названием "красного калануса", так как содержат большое количество жира буровато-красного цвета.
"Красный каланус" - излюбленная пища многих рыб, в частности сельди. Подобный характер сезонных миграций, т. е. подъем в поверхностные слои воды для размножения, обнаружен у многих других массовых видов, например у Calanus glacialis, C. helgolandicus, Eucalanus bungii и т. д. Самки этих видов нуждаются для развития половых продуктов в обильном питании водорослями, а возможно, и в освещении. Другие виды (Calanus cristatus, C. hyperboreus), наоборот, размножаются в глубоких слоях, а к поверхности поднимается только их молодь. Взрослые рачки С. cristatus совсем не питаются; у половозрелых особей даже редуцируются жвалы. Протяженность сезонных миграций обычно бывает больше, чем суточных. Первые иногда захватывают 3-4 тысячи метров, а вторые - самое большее несколько сотен метров.
Представители подотряда Calanoida преимущественно морские животные. В настоящее время известно около 1200 морских видов этих рачков, принадлежащих к 150 родам и 26 семействам. В пресных водах обитает всего около 420 видов, распределяющихся между 12 родами и 4 семействами.
Проведенные в последнее время подробные исследования фауны морских каланид показали, что прежние представления о широком распространении многих видов этих ракообразных неправильны. В каждой части океана обитают в основном присущие только ей виды. Каждый вид морских каланид расселяется благодаря несущим рачков течениям. Так, например, поступающие в Полярный бассейн ответвления Гольфстрима заносят туда каланид из Атлантического океана. В северо-западной части Тихого океана в водах теплого течения Куросио обитают одни виды, а в водах холодного течения Оясио - другие. Нередко по фауне каланид удается определить происхождение тех или иных вод в определенных частях океана. Особенно резко различаются по своему составу фауны вод умеренных широт и вод тропиков, причем тропическая фауна богаче видами.
Каланиды обитают на всех океанических глубинах. Среди них ясно различаются поверхностные виды и виды глубоководные, никогда не поднимающиеся в поверхностные слои воды. Как уже указывалось, на больших глубинах преобладают хищники, а на малых - фильтраторы. Наконец, существуют виды, совершающие вертикальные миграции огромного диапазона, то поднимающиеся к поверхности, то опускающиеся на глубину до 2-3 км.
Некоторые мелководные виды каланид в умеренных водах развиваются в огромном количестве и по весу составляют преобладающую часть планктона. Так, например, планктон Баренцева моря приблизительно на 90% состоит из Calanus finmarchicus (табл. 31, 3). Эти рачки характеризуются высокой питательностью: в их теле содержится 59% белков, 20% углеводов и часто более 10-15% жиров. Многие рыбы, а также усатые киты питаются главным образом каланидами. Таковы, например, сельдь, сардина, скумбрия, анчоус, шпрот и многие другие. В желудке одной сельди было обнаружено 60 000 проглоченных ею веслоногих рачков. Из китов активно потребляют огромные массы каланид финвал, сейвал, синий кит и горбач.
Calanoida внутренних водоемов по своей биологии напоминают морские виды. Они также приурочены только к толще воды, также совершают вертикальные миграции и питаются так же, как морские фильтраторы. Они обитают в самых разнообразных водоемах. Некоторые виды, как например Diaptomus graciloides и D. gracilis, живут почти во всех озерах и прудах северной и средней части СССР. Другие приурочены только к Дальнему Востоку или к южной части нашей страны. Весьма интересно распространение Limnocalanus grimaldii, который населяет многие озера севера нашей страны (в том числе Онежское и Ладожское) и Скандинавии. Этот вид близок к прибрежному солоновато-водному L. macrurus, обитающему в пред-устьевых пространствах северных рек. Озера, населенные L. grimaldii, некогда покрывались холодным Иольдиевым морем. В Байкале в огромном количестве обитает свойственный только этому озеру рачок Epischura baicalensis, служащий основной пищей омулю. Очень своеобразны, хотя и еще мало известны, условия существования недавно открытого единственного подземного представителя каланид - Speodiaptomus birsteini.
Этот слепой рачок найден в глубоких и узких заполненных водой расселинах нижнего этажа Скельской пещеры, расположенной в Крыму, недалеко от Севастополя. За ним удалось наблюдать в аквариуме, причем оказалось, что он плавает так же, как обычные пресноводные каланиды. Остается загадкой, чем он питается, фильтруя чистую, полностью лишенную водорослей и, вероятно, очень бедную бактериями воду подземного бассейна. По-видимому, он может считаться единственным настоящим подземным планктонным животным.
Некоторые пресноводные каланиды появляются в водоемах только в определенное время года, например весной. В весенних лужах часто попадается сравнительно крупный (около 5 мм) Diaptomus amblyodon, окрашенный в яркий красный или синий цвет. Этот вид и некоторые другие широко распространенные пресноводные каланиды способны к образованию покоящихся яиц, стойко переносящих высыхание и вымерзание и легко разносящихся ветром на далекие расстояния.
Подотряд Циклопы (Cyclopoida)
Другой подотряд веслоногих ракообразных - Cyclopoida - наибольшим количеством видов представлен в пресных водах.
Пресноводные циклопы живут во всевозможных водоемах, от мелких луж до крупных озер, причем нередко встречаются в очень большом количестве экземпляров. Основной зоной их обитания служит прибрежная полоса с зарослями водных растений. При этом во многих озерах к зарослям определенных растений приурочены определенные виды циклопов. Так, например, для Валдайского озера в Ивановской области описано 6 группировок растений с соответствующими им группировками видов циклопов.
Сравнительно немногие виды могут считаться настоящими планктонными животными. Некоторые из них, принадлежащие преимущественно к роду Mesocyclops, постоянно обитают в поверхностных слоях воды, другие (Cyclops strenuus и другие виды того же рода) совершают регулярные суточные миграции, опускаясь днем на значительную глубину.
Плавают циклопы несколько иначе, чем каланиды. Одновременно взмахивая четырьмя парами грудных ножек (пятая пара редуцирована), рачок делает резкий прыжок вперед, вверх или вбок, а затем при помощи передних антенн может некоторое время парить в воде. Поскольку центр тяжести его тела смещен вперед, во время парения его передний конец наклоняется и тело может принять вертикальное положение, причем погружение замедляется. Новый взмах ножками позволяет циклопу подняться. Эти взмахи молниеносны - они занимают 1/60 секунды.
Много занимавшийся биологией циклопов Л. Исаев описывает их движения следующим образом: "Передвигаясь ритмическими скачками, циклоп может хорошо держаться на одном уровне, подниматься вверх и опускаться вниз под углами различной крутизны. Циклоп может плавать с одинаковой легкостью, перевернувшись на спину. Циклоп хорошо описывает дуги, делает мертвые петли, одинарные и множественные, прямые и обратные. Циклоп может делать поворот под углом 90°, вращаться вокруг оси не только со снижением, напоминающим витки "штопора" самолета, но и с поднятием вверх. Циклоп может скользить на антенну, делать через нее переворот, пикировать вниз головой под углом в 90° и скользить на хвост. Характер "фигур", выполняемых циклопом, очень сходен с фигурами высшего пилотажа. Обладание фигурами высшего пилотажа, необходимыми для самолетов-истребителей, несомненно облегчает циклопу - активному хищнику - возможность обеспечить себе существование охотой за водными обитателями, служащими ему пищей".
Большинство циклопов хищники, но среди них есть и растительноядные виды. Такие обычные, широко распространенные виды, как Macrocyclops albidus, M. fuscus, Acanthocyclops viridis и многие другие, быстро плавают над дном или среди зарослей в поисках добычи. При помощи своих антенн на очень коротком расстоянии они чуют мелких олигохет и хирономид, которых хватают вооруженными шипами передними челюстями. В передаче пищи жвалам участвуют задние челюсти и ногочелюсти. Жвалы совершают быстрые режущие движения в течение 3-4 секунд, за которыми следует минутная пауза. Циклопы могут поедать олигохет и хирономид более крупных, чем они сами. Скорость поедания жертв зависит от их размеров и от твердости их покровов. Для размельчения и заглатывания мотыля длиною 2 мм требуется 9 минут, а личинка длиною 3 мм уничтожается уже в течение получаса. Более нежный, хотя и более длинный (4 мм), малощетинковый червь Nais поедается всего за 3,5 минуты.
Хищные циклопы нападают не только на таких малоподвижных животных, как сидящие на дне или на растениях олигохеты и мотыли, но и на плавающих в воде других рачков, в том числе и на себе подобных, а также на коловраток и т. д. Они охотно заглатывают и личинок паразитических червей - корацидиев широкого лентеца и личинок ришты (см. том 1) - и становятся, таким образом, промежуточными хозяевами этих паразитов человека. Неоднократно наблюдалось нападение циклопов на икринки и даже личинок рыб.
Растительноядные циклопы, в частности обычные Eucyclops macrurus и E. macruroides, питаются главным образом зелеными нитчатыми водорослями (Scenedesmus, Micractinium), захватывая их приблизительно так же, как хищные захватывают червей и мотылей; кроме того, используются различные диатомовые, перидиниевые и даже сине-зеленые водоросли. Многие виды могут поедать только относительно крупные водоросли. Mesocyclops leuckarti быстро набивает себе кишечник колониями Pandorina (диаметр колонии 50-75 мк) и почти совсем не заглатывает мелких Chlamydomonas.
Пресноводные циклопы распространены очень широко. Некоторые виды встречаются почти повсеместно. Этому способствуют прежде всего приспособления к перенесению неблагоприятных условий, в частности способность рачков переносить высыхание водоемов и в виде цист пассивно расселяться по воздуху. Кожные железы многих циклопов выделяют секрет, обволакивающий тело рачка, часто вместе с яйцевыми мешками, и образующий нечто вроде кокона. В таком виде рачки могут подвергаться высыханию и вмерзанию в лед, не теряя жизнеспособности. В опытах Камерера циклопы быстро выводились при размачивании сухого ила, сохранявшегося около 3 лет. Поэтому нет ничего удивительного в появлении циклопов в весенних лужах, возникающих при таянии снега, в только что залитых рыбоводных прудах и т. д.
Второй причиной широкого распространения многих видов циклопов следует считать стойкость рачков, находящихся в активном состоянии, по отношению к недостатку кислорода в воде, кислой ее реакции и многим другим факторам, неблагоприятным для остальных пресноводных животных. Cyclops strenuus в течение нескольких дней может жить не только при полном отсутствии кислорода, но даже в присутствии сероводорода. Некоторые другие виды также хорошо переносят неблагоприятный газовый режим. Многие циклопы превосходно существуют в воде с кислой реакцией, при высоком содержании гуминовых веществ и крайней бедности солей, например в водоемах, связанных с верховыми (сфагновыми) болотами.
Тем не менее известны виды и даже роды циклопов, ограниченные в своем распространении какими-нибудь определенными, в частности температурными и солевыми, условиями. Так, например, род Ochridocyclops обитает только в озере Охрид в Югославии, род Bryocyclops - в Юго-Восточной Азии и в экваториальной Африке. К последнему роду близок исключительно подземный род Speocyclops, виды которого найдены в пещерах и грунтовых водах Южной Европы, Закавказья, Крыма и Японии. Эти слепые мелкие рачки считаются остатками некогда более широко распространенной теплолюбивой фауны.
Некоторые циклопы приспособились к жизни в солоноватых и даже в очень соленых водоемах. Род Halicyclops, например, весьма обычен в Каспийском море и не встречается в пресной воде. Microcyclops dengizicus широко распространен только в солоноватых и соленых водоемах зоны пустынь (Ирак, Индия, о. Гаити, Египет, Калифорния, в СССР - в Карагандинской области, в Муганской степи) и прекрасно живет даже при солености, превосходящей морскую (до 410/00). Многие обычные пресноводные виды могут существовать и в солоноватой воде, как например Mesocyclops leuckarti в Финском и Ботническом заливах.
Значение циклопов в жизни наших внутренних водоемов очень велико. С одной стороны, они служат пищей многим рыбам и их молоди, но, с другой стороны, выступают в качестве конкурентов тех же самых рыб, поедая множество мелких животных, как планктонных, так и донных. Не следует забывать и о том, что циклопы способствуют заражению человека некоторыми паразитическими червями, в первую очередь широким лентецом (Diphyllobothrium latum) и риштой (Dracunculus medinensis). Циклопы заглатывают плавающих в воде личинок этих червей. Корацидий широкого лентеца, попав в циклопа, сбрасывает свой мерцательный покров, проникает из кишечника в полость тела рачка и превращается там в стадию процеркоида. Если такой циклоп будет проглочен рыбой, процеркоид перейдет в стадию плевроцеркоида, а затем, при поедании рыбы человеком, кошкой и другими млекопитающими, из плевроцеркоида возникнет взрослый червь, часто вызывающий тяжелое малокровие. Личинки ришты, проглоченные циклопами, также проникают в полость их тела и остаются там в течение 1-1,5 месяцев. При питье сырой воды человек проглатывает циклопов и заражается риштой.
Морские представители подотряда Cyclopoida менее разнообразны, чем пресноводные. Среди них обычны и нередко многочисленны в морском планктоне виды рода Oithona. Очень характерны также крупные (до 8 мм) уплощенные виды рода Sapphirina, поверхность тела которых отливает яркими синими, золотистыми или темно-красными тонами (табл. 31, 1). Другой близкий морской род - Oncaea (табл. 31, 4) - обладает железами, выделяющими светящийся секрет, и нередко, совместно с другими организмами, вызывает свечение моря.
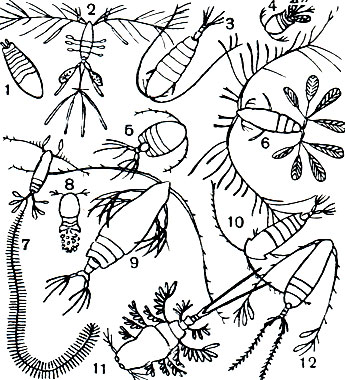
Таблица 31. Веслоногие ракообразные: 1 - Sapphirina ovatolanceolata; 2 - Oithona plumifera; 3 - Galanus finmarchicus; 4 - Oncaea mediterranea; 5 - Phoenna spinifera; 6 - Galocalanus pavo; 7 - Sapphirina vorax; 8 - Corycaeus obtusus; 9 - Euchirella maxima; 10 - Pontella mediterranea; 11 - Copilia vitrea; 12 - Calanus plumchrus

Таблица 31. Веслоногие ракообразные: 1 - Sapphirina ovatolanceolata; 2 - Oithona plumifera; 3 - Galanus finmarchicus; 4 - Oncaea mediterranea; 5 - Phoenna spinifera; 6 - Galocalanus pavo; 7 - Sapphirina vorax; 8 - Corycaeus obtusus; 9 - Euchirella maxima; 10 - Pontella mediterranea; 11 - Copilia vitrea; 12 - Calanus plumchrus
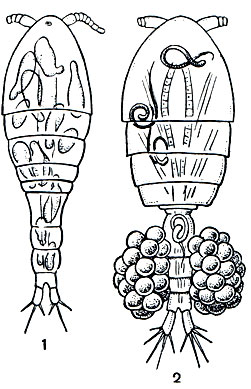
Рис. 208. Циклопы - промежуточные хозяева паразитов человека: 1 - Paracyclops с процеркоидами лентеца широкого в полости тела; 2 - Mesocyclops с личинками ришты в полости тела
Подотряд Гарпактициды (Harpacticoida)
Об образе жизни представителей третьего подотряда - Harpacticoida - известно гораздо меньше. Эти червеобразные, большей частью очень мелкие рачки, чрезвычайно разнообразны как в морских, так и в пресных водах, но никогда не встречаются в массовом количестве. Описано более 30 семейств и несколько сотен видов Harpacticoida.
Большинство гарпактицид ползает по дну и донным растениям. Лишь единичные виды способны продолжительное время плавать и входят в состав морского планктона (Microsetella). Гораздо более характерны целые группы родов и видов гарпактицид, приспособившиеся к обитанию в особых, необычных условиях, в частности в капиллярных ходах между песчинками морских пляжей и в подземных пресных водах.
Всего несколько лет тому назад зоологи применили очень простой прием, позволяющий изучать население капиллярных ходов морского песка. На пляже, выше уровня моря, выкапывается яма. В ней постепенно накапливается вода, заполнявшая капилляры песка. Эту воду фильтруют через планктонную сеть и таким образом добывают представителей своеобразной фауны, получившей название интерстициальной.
Гарпактициды составляют заметную часть интерстициальной фауны. Их обнаружили всюду, где проводились соответствующие исследования,- на пляжах Англии, вдоль европейского и американского берегов Атлантического океана, на Средиземном и Черном морях, у берегов Африки и Индии, на островах Мадагаскар, Реюньон и Багамских. Большинство интерстициальных гарпактицид принадлежит к особым, обитающим только в таких условиях родам, отличающимся необыкновенно тонким и длинным телом (рис. 209), позволяющим рачкам двигаться в узких капиллярных ходах. Замечательно, что некоторые из этих специализированных видов были найдены в очень отдаленных друг от друга местах. Так, например, на Багамских островах оказались Arenosetella palpilabra, ранее известная только из Шотландии, и Horsiella trisaetosa, ранее известная только из Кильской бухты. Трудно объяснить подобное распространение, поскольку покоящихся яиц у интерстициальных гарпактицид нет.
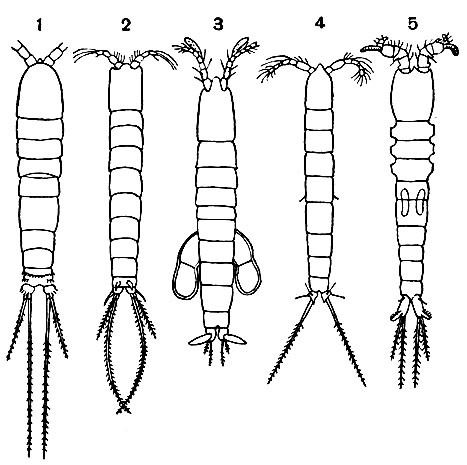
Рис. 209. Характерные интерстициальные гарпактициды: 1 - Nitocra chelifer; 2 - Arenosetella spinicauda; 3 - Goffinella stylifer; 4 - Paraleptastacus brevicaudatus; 5 - Emertonia gracilis
Гарпактициды пресных подземных вод также представлены рядом специализированных родов - Parastenocaris, Elaphoidella, Ceuthonectes и других, частично очень широко распространенных, частично имеющих узкие и разорванные ареалы. Так, например, два вида рода Ceuthonectes обитают только в пещерах Закавказья, Югославии, Румынии, Италии и Южной Франции. Эти удаленные друг от друга местонахождения считаются остатками некогда гораздо более обширной области распространения древнего рода. В некоторых случаях можно допускать тропическое происхождение подземных гарпактицид Европы. Среди многочисленных видов рода Elaphoidella есть как тропические, так и европейские виды. Первые обитают в наземных, вторые (за единичными исключениями) в подземных водах. По всей вероятности, под землей сохранились остатки древней тропической фауны, погибшей на поверхности земли под влиянием климатических изменений. В тропических наземных водоемах некоторые гарпактициды приспособлены к условиям существования, напоминающим условия жизни в подземных водах. Известны тропические виды Elaphoidella, живущие в своеобразных микроводоемах, образующихся в пазухах листьев водных растений из семейства Bromeliaceae. В таких же условиях живет тропическая Viguierella coeca, обнаруженная на этих растениях в ботанических садах почти всех стран.
Чрезвычайно богата видами гарпактицид своеобразная фауна Байкала. Она состоит из 43 видов, из которых 38 эндемичны для этого озера. Особенно много этих рачков в прибрежной части Байкала, на камнях и водных растениях, а также на растущих здесь губках. По-видимому, они питаются за счет губок и в свою очередь становятся жертвами также ползающего по губкам бокоплава Brandtia parasitica.
Некоторые виды гарпактицид приурочены только к водоемам, очень бедным солями, характеризующимся повышенной кислотностью, т. е. связанным с верховыми, сфагновыми, болотами. Таков, например, Arcticocamptus arcticus, биология которого подробно изучена Е. В. Боруцким.
A. arcticus широко распространен на севере Европы, от Болынеземельской тундры до Скандинавии, на западном побережье Гренландии, на Новой Земле. Кроме того, он найден в Альпах и в нескольких пунктах средней полосы Европейской части СССР, в том числе в Косине под Москвой, в Звенигороде, под Ярославлем и т. д. Повсюду он обитает в водоемах, связанных со сфагновыми болотами.
Из многочисленных водоемов, расположенных близ Косина, рачок живет только в двух болотах и в Святом озере, лежащем среди сфагнового торфяника. Очевидно, условия существования в других разнообразных соседних водоемах неблагоприятны для A. arcticus. При этом даже в немногих населенных им водоемах рачок в активном состоянии существует только в течение 11/2-2 месяцев весной, остальную часть года, т. е. 10-101/2 месяцев, он проводит в стадии покоящегося яйца.
Жизненный цикл A. arcticus тесно связан с изменениями растительного покрова болота. Е. В. Боруцкий пишет: "Как только с первыми весенними теплыми лучами рыхлый снег начинает таять и на поверхности болота образуются лужи - все животные, проведшие суровую зиму в тех или других стадиях в ледяном заключении, начинают реагировать на первые весенние лучи. A. arcticus одним из первых выходит из состояния анабиоза и появляется в водоеме. Уже в небольших лужах, еще среди снега, там, где поверхностные слои сфагнума оттаяли, можно найти его личинок, медленно и неуклюже передвигающихся среди листочков мха в поисках пищи. Личинки - на первой науплиальной стадии и, очевидно, только что вылупившиеся из яйца. С каждым днем науплиус крепнет, движения его становятся более уверенными и быстрыми, наконец, наступает момент первой линьки - он меняет старую узкую оболочку на новую, более просторную. За первой линькой следует вторая, третья и т. д., и вот спустя две-три недели мы уже встречаем взрослые экземпляры или личинок на последней копеподитной стадии. Но они уже не пользуются тем простором, который имели в своем раннем личиночном состоянии: вместо обширных луж, полных воды, в которых они свободно переплывали с одного сфагнового кустика на другой, теперь только влажный мох и незначительное количество воды. Вместо жалких голых ветвей теперь нежные розовые цветы Кассандры и клюквы, белые чашечки андромеды и шапки цветущего богульника. Болото изменилось - яркий зеленый сфагновый ковер пестрит розовыми и белыми пятнами цветов. И это изменение картины болота как нельзя лучше совпадает с определенным моментом в биологии A. arcticus, именно с периодом копуляции. В продолжение нескольких дней мы встречаем почти исключительно копулирующие пары. Но эти цветы - роковые для A. arcticus: с постепенным увяданием их наблюдается постепенное уменьшение численности рачков, копуляция встречается все реже и реже, чаще попадаются самки с яйцевыми мешками и, наконец, к середине или к концу июня A. arcticus совсем исчезает из водоема, и лишь только запоздалые экземпляры встречаются в незначительном количестве в июле или в начале августа".
Рачок оставляет в водоеме свои яйцевые мешки, имеющие форму двух соединенных вместе шаров, покрытых общей "мешковой" оболочкой, осуществляющей механическую защиту яиц, а также защищающей их от высыхания. Кроме того, каждое яйцо имеет свою более тонкую прозрачную оболочку. Она непроницаема как для солей, так и для воды. К осени в каждом яйце формируется науплиус, причем науплиусы двух соединенных яиц всегда направлены своими передними концами в противоположные стороны. Науплиусы одеты еще одной очень тонкой и эластичной внутренней оболочкой, снабженной различными тяжами и нитями. Эта оболочка проницаема для воды, но не для солей.
Когда наступает время вылупления науплиусов, т. е. в период весеннего таяния льда, в мешковой оболочке с одной стороны образуется трещина, через которую начинает выпячиваться эластичная внутренняя оболочка. Сначала этот процесс идет очень медленно, но после того, как около половины окруженной оболочкой личинки окажется снаружи, происходит резкий толчок и личинка, заключенная в полый шар, как бы "выстреливает" из яйцевого мешка и отскакивает в сторону или задерживается за края щели придатками эластичной оболочки. Замечательно, что сам науплиус при этом остается почти все время совершенно пассивным. Только в самом начале процесса вылупления науплиус производит несколько довольно слабых движений, приводящих, по-видимому, к разрыву яйцевой оболочки. Основную роль здесь играет полупроницаемая эластичная оболочка, через которую диффундирует вода, вызывающая ее разбухание, что сначала заставляет лопнуть мешковую оболочку, а затем выпятиться из нее окруженному эластичной оболочкой науплиусу. Тяжи и нити этой оболочки действуют как пружины, причем расположены они таким образом, что науплиус внутри образованного оболочкой полого шара всегда выходит своим головным концом вперед. За первым науплиусом через ту же щель в мешковой оболочке выпячивается таким же способом второй, или оба "выстреливают" одновременно. Первым толчком к разбуханию эластичной оболочки служит, по-видимому, разрыв науплиусом яйцевой оболочки (рис. 210).
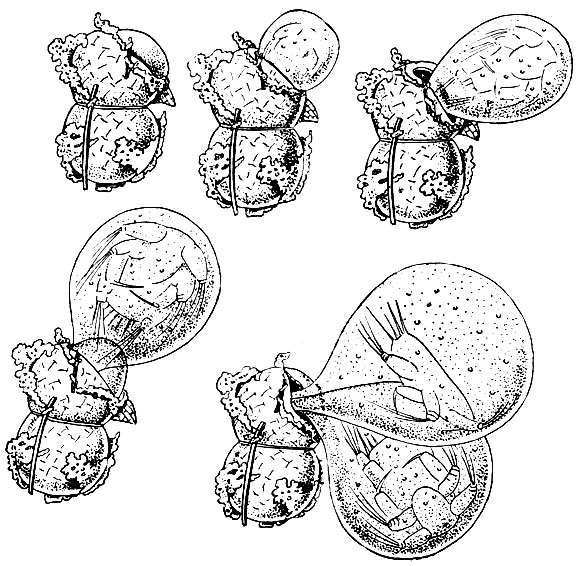
Рис. 210. Стадии вылупления науплиуса Arcticocamptus arcticus из яйца
Только через некоторое время новорожденный науплиус, находящийся внутри полностью раздутой, принявшей шаровидную форму эластичной оболочки, начинает двигаться и пытается ее разорвать. Это удается ему не сразу, после чего оболочка спадается и личинка оказывается на свободе. Утомленная тяжелой работой, она сначала почти не способна быстро передвигаться, но это ей и не нужно, так как она находит достаточное количество пищи на поверхности только что покинутой мешковой оболочки, которая обычно за многомесячное пребывание в водоеме обрастает водорослями и покрывается частицами детрита.
При помещении яиц A. arcticus в воду неблагоприятных для его существования водоемов науплиусы внутри яиц развивались нормально, но их вылупления не происходило. Специально поставленными опытами Е. В. Боруцкий доказал, что при сравнительно высоком содержании солей в воде не происходит диффузии воды через эластичную внутреннюю оболочку и она не разбухает.
Если вода не имеет кислой реакции, яйцевая оболочка частично не растворяется и не размягчается, что также исключает возможность вылупления науплиуса. Таким образом, обе эти оболочки препятствуют вылуплению науплиуса при попадании яйца в неблагоприятные для рачка условия, обрекающие его на гибель. Действительно, личиночные стадии и взрослые рачки погибали в воде низовых (несфагновых) болот, а также других водоемов, содержащей обычное количество солей и имеющей нейтральную или щелочную реакцию. Все это делает понятным строгую приуроченность A. arcticus к верховым, сфагновым, болотам с их специфическим гидрохимическим режимом.
Если A. arcticus в активном состоянии существует весной, то некоторые другие виды пресноводных гарпактицид встречаются только зимой или только летом. При этом известны виды, проводящие период покоя не в стадии покоящегося яйца, как A. arcticus, а в стадии цист, несколько напоминающих описанные выше цисты циклопов. У Canthocamptus staphylinus такие цисты округлые, у Attheyella wulmeri и A. northumbrica они овальные, причем фуркальные щетинки рачка торчат из оболочки (рис. 211).
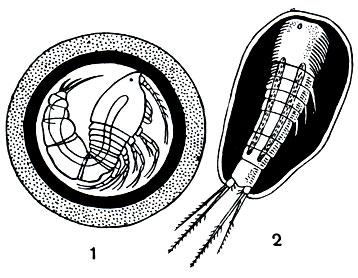
Рис. 211. Цисты гарпактицид: 1 - Canthocamptus staphylinus; 2 - Attheyella wulmeri
Среди пресноводных гарпактицид есть виды, способные к партеногенетическому размножению, не свойственному всем остальным веслоногим ракообразным. У широко распространенной в Европе Еlаphoidella bidens самцы вообще неизвестны, а в условиях эксперимента у этого вида удалось получить 5 поколений партеногенетических самок. К партеногенетическому размножению оказался способным и Epactophanes richardi, хотя в естественных условиях он представлен как самками, так и самцами. По-видимому, и некоторые другие виды гарпактицид могут размножаться партеногенетически.
Практическое значение гарпактицид несравненно меньше, чем каланид и циклопов. В некоторых водоемах они составляют заметную часть пищи рыб, в особенности их молоди.
Паразитические веслоногие
Паразитические веслоногие ракообразные прикрепляются к самым различным рыбам, как морским, так и пресноводным, а также ко многим морским беспозвоночным - сифонофорам, кораллам, моллюскам, полихетам, эхиуридам, высшим ракообразным, иглокожим и к некоторым другим. Как правило, каждый вид паразита паразитирует на определенном виде хозяина, однако есть и менее разборчивые, способные использовать разные виды рыб.
Большинство паразитических веслоногих представляет собой эктопаразитов. Они поселяются на наружных покровах, на жабрах, на глазах, реже в ротовой полости рыб. Среди паразитов морских беспозвоночных много эндопаразитов. Так, например, Monstrillidae живут в кровеносных сосудах полихет, Mytilicola -в кишечнике моллюсков, несколько родов- в кишечнике иглокожих.
Паразитические веслоногие принадлежат к трем подотрядам - Cyclopoida, включающему наряду с паразитическими и только что рассмотренные свободноживущие виды, Caligoida и Lernaeoida, в состав которых входят исключительно паразитические виды.
Паразитические Cyclopoida отличаются от свободноживущих прежде всего строением своих головных придатков. Задние антенны крючковидные, приспособленные для прикрепления к хозяину. Жвалы, если есть, не бывают жующего типа, другие ротовые части также видоизменены или отсутствуют. Тело некоторых видов сегментировано так же, как у свободно-живущих, но у других сегментация в той или иной степени исчезает. У таких специализированных видов редуцируются и грудные ножки. К этому подотряду относится большое количество паразитов пресноводных и морских рыб, а также морских беспозвоночных.
Одними из самых обычных паразитов как пресноводных, так и морских рыб можно считать представителей рода Ergasilus, прикрепляющихся к жабрам линей, щук и многих других рыб. Нередко к одной рыбе прикрепляется по нескольку десятков, а иногда даже до 3000 рачков, что приводит ее к гибели. Замечено, что, чем старше рыба, тем больше рачков на ней паразитируете Рачки охватывают жаберные стебельки своими крючковидными задними антеннами, при смыкании образующими кольцо, на котором паразит, по выражению В. А. Догеля, болтается, как замок на скобе (рис. 212, 3). Постепенно эпителий жабр разрушается.
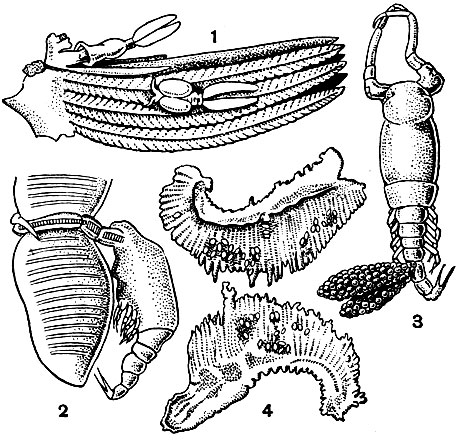
Рис. 212. Ergasilus sieboldi: 1 - рачок на жабре линя; 2 - рачок на жаберном лепестке; 3 - то же, вид сверху; 4 - жабры линя, пораженные Ergasilus
Вышедшие из яиц личинки Ergasilus ведут свободный образ жизни. Через 2-21/2 месяца рачки достигают половозрелости и спариваются. Оплодотворенные самки активно двигаются против течения. Это помогает им оседать на жабры рыб, поскольку из-под жаберной крышки направляется ток воды.
Таким же путем жабры рыб поражаются глохидиями перловиц (см. выше). Интересно, что между эргазилидами и глохидиями существует антагонизм: одни вытесняют других и совместно на жабрах одной рыбы не встречаются.
Если эргазилиды паразитируют во взрослом состоянии и ведут свободный образ жизни на ранних стадиях своего развития, то представители семейства Monstrillidae, паразитирующие в морских многощетинковых червях, имеют совсем другой жизненный цикл. Взрослые рачки оживленно плавают подобно обычным свободноживущим веслоногим. Замечательно, однако, что они полностью лишены кишечника и совсем не питаются. Из яиц этих самок выходят также свободноплавающие науплиусы, как и взрослые, лишенные кишечника. Науплиусы отыскивают определенные виды полихет (из родов Polydora, Salmacina и др.) и внедряются в них, проникая в спинной кровеносный сосуд.
Здесь науплиус линяет, превращаясь в многоклеточное овальное тельце. Впоследствии у этого зародыша на переднем конце вырастают два длинных придатка, служащих для всасывания пищи. Зародыш линяет еще раз и преобразуется в длинное колбасовидное тело, внутри которого формируется взрослый рачок с хорошо развитыми половыми органами. Он прорывает стенку кровеносного сосуда и покровы хозяина и приступает к активному существованию (рис. 213).
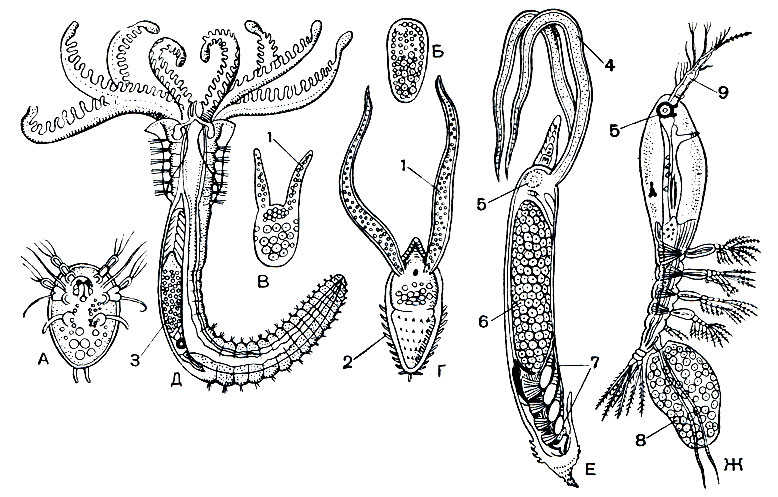
Рис. 213. Развитие Haemocera danae (сем. Monstrillidae): А - свободноплавающий науплиус; В - науплиус после проникновения в червя; В, Г - две стадии зародыша в кровеносном сосуде червя; Д - многощетинковый червь Salmacina danae с почти взрослой самкой рачка в спинном кровеносном сосуде; Е - самка из кровеносного сосуда Salmacina; Ж - свободноплавающая самка; 1 - отростки зародыша; 2 - шипы на заднем конце его тела; 3 - самка; 4 - всасывающие пищу отростки самки; 5 - глаз; 6 - яичник; 7 - зачатки грудных ножек; 8 - яйцевой мешок; 9 - передняя антенна
Из паразитов беспозвоночных наибольшее хозяйственное значение имеет Mytilicola (рис. 214). Взрослые самки рачка достигают 8 мм длины и живут в кишечнике мидий и устриц. Тело паразита вытянутое, червеобразное, но следы сегментации сохраняются; грудные ножки редуцированы. Науплиусы и метанауплиусы живут свободно, первая копеподитная личинка внедряется в кишечник моллюсков и превращается там во взрослого рачка, Заражение Mytilicola вызывает массовую гибель мидий. Запасы этих промысловых моллюсков у берегов Голландии в результате заражения рачками в 1949 и 1950 годах уменьшились в 10 раз.
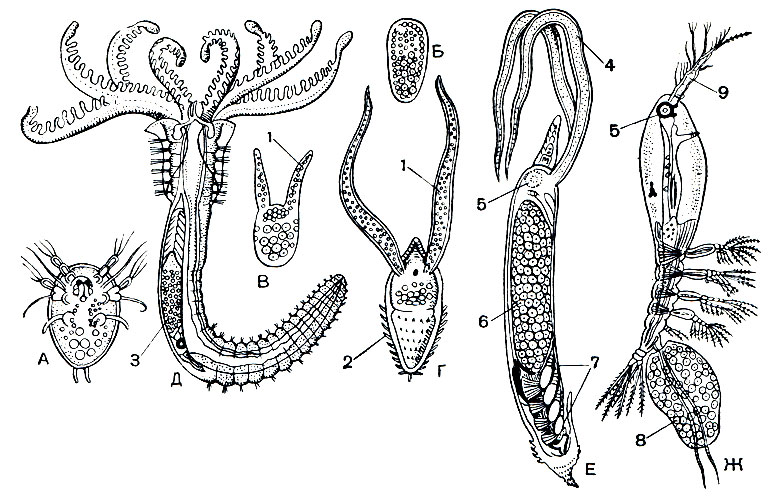
Рис. 214. Самка Mytilicola (справа крайняя)
Представители подотряда Caligoida характеризуются расширенным, сплющенным в спинно-брюшном направлении телом, сегментация грудного отдела в той или иной степени утеряна, самки имеют очень крупный передний брюшной (генитальный) сегмент, к которому прикрепляются два яйцевых мешка, ротовые придатки образуют хоботок, позволяющий высасывать кровь хозяина. Самки и самцы мало различаются по величине и по строению.
Эти рачки паразитируют главным образом на рыбах, как морских, так и пресноводных, но род Archicaligus живет на замечательном головоногом моллюске Nautilus. Взрослые особи могут на некоторое время покидать хозяина, плавать, а затем садиться на другого. Науплиусы ведут свободное существование. Личинка на копеподитной стадии снабжена специальной лобной нитью, при помощи которой она прикрепляется к хозяину. После этого она переходит в стадию куколки, помещающейся в кутикулярной оболочке предыдущей стадии, неподвижной и непитающейся. Куколка линяет и превращается в также прикрепленную последнюю личиночную стадию - халимус, которая преобразуется, наконец, во взрослого рачка (рис. 215, Б).
Некоторые калигиды, будучи сами паразитами рыб, подвергаются нападению других паразитов. Так, например, на Lepeophtheirus, живущем на многих морских рыбах, паразитируют плоские черви Udonella и инфузории Conidophrys, выступающие в качестве "сверхпаразитов", т. е. паразитов паразита (рис. 215, А).
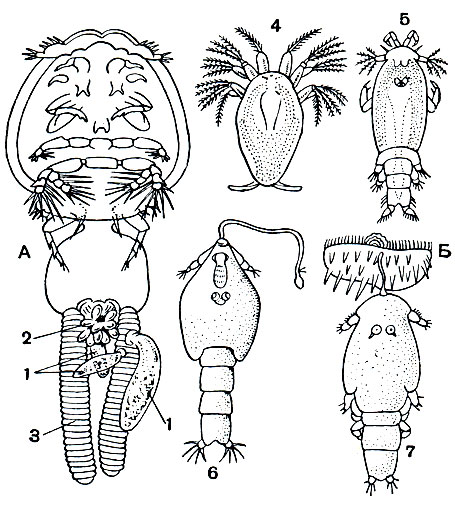
Рис. 215. Калигоидные рачки: А - Lepeophtheirus с гиперпаразитами - Udonella (1) и Conidophrys (2); 3 - яйцевой мешок рачка; В - личиночные стадии Caligus: 4 - науплиус, 5 - копеподитная личинка, 6 - куколка, 7 - халимус
Последний подотряд паразитических веслоногих - Lernaeoida - объединяет наиболее сильно видоизмененных под влиянием паразитизма рачков. Взрослые половозрелые самки обычно имеют червеобразное тело без всяких следов сегментации или даже разделения на отделы тела, с выростами на переднем конце, погруженными в кожу хозяина, и с двумя яйцевыми мешками на заднем конце. Самцы гораздо мельче самок и по своему строению напоминают копеподитные стадии. Рачки паразитируют на рыбах, реже на амфибиях и китах. Некоторые из них достигают очень крупных размеров: паразит китов Penella balaenoptera длиною 32 см может считаться крупнейшим представителем веслоногих. Он погружен в наружные покровы китов на 5-7 см.
Развитие этих рачков обычно укорочено. Стадии науплиуса и метанауплиуса они проходят в яйце. Из него вылупляется свободная копеподитная личинка, достигающая половозрелости. Вскоре происходит копуляция, после которой самцы умирают. Молодые самки семейства Lernaeidae сначала паразитируют на одних рыбах, например на камбаловых, а затем переходят на тресковых, теряя следы сегментации, конечности и превращаясь в червеобразные существа (рис. 216). Первым хозяином гигантской Penella служат головоногие моллюски, а вторым - киты. У представителей другого обширного семейства этого подотряда - Lernaeopodidae - смены хозяев не происходит и копеподитная личинка совершает свой метаморфоз на одном хозяине.
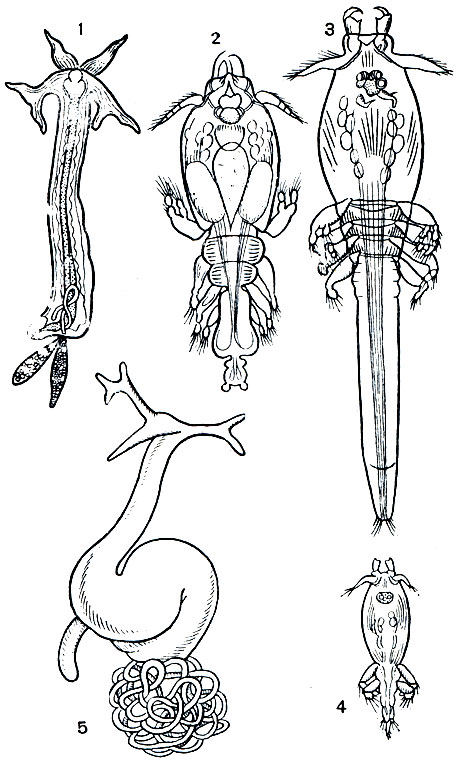
Рис. 216. Стадии развития Lernaeocera branchialis: 1 - взрослая самка с еще недоразвитыми яйцевыми мешками; 2 - самец с жабр камбалы; 3 - неоплодотворенная самка; 4 - копеподитная свободноплавающая личинка; 5 - взрослая самка с развитыми яйцевыми мешками с жабр трески
Лернеиды нередко вызывают гибель пораженных ими рыб, как морских, так и пресноводных. Они вредят лососевым рыбам, разрушая их жабры, описаны случаи смерти морских рыб, вызванной этими рачками. Глубоко внедряясь в тело рыб, некоторые из лернеид достигают сердца и печени своих хозяев, другие вызывают образование кровоточащих ран на поверхности тела хозяина. Кроме того, питаясь кровью рыб, эти паразиты сильно их истощают.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'