Тип Оболочники (Tunicata) (Н. Г. Виноградова)
Оболочники, или туникаты, к которым относятся асцидии, пиросомы, сальпы и аппендикулярии,- одна из наиболее удивительных групп морских животных. Свое название они получили за то, что тело их одето снаружи особой студенистой оболочкой, или туникой. Туника состоит из вещества, чрезвычайно близкого по своему составу к целлюлозе, которая встречается только в царстве растений и неизвестна ни для одной другой группы животных. Оболочники - исключительно морские животные, ведущие частично прикрепленный,частично свободноплавающий пелагический образ жизни. Они могут быть или одиночными, или образуют удивительные колонии, возникающие при чередовании поколений в результате почкования бесполых одиночных особей. О способах размножения этих животных - наиболее необычайных среди всех живых существ на Земле - мы специально будем говорить ниже.
Очень интересно положение оболочников в системе животного царства. Природа этих животных долго оставалась загадочной и непонятной, хотя они были известны еще Аристотелю более двух с половиной тысяч лет назад под именем Tethya. Только в начале XIX столетия было установлено, что одиночные и колониальные формы некоторых оболочников - сальп - представляют собой лишь различные поколения одного и того же вида. До тех пор их относили к разным видам животных. Эти формы отличаются друг от друга не только по внешнему виду. Оказалось, что только колониальные формы обладают половыми органами, а одиночные - бесполы. Явление чередования поколений у сальп было открыто поэтом и естествоиспытателем Альбертом Шамиссо во время его плавания в 1819 г. на русском военном корабле "Рюрик" под командованием Коцебу. Старые авторы, в том числе и Карл Линней, относили одиночных туникат к типу моллюсков. Колониальные же формы были отнесены им к совершенно другой группе - зоофитам, а некоторые считали их особым классом червей. Но на самом деле эти внешне очень просто устроенные животные не так примитивны, как они кажутся. Благодаря работам замечательного русского эмбриолога А. О. Ковалевского в середине прошлого века было установлено, что оболочники близки к хордовым животным. А. О. Ковалевский установил, что развитие асцидий идет по тому же самому типу, что и развитие ланцетника, представляющего, по меткому выражению академика И. И. Шмальгаузена, "как бы живую упрощенную схему типичного хордового животного". Группа хордовых животных характеризуется целым рядом определенных важных черт строения. Прежде всего это наличие спинной струны, или хорды, представляющей собой внутренний осевой скелет животного. Личинки туникат, свободно плавающие в воде, также имеют спинную струну, или хорду, которая совершенно исчезает при превращении их во взрослую особь. Личинки и по другим важнейшим признакам строения стоят гораздо выше родительских форм. По филогенетическим причинам, т. е. по причинам, связанным с происхождением группы, большее значение у оболочников придается организации их личинок, чем организации взрослых форм. Такая аномалия неизвестна более ни для одного другого типа животных. Кроме наличия хорды, хотя бы в личиночной стадии, с настоящими хордовыми животными оболочников сближает целый ряд других признаков. Очень важно, что нервная система оболочников расположена на спинной стороне тела и представляет собой трубку, имеющую внутри канал. Нервная трубка оболочников образуется как желобообразное продольное впячивание поверхностных покровов тела зародыша, эктодермы, как это имеет место у всех других позвоночных животных и у человека. У беспозвоночных животных нервная система всегда лежит на брюшной стороне тела и образуется иным способом. Основные сосуды кровеносной системы оболочников, наоборот, располагаются на брюшной стороне, в противоположность тому, что характерно для беспозвоночных животных. И наконец, передний отдел кишки, или глотка, пронизан у оболочников многочисленными отверстиями и превратился в орган дыхания. Как мы видели в других главах, у беспозвоночных животных органы дыхания очень разнообразны, но никогда кишечник не образует жаберных щелей. Это признак хордовых животных. Эмбриональное развитие тунпкат также имеет много общих черт с развитием Chordata.
В настоящее время считают, что оболочники путем вторичного упрощения, или деградации, произошли от каких-то форм, очень близких с позвоночными животными.
Вместе с другими хордовыми и иглокожими они образуют ствол вторичноротых животных - один из двух основных стволов эволюционного древа.
Оболочники рассматриваются или как отдельный подтип типа хордовых животных - Chordata, куда вместе с ними относятся еще три подтипа животных, в том числе и позвоночные (Vertebrata), или же как самостоятельный тип -Tunicata, или Urochordata. К этому типу относятся три класса: Аппендикулярии (Appendiculariae, или Copelata), Асцидии (Ascidiae) и Сальпы (Salpae).
Прежде асцидий делили на три отряда: простые, или одиночные, асцидий (Monascidiae); сложные, или колониальные, асцидий (Synascidiae) и пиросомы, или огнетелки (Ascidiae Salpaeformes, или Pyrosomata). Однако в настоящее время деление на простых и сложных асцидий потеряло систематическое значение. Асцидий делятся на подклассы по иным признакам.
Сальпы делятся на два отряда - бочоночников (Cyclomyaria) и собственно салъп (Desmomyaria). Иногда этим отрядам придают значение подклассов. К сальпам же, видимо, относится и очень своеобразное семейство глубоководных донных оболочников - Octacnemidae, хотя до сих пор большинство авторов считало его сильно уклонившимся подклассом асцидий.
Очень часто сальпы и пиросомы, ведущие свободноплавающий образ жизни, объединяются в группу пелагических оболочников Thaliacea, которой придается значение класса. Класс Thaliacea делят тогда на три подкласса: Pyrosomida, или Luciae, Desmomyaria, или Salpae, и Cyclomyaria, или Doliolida. Как видно, взгляды на систематику высших групп Tunicata весьма различны.
В настоящее время известно более тысячи видов оболочников. Громадное большинство их падает на долю асцидий, аппендикулярий насчитывается около 60 видов, около 25 видов сальп и примерно 10 видов пиросом (табл. 28-29).
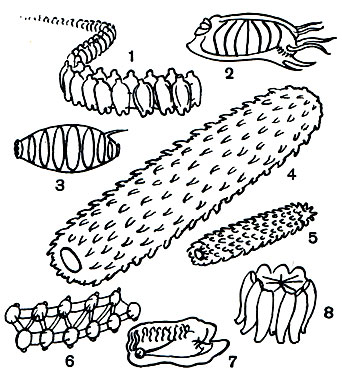
Таблица 28. Пелагические оболочники: 1 - колония сальп Thalia sp.; 2 - Salpa maxima (агрегатная форма); 3 - бочоночник Doliolum rarum; 4, 5 - Pyrosoma sp.; 6 - Thalia democratica; 7 - колония сальп Gyclosalpa bakeri; 8 - Salpa maxima (одиночная форма)
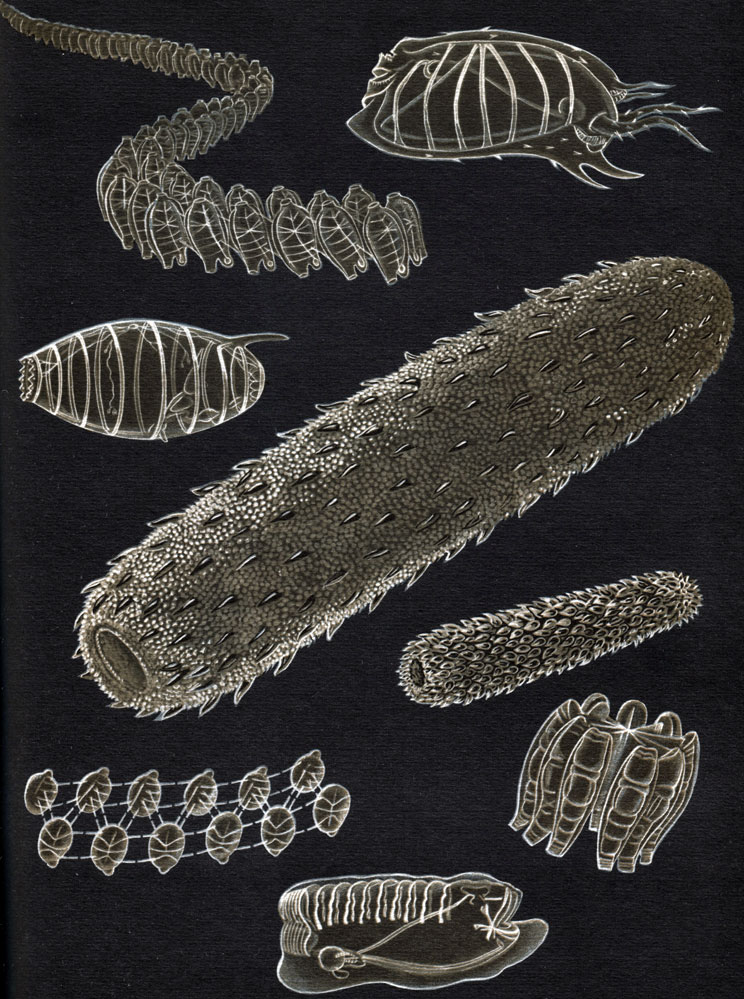
Таблица 28. Пелагические оболочники: 1 - колония сальп Thalia sp.; 2 - Salpa maxima (агрегатная форма); 3 - бочоночник Doliolum rarum; 4, 5 - Pyrosoma sp.; 6 - Thalia democratica; 7 - колония сальп Gyclosalpa bakeri; 8 - Salpa maxima (одиночная форма)
Как уже говорилось, оболочники обитают только в море. Аппендикулярип, сальпы и пиросомы плавают в толще вод океана, в то время как асцидий ведут прикрепленный образ жизни на дне. Аппендикулярий никогда не образуют колоний, в то время как сальпы и асцидий могут встречаться и в виде одиночных организмов, и в виде колоний. Пиросомы же всегда колониальны. Все оболочники - активные фильтраторы, питающиеся или микроскопическими пелагическими водорослями и животными, или взвешенными в воде частицами органического вещества - детритом. Прогоняя воду через глотку и жабры наружу, они отфильтровывают мельчайший планктон, иногда пользуясь при этом очень сложными приспособлениями.
Пелагические оболочники обитают в основном в верхних 200 м воды, но иногда могут опускаться и глубже. Пиросомы и сальпы редко встречаются глубже 1000 м, аппендикулярии известны до 3000 м. При этом специальные глубоководные виды среди них, видимо, отсутствуют. Асцидии в основной своей массе также распределены в приливо-отливной литоральной и в сублиторальной зонах океанов и морей - до 200-500 м, однако значительное количество их видов встречается и глубже. Максимальная глубина их нахождения - 7230 м.
Оболочники встречаются в океане то единичными экземплярами, то в виде колоссальных скоплений. Последнее особенно характерно для пелагических форм. В целом оболочники довольно обычны в морской фауне и, как правило, попадаются в планктонные сети и донные тралы зоологов повсеместно. Аппендикулярии и асцидии распространены в Мировом океане на всех широтах. Они столь же характерны для морей Северного Ледовитого океана и Антарктики, как и для тропиков. Сальпы и пиросомы, наоборот, в основном приурочены в своем распределении к теплым водам и лишь изредка встречаются в водах высоких широт, главным образом будучи занесенными туда теплыми течениями.
Строение тела почти всех туникат до неузнаваемости сильно отличается от общего плана строения тела в типе хордовых. Ближе всего к исходным формам стоят аппендикулярии, и в системе туникат они занимают первое место. Однако, несмотря на это, строение их тела является наименее характерным для оболочников. Знакомство с оболочниками, видимо, лучше всего начать с асцидий.
Строение асцидий. Асцидии - это донные животные, ведущие прикрепленный образ жизни. Многие из них одиночные формы. Размеры их тела в среднем составляют несколько сантиметров в диаметре и столько же в высоту. Однако среди них известны некоторые виды, достигающие 40-50 см, например широко распространенная Cione intestinalis или глубоководная Ascopera gigantea. С другой стороны, есть очень мелкие асцидии, размером менее 1 мм. Кроме одиночных асцидий, существует большое количество колониальных форм, у которых отдельные мелкие особи, размером в несколько миллиметров, погружены в общую тунику. Такие колонии, очень разнообразные по форме, обрастают поверхности камней и подводных предметов.
Больше всего одиночные асцидии похожи на продолговатый раздутый мешок неправильной формы, прирастающий своей нижней частью, которая называется подошвой, к различным твердым предметам (рис. 173, А). На верхней части животного хорошо заметны два отверстия, расположенные или на небольших бугорках, или же на довольно длинных выростах тела, напоминающих горлышко бутылки. Это сифоны. Один из них - ротовой, через который асцидия всасывает воду, второй - клоакальный. Последний обычно бывает несколько сдвинут на спинную сторону. Сифоны могут открываться и закрываться при помощи мускулов - сфинктеров. Тело асцидий одето однослойным клеточным покровом - эпителием, который выделяет на своей поверхности особую толстую оболочку - тунику. Внешняя окраска туники бывает различна. Обычно асцидии окрашены в оранжевые, красноватые, буро-коричневые или фиолетовые тона. Однако глубоководные асцидии, как и многие другие глубоководные животные, теряют свою окраску и становятся грязно-белыми. Иногда туника бывает полупрозрачной и через нее просвечивают внутренности животного. Часто туника образует по поверхности морщины и складки, зарастает водорослями, гидроидами, мшанками и другими сидячими животными. У многих видов ее поверхность покрыта песчинками и мелкими камешками, так что животное бывает трудно отличить от окружающих предметов.
Туника бывает студенистой, хрящевидной или желеобразной консистенции. Замечательной ее особенностью является то, что она более чем на 60% состоит из целлюлозы. Толщина стенок туники может достигать 2-3 см, обычно же она значительно тоньше.
Часть клеток эпидермиса может проникать в толщу туники и заселять ее. Это оказывается возможным только благодаря ее студенистой консистенции. Ни в одной другой группе животных клетки не заселяют образований аналогичного типа (например, кутикула у нематод). Кроме того, в толщу туники могут врастать и кровеносные сосуды.
Под туникой залегает собственно стенка тела, или мантия, в состав которой входит однослойный эктодермический эпителий, покрывающий тело, и соединительнотканый слой с мышечными волокнами. Наружные мышцы состоят из продольных, а внутренние из кольцевых волокон. Такая мускулатура позволяет асцидиям производить сократительные движения и при необходимости выбрасывать воду из тела. Мантия покрывает тело под туникой, так что оно свободно лежит внутри туники и срастается с нею только в области сифонов. В этих местах располагаются сфинктеры - мышцы, замыкающие отверстия сифонов.
Твердого скелета в теле асцидий нет. Только у некоторых из них в различных частях тела разбросаны мелкие известковые спикулы разной формы.
Пищеварительный канал асцидий начинается ртом, расположенным на свободном конце тела на вводном, или ротовом, сифоне (рис. 173, Б). Вокруг рта располагается венчик щупалец, иногда простых, иногда довольно сильно разветвленных. Количество и форма щупалец различны у разных видов, однако их не бывает менее 6. Ото рта свешивается внутрь огромная глотка, занимающая почти все пространство внутри мантии. Глотка асцидий образует сложный дыхательный аппарат. По стенкам ее в строгом порядке в несколько вертикальных и горизонтальных рядов располагаются жаберные щели, иногда прямые, иногда изогнутые (рис. 173, В). Часто стенки глотки образуют по 8-12 довольно больших свисающих внутрь складок, расположенных симметрично на двух ее сторонах и сильно увеличивающих ее внутреннюю поверхность. Складки также пронизаны жаберными щелями, причем сами щели могут принимать очень сложные очертания, закручиваясь спиралями на конусовидных выростах на стенках глотки и складок. Жаберные щели покрыты клетками, несущими длинные реснички. В промежутках между рядами жаберных щелей проходят, также правильно располагаясь, кровеносные сосуды. Количество их может достигать 50 на каждой стороне глотки. Здесь кровь обогащается кислородом. Иногда тонкие стенки глотки содержат небольшие спикулы, поддерживающие их.
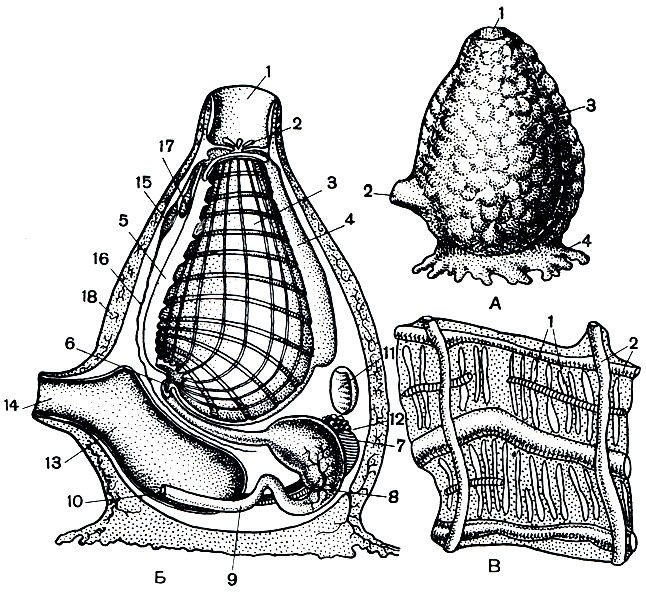
Рис. 173. Строение асцидий. А - внешний вид асцидий: 1 - вводной, или ротовой,, сифон; 2 - выводной, или клоакальный, сифон; 3 - туника; 4 - подошва. Б - схема строения одиночной асцидий: 1 - ротовой сифон; 2 - ротовые щупальца; 3 - глотка с жабрами; 4 - эндостиль; 5 - спинная пластинка; 6 - начало пищевода; 7 - желудок; 8 - пищеварительная железа; 9 - кишка; 10 - задний проход; 11 - сердце; 12 - яичник и семенник; 13 - клоака; 14 - клоакальный сифон; 15 - нервный ганглий; 16 - спинной нерв; 17 - околонервная железа; 18 - туника. В - часть стенки глотки с жаберными щелями: 1 - жаберные щели; 2 - продольные и поперечные кровеносные сосуды
Жаберные щели, или стигмы, асцидий незаметны, если рассматривать животное снаружи, сняв только тунику. Из глотки они ведут в особую полость, выстланную энтодермой и состоящую из двух половин, сросшихся на брюшной стороне с мантией. Эта полость называется околожаберной, атриальной или перибранхиальной (рис. 173, Б). Она залегает с каждой стороны между глоткой и наружной стенкой тела. Часть ее образует клоаку. Эта полость не является полостью тела животного. Она развивается из особых впячиваний наружной поверхности внутрь тела. Околожаберная полость сообщается с внешней средой при помощи клоакального сифона.
Со спинной стороны глотки свисает тонкая спинная пластинка, иногда рассеченная на тонкие язычки, а по брюшной стороне проходит особый поджаберный желобок, или эндостиль. Биением ресничек на стигмах асцидия гонит воду так, что устанавливается постоянный ток через ротовое отверстие. Далее вода прогоняется через жаберные щели в околожаберную полость и оттуда через клоаку наружу. Проходя через щели, вода отдает кислород в кровь, а различные мелкие органические остатки, одноклеточные водоросли и пр. захватываются эндостилем и гонятся по дну глотки к ее заднему концу. Здесь расположено отверстие, ведущее в короткий и узкий пищевод. Загибаясь на брюшную сторону, пищевод переходит во вздутый желудок, из которого выходит кишка. Кишка, изгибаясь, образует двойную петлю и открывается анальным отверстием в клоаку. Экскременты выталкиваются из тела через клоакальный сифон. Таким образом, пищеварительная система асцидий очень проста, но обращает на себя внимание наличие эндостиля, являющегося частью их ловчего аппарата. Клетки эндостиля двух родов - железистые и реснитчатые. Реснитчатые клетки эндостиля улавливают пищевые частицы и гонят их к глотке, склеивая выделениями железистых клеток. Оказывается, эндостиль является гомологом щитовидной железы позвоночных и выделяет органическое вещество, содержащее йод. Видимо, это вещество близко по своему составу к гормону щитовидной железы. У некоторых асцидий у основания стенок желудка имеются особые складчатые выросты и дольчатые массы. Это так называемая печень. Она соединяется с желудком особым протоком.
Кровеносная система асцидий незамкнутая. Сердце расположено на брюшной стороне тела животного. Оно имеет вид небольшой вытянутой трубки, окруженной тонкой околосердечной сумкой, или перикардием. От двух противоположных концов сердца отходит по крупному кровеносному сосуду. От переднего конца начинается жаберная артерия, которая тянется посередине брюшной стороны и посылает от себя многочисленные ветви к жаберным щелям, дающие между ними побочные маленькие веточки и окружающие жаберный мешок целой сетью продольных и поперечных кровеносных сосудов. От задней спинной стороны сердца отходит кишечная артерия, дающая ветви к внутренним органам. Здесь кровеносные сосуды образуют широкие лакуны-пространства между органами, не имеющие собственных стенок, очень похожие по своему строению на лакуны двустворчатых моллюсков. Кровеносные сосуды заходят также в стенку тела и даже в тунику.
Вся система кровеносных сосудов и лакун открывается в жаберно-кишечный синус, иногда называемый спинным сосудом, с которым соединяются и спинные концы поперечных жаберных сосудов. Этот синус значителен по размерам и тянется посередине спинной части глотки. Всем туникатам, в том числе и асцидиям, свойственна периодическая смена направления тока крови, так как их сердце поочередно сокращается в течение некоторого времени то сзади наперед, то спереди назад. Когда сердце сокращается от спинного отдела к брюшному, кровь движется по жаберной артерии к глотке, или жаберному мешку, где она окисляется и откуда поступает в кишечно-жаберный синус. Затем кровь проталкивается в кишечные сосуды и снова к сердцу, так же, как это имеет место у всех позвоночных животных. При последующем же сокращении сердца направление тока крови меняется на обратное, и она течет, как у большинства беспозвоночных. Таким образом, тип кровообращения у ту ни кат является переходным между кровообращением беспозвоночных и позвоночных животных. Кровь у асцидий бесцветная, кислая. Замечательной ее особенностью является наличие ванадия, который принимает участие в переносе кислорода кровью и заменяет железо.
Нервная система у взрослых асцидий чрезвычайно проста и развита значительно слабее, чем у личинки. Упрощение нервной системы происходит благодаря сидячему образу жизни взрослых форм. Нервная система состоит из надглоточного, или мозгового, ганглия, расположенного на спинной стороне тела между сифонами. От ганглия берут начало 2-5 пар нервов, идущих к краям ротового отверстия, глотке и к внутренностям - кишечнику, половым органам и к сердцу, где имеется нервное сплетение. Между ганглием и спинной стенкой глотки имеется небольшая околонервная железка, проток которой впадает в глотку на дне ямки в особом мерцательном органе. Эту железку считают иногда гомологом нижнего придатка головного мозга позвоночных животных - гипофиза. Органы чувств отсутствуют, но, вероятно, ротовые щупальца имеют осязательную функцию. Но тем не менее нервная система оболочников в сущности не примитивна. Личинки асцидий имеют спинномозговую трубку, лежащую под хордой и образующую вздутие на своем переднем конце. Это вздутие, по-видимому, соответствует головному мозгу позвоночных животных и содержит личиночные органы чувств - пигментированные глазки и орган равновесия, или статоцист. Когда личинка превращается во взрослое животное, вся задняя часть нервной трубки исчезает, а мозговой пузырь вместе с личиночными органами чувств распадается; за счет его спинной стенки и образуется спинной ганглий взрослой асцидий, а брюшная стенка пузыря образует околонервную железу. Как отмечает В. Н. Беклемишев, строение нервной системы оболочников является одним из лучших доказательств происхождения их от высокоорганизованных подвижных животных. Нервная система личинок асцидий по своему развитию выше, чем нервная система ланцетника, у которого отсутствует мозговой пузырь.
Специальных выделительных органов у асцидий нет. Вероятно, в какой-то степени в выделении принимают участие стенки пищеварительного канала. Однако у многих асцидий имеются особые так называемые рассеянные почки накопления, состоящие из особых клеток - нефроцитов, в которых накапливаются продукты выделения. Эти клетки располагаются характерным рисунком, часто группируются вокруг кишечной петли или гонад. Красновато-бурая окраска многих асцидий зависит именно от накопившихся в клетках экскретов. Только после смерти животного и распада тела продукты выделения освобождаются и выходят в воду. Иногда во втором колене кишки располагается скопление прозрачных пузырьков, не имеющих выводных протоков, в которых накапливаются конкреции, содержащие мочевую кислоту. У представителей семейства Molgulidae почка накопления усложняется еще больше и скопление пузырьков превращается в один крупный изолированный мешочек, полость которого содержит конкреции. Большое своеобразие этого органа заключается в том, что почечный мешок мольгулид у некоторых других асцидий всегда содержит симбиотические грибки, которые не имеют даже отдаленных родственников среди других групп низших грибов. Грибки образуют тончайшие нити мицеллы, оплетающие конкреции. Среди них присутствуют более толстые образования неправильной формы, иногда образуются спорангии со спорами. Эти низшие грибы питаются уратами - продуктами выделения асцидий, и их развитие освобождает последних от накопленных экскретов. Видимо, эти грибки необходимы асцидиям, так как даже ритм размножения у некоторых форм асцидий связан с накоплением экскретов в почках и с развитием симбиотических грибков. Как происходит перенос грибков от одной особи к другой, неизвестно. Яйца асцидий стерильны в этом отношении, а молодые личинки не содержат в почке грибков, даже когда экскреты уже накапливаются в них. Видимо, молодые животные вновь "заражаются" грибками из морской воды. Асцидии - гермафродиты, т. е. у одной и той же особи имеются одновременно и мужские и женские половые железы. Яичники и семенники лежат по одному или по нескольку пар с каждой стороны тела, обычно в петле кишечника. Протоки их открываются в клоаку, так что клоакальное отверстие служит не только для выхода воды и экскрементов, но и для выведения половых продуктов. Самооплодотворения у асцидии не происходит, так как яйца и сперма созревают в разное время. Оплодотворение чаще всего происходит в околожаберной полости, куда с током воды проникают сперматозоиды другой особи. Реже оно бывает снаружи. Оплодотворенные яйца выходят через клоакальный сифон, но иногда яйца развиваются в околожаберной полости и наружу выходят уже сформированные плавающие личинки. Такое живорождение характерно особенно для колониальных асцидии.
Кроме полового размножения, асцидиям свойствен и бесполый способ размножения путем почкования. При этом образуются разнообразные колонии асцидии.
Строение асцидиозооида - члена колонии сложных асцидии - в принципе не отличается от строения одиночной формы. Но размеры их много меньше и обычно не превышают нескольких миллиметров. Тело асцидиозооида вытянуто в длину и разделено на два или три отдела (рис. 174, А): в первом, грудном, отделе располагается глотка, во втором - кишечник, а в третьем - половые железы и сердце. Иногда разные органы располагаются несколько иначе.
Степень связи между отдельными особями в колонии-асцидиозооидами-может быть различной. Иногда они вполне самостоятельны и соединяются только тонким столоном, который стелется по грунту. В других случаях асцидиозооиды заключены в общую тунику. Они могут быть или рассеяны в ней, и тогда наружу выходят и ротовые и клоакальные отверстия асцидиозооидов, или располагаются правильными фигурами в виде колец или эллипсов (рис. 174, Б). В последнем случае колония состоит из групп особей, обладающих самостоятельными ртами, но имеющих общую клоакальную полость с одним общим клоакальным отверстием, в которую открываются клоаки отдельных особей. Как уже указывалось, размеры таких асцидиозооидов составляют всего лишь несколько миллиметров. В случае же, когда связь между ними осуществляется только при помощи столона, асцидиозооиды достигают более крупных размеров, но обычно меньших, чем одиночные асцидии.
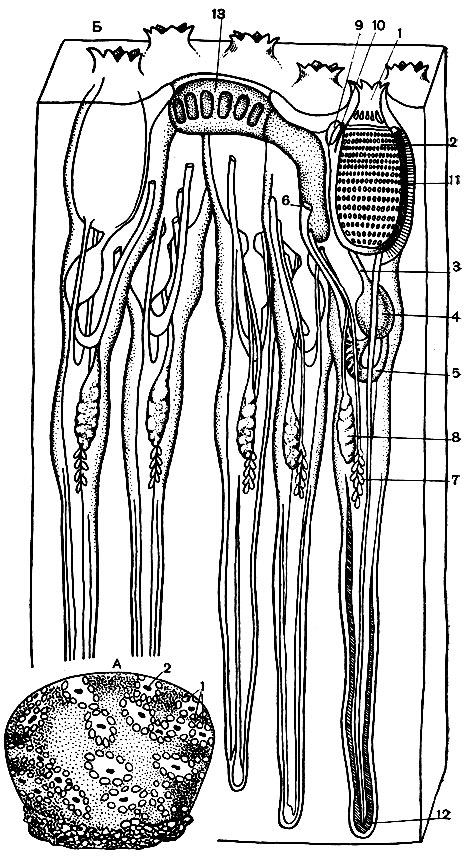
Рис. 174. Строение сложных асцидии. А - внешний вид колонии: 1 - ротовые сифоны отдельных особей; 2 - выход общей клоакальной полости. Б - схема строения сложной асцидии: 1 - ротовой сифон; 2 - глотка; 3 - пищевод; 4 - желудок; 5 - кишка; 6 - задний проход; 7 - семенник; 8 - яичник; 9 - нервный ганглий; 10 - околонервная железка; 11 - эндостиль; 12 - сердце с выростами перикардия; 13 - общая клоака с отверстиями клоакальных сифонов отдельных зооидов
Развитие асцидий, их бесполое и половое размножение будут описаны ниже.
Строение пиросом. Пиросомы, или огнетелки,- это свободноплавающие колониальные пелагические оболочники. Свое название они получили из-за способности светиться ярким фосфорическим светом.
Из всех планктонных форм туникат они ближе всего стоят к асцидиям. По существу, это плавающие в воде колониальные асцидии. Каждая колония состоит из многих сотен отдельных особей - асцидиозооидов, заключенных в общую, часто очень- плотную тунику (рис. 175, А). У пиросом все зооиды равноценны и независимы в отношении питания и размножения. Колония образуется путем почкования отдельных особей, причем почки попадают на свое место, передвигаясь в толще туники с помощью особых странствующих клеток - фороцитов. Колония имеет форму длинного вытянутого цилиндра с заостренным концом, имеющего внутри полость и открытого на своем широком заднем конце (рис. 175, Б). Снаружи пиросома покрыта небольшими мягкими шиловидными выростами. Важнейшее отличие их от колоний сидячих асцидий заключается еще и в строгой геометрической правильности формы колонии. Отдельные зооиды стоят перпендикулярно к стенке конуса. Их ротовые отверстия обращены наружу, а клоакальные находятся на противоположной стороне тела и открываются внутрь полости конуса. Отдельные маленькие асцидиозооиды захватывают своими ртами воду, которая, пройдя через их тело, попадает в полость конуса. Движения отдельных особей согласованы между собой, причем эта согласованность движений возникает механическим путем при отсутствии мышечных, сосудистых или нервных связей. В тунике пиросом от одной особи к другой протянуты механические волокна, соединяющие их двигательные мышцы. Сокращение мышцы одной особи дергает при помощи волокон туники другую особь и передает ей раздражение. Сокращаясь одновременно, маленькие зооиды проталкивают воду через полость колонии. При этом вся колония, похожая по форме на ракету, получив обратный толчок, движется вперед. Таким образом, пиросомы избрали для себя принцип реактивного движения. Такой способ движения используется не только пиросомами, но и другими пелагическими туникатами.
Туника пиросом содержит такое большое количество воды (у некоторых оболочников вода составляет 99% от веса тела), что вся колония становится прозрачной, как будто стеклянной, и почти невидима в воде. Однако встречаются и окрашенные в розовый цвет колонии. Такие пиросомы гигантских размеров - длина их достигает 2,5 и даже 4 м, а диаметр колонии 20-30 см - неоднократно ловились в Индийском океане. Их название - Pyrosoma spinosum. Туника этих пиросом обладает настолько нежной консистенцией, что, попадая в планктонные сети, колонии обычно распадаются на отдельные куски. Обычно же размеры пиросом значительно меньше - от 3 до 10 см длиной с диаметром от одного до нескольких сантиметров. Недавно описан новый вид пиросом - P. vitjasi. Колония этого вида также имеет цилиндрическую форму и размеры до 47 см. По описанию автора, сквозь розоватую мантию как темно-коричневые (вернее, темно-розовые у живых экземпляров) включения просвечивают внутренности отдельных асцидиозооидов. Мантия имеет полужидкую консистенцию, и при повреждении поверхностного слоя вещество ее растекается в воде в виде тягучей слизи, а отдельные зооиды свободно распадаются.
Строение асцидиозооида пиросом мало чем отличается от строения одиночной асцидии, за исключением того, что сифоны его располагаются на противоположных сторонах тела, а не сближены на спинной стороне (рис. 175, В). Размеры асцидиозооидов обычно составляют 3-4 мм, а у гигантских пиросом - до 18 мм длины. Тело их может быть сплющенным с боков или овальным. Ротовое отверстие окружено венчиком щупалец, или же может присутствовать только одно щупальце с брюшной стороны тела. Часто мантия перед ротовым отверстием, также с брюшной стороны, образует по небольшому бугорку или же довольно значительному выросту. За ртом следует крупная глотка, прорезанная жаберными щелями, количество которых может достигать 50. Эти щели располагаются или вдоль, или поперек глотки. Приблизительно перпендикулярно к жаберным щелям проходят кровеносные сосуды, число которых также варьирует от одного до трех-четырех десятков. В глотке имеются эндостиль и спинные язычки, свешивающиеся в ее полость. Кроме того, в передней части глотки по бокам расположены светящиеся органы, представляющие собой скопления клеточных масс. У некоторых видов светящиеся органы есть еще и у клоакального сифона. Органы свечения пиросом заселены симбиотическими светящимися бактериями. Под глоткой лежит нервный ганглий, имеется и околонервная железка, канал которой открывается в глотку. Мышечная система асцидиозооидов пиросом развита слабо. Имеются довольно хорошо выраженные кольцевые мышцы, расположенные вокруг ротового сифона, и незамкнутое кольцо мышц у клоакального сифона. Небольшие пучки мышц - спинной и брюшной - располагаются в соответствующих местах глотки и лучеобразно расходятся по бокам тела. Кроме того, есть еще пара клоакальных мышц. Между спинной частью глотки и стенкой тела имеются два кроветворных органа, представляющих собой продолговатые скопления клеток. Размножаясь путем деления, эти клетки превращаются в различные элементы крови - лимфоциты, амебоциты и пр.
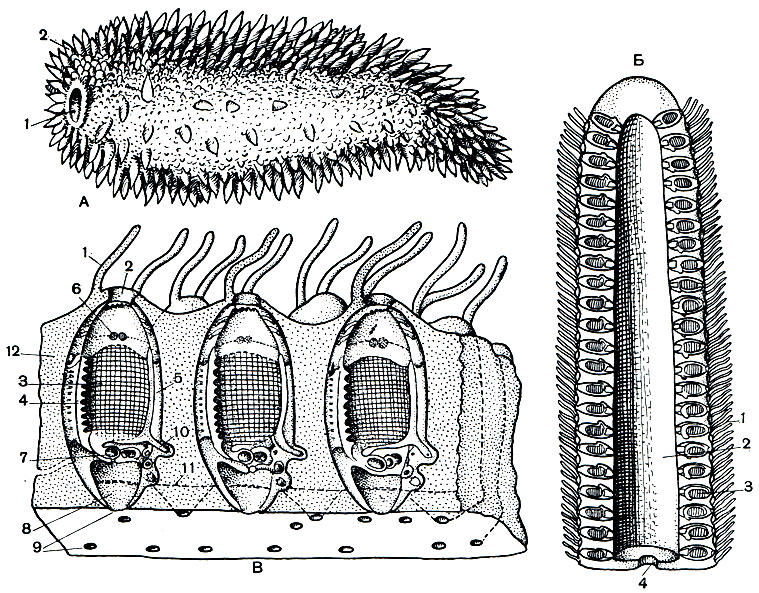
Рис. 175. Строение пиросом. А - внешний вид колонии: 1 - отверстие полости колонии; 2 - околоротовые выросты туники. Б - продольный разрез колонии: 1 - околоротовые выросты туники; 2 - полость колонии; 3 - зооиды; 4 - отверстие полости колонии. В - строение зооида пиросом: 1 - околоротовой вырост туники; 2 - ротовой сифон; 3 - глотка с жаберными щелями; 4 - спинная пластинка; 5 - эндостиль; 6 - светящиеся органы; 7 - кишечник; 8 - клоака; 9 - клоакальные отверстия; 10 - столон; 11 - волокнистые тяжи туники; 12 - общеколониальная туника
Пищеварительный отдел кишечника состоит из пищевода, отходящего от задней части глотки, желудка и кишки. Кишка образует петлю и открывается анальным отверстием в клоаку. На брюшной стороне тела лежит сердце, представляющее собой тонкостенный мешочек. Имеются семенники и яичники, протоки которых также открываются в клоаку, которая может быть более или менее вытянута и открывается клоакальным сифоном в общую полость колонии. В области сердца у асцидиозооидов пиросом имеется маленький пальцевидный придаток - столон. Он играет важную роль при формировании колонии. В результате деления столона в процессе бесполого размножения от него отпочковываются новые особи.
Строение сальп. Так же как и пиросомы, сальпы являются свободноплавающими животными и ведут пелагический образ жизни. Они делятся на два отряда: бочоночников, или долиолид (Gyclomyaria), и собственно сальп (Desmomyaria). Это совершенно прозрачные животные в форме бочонка или огурца, на противоположных концах которого расположены ротовое и анальное отверстия - сифоны. Только у некоторых видов сальп отдельные части тела, например столон и кишечник, бывают окрашены у живых экземпляров в голубовато-синий цвет. Тело их одето нежной прозрачной туникой, иногда снабженной выростами разной длины. Сквозь стенки тела хорошо просвечивает маленький, обычно зеленовато-бурый кишечник. Размеры сальп колеблются от нескольких миллиметров до нескольких сантиметров в длину. Самая крупная сальпа - "Thetys vagina - была поймана в Тихом океане. Длина ее тела (вместе с придатками) составляла 33,3 см.
Одни и те же виды сальп встречаются или одиночными формами, или в виде длинных цепочкообразных колоний. Такие цепочки сальп представляют собой соединенные друг с другом в ряд отдельные особи. Связь между зооидами в колонии сальп, как анатомическая, так и физиологическая, чрезвычайно слаба. Члены цепочки как бы слипаются друг с другом прикрепительными сосочками, и по существу колониальность их и зависимость друг от друга едва выражены. Такие цепочки могут достигать длины более одного метра, но они легко разрываются на части, иногда просто при ударе волны. Отдельные особи и особи, являющиеся членами цепочки, настолько сильно отличаются друг от друга и по величине и по внешнему виду, что даже описывались старыми авторами под разными видовыми названиями.
Представители же другого отряда - бочоночники, или долиолиды, - наоборот, строят чрезвычайно сложные колонии. Один из крупнейших современных зоологов В. Н. Беклемишев назвал бочоночников одними из самых фантастических существ в море. В отличие от асцидий, у которых образование колоний происходит благодаря почкованию, возникновение колоний у всех сальп строго связано с чередованием поколений. Одиночные сальпы - это не что иное, как появившиеся из яиц бесполые особи, которые, почкуясь, дают начало колониальному поколению.
Как уже говорилось, тело отдельной особи, будь она или одиночной, или членом колонии, одето тонкой прозрачной туникой. Под туникой подобно обручам бочонка просвечивают белесоватые ленты кольцевых мышц. Таких колец у них имеется 8. Они опоясывают тело животного на определенном расстоянии друг от друга. У бочоночников мышечные ленты образуют замкнутые обручи, а у собственно сальп они не смыкаются на брюшной стороне. Последовательно сокращаясь, мышцы проталкивают поступающую через рот воду по организму животного и выталкивают ее через выводной сифон. Как и пиросомы, все сальпы движутся, используя реактивный способ движения.
В отряде долиолид бочонки широко открыты на обоих концах (рис. 176). На одном конце располагается ротовое отверстие, на противоположном - анальное. Оба отверстия окружены чувствительными бугорками. Внутренность бочонка поделена косой перегородкой или спинным выростом на две полости. Передняя полость - глотка, задняя - клоака. Рот ведет прямо в огромную глотку, занимающую почти весь объем тела. В отличие от асцидий боковые стенки глотки бочоночников сплошные и только задняя стенка, которая отгораживает полость глотки от клоаки, пронизана двумя сходящимися рядами жаберных щелей. Щели соединяют глотку прямо с клоакой, а особые околожаберные полости, которые имеются у асцидий, здесь отсутствуют. От них остается только одна клоакальная полость. На дне глотки имеется эндостиль, а по спинной стороне, как и у других рассмотренных нами туникат, проходит продольный вырост - спинная пластинка. Эндостиль ведет из глотки в кишечник, очень укороченный, расположенный на брюшной части перегородки между двумя полостями. Кишечник состоит из короткого пищевода, переходящего в колбообразный желудок, к спинкам которого примыкает пищеварительная железа, и кишки. Кишка открывается анальным отверстием в клоаку.
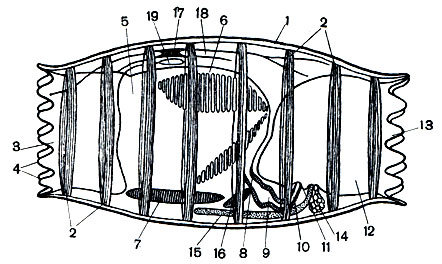
Рис. 176. Строение бочоночника: 1 - туника; 2 - мышечные кольца; 3 - ротовой сифон; 4 - щупальца; 5 - глотка; 6 - жаберные щели; 7 - эндостиль; 8 - пищевод; 9 - желудок; 10 - кишка; 11 - задний проход; 12 - клоака; 13 - клоакальный сифон со щупальцами; 14 - яичник; 15 - семенник; 16 - сердце; 17 - нервный ганглий; 18 - нервы; 19 - околонервная железка
Нервная система состоит из расположенного над глоткой мозгового ганглия, от которого отходят нервы. Рядом с желудком лежит сердечный мешок. От сердца отходят кровеносные сосуды, которые, как и у всех оболочников, образуют незамкнутые лакуны, расположенные неправильной сетью.
Как и все оболочники, бочоночники- гермафродиты. У них имеется один яичник и один семенник. Половые железы лежат по одну сторону от желудка и также открываются протоками в клоакальную полость. В яичнике развивается одновременно только по одному крупному яйцу.
Органы выделения отсутствуют. Вероятно, их функцию выполняют некоторые клетки крови, в которых найдены желтовато-коричневые конкреции. Током крови эти конкреции переносятся в область желудка, где они концентрируются, проникают затем в кишку и выбрасываются вон из организма. У некоторых сальп, например у Gyclosalpa, найдены скопления ампул ОБИДНЫХ клеток, очень похожих на таковые у асцидий. Они располагаются также в области кишечника и, видимо, играют роль почек накопления. Однако это еще не установлено окончательно.
Только что описанное строение тела относится к половому поколению бочоночников. Бесполые же особи не имеют половых гонад. Для них характерно наличие двух столонов. Один из них, почкородный, как и у пиросом, располагается на брюшной стороне тела и называется брюшным столоном; второй столон-спинной.
Собственно сальпы по своему строению очень похожи на бочоночников и отличаются от них только в деталях (рис. 177, А, Б). По внешнему виду это также прозрачные цилиндрообразные животные, сквозь стенки тела которых хорошо просвечивает компактный, обычно оливкового цвета, желудок. Туника сальп может давать разнообразные выросты, иногда довольно длинные у колониальных форм. Как уже указывалось, мышечные обручи у них не замкнуты, а количество их может быть больше, чем у бочоночников. Кроме того, клоакальное отверстие несколько сдвинуто на спинную сторону, а не лежит прямо на заднем конце тела, как у бочоночников. Перегородка между глоткой и клоакой пронизана всего лишь двумя жаберными щелями, но зато эти щели огромны по размерам. И наконец, мозговой ганглий у сальп развит несколько сильнее, чем у бочоночников. У сальп он имеет сферическую форму с подковообразным вырезом на спинной стороне. Здесь помещается довольно сложно устроенный пигментированный глазок.
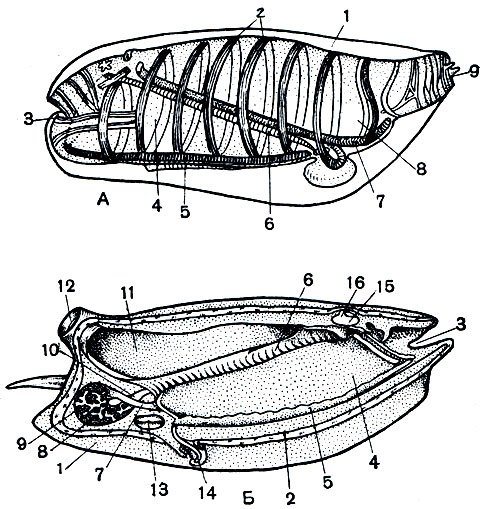
Рис. 177. Строение сальпы. А - внешний вид: 1 - туника; 2 - мышечные тяжи; 3 - ротовой сифон; 4 - глотка; 5 - эндостиль; 6 - жаберное отверстие; 7 - кишка; 8 - клоака; 9 - клоакальный сифон. Б - строение сальпы: 1 - туника; 2 - мантия; 3 - ротовой сифон; 4 - глотка; 5 - эндостиль; 6 - жабра; 7 - пищевод; 8 - желудок; 9 - пищеварительная железа; 10 - кишка; 11 - клоака; 12 - клоакальный сифон; 13 - сердце; 14 - столон; 15 - нервный ганглий; 16 - глаз
Сальпы и бочоночники обладают способностью светиться. Органы свечения у них очень похожи на органы свечения пиросом и представляют собой скопления клеток, расположенных на брюшной стороне в области кишечника и содержащих симбиотические светящиеся бактерии. Особенно сильно органы свечения развиты у видов рода Cyclosalpa, светящихся более интенсивно, чем другие виды. У них образуются так называемые "боковые органы", расположенные по бокам с каждой стороны тела.
Как уже неоднократно указывалось, сальпы являются типичными планктонными организмами. Однако существует одна очень небольшая группа своеобразных донных оболочников - Octacnemidae, насчитывающая всего четыре вида. Это бесцветные животные до 7 см в диаметре, обитающие на морском дне. Тело их покрыто тонкой полупрозрачной туникой, образующей вокруг ротового сифона восемь довольно длинных щупалец. Оно уплощено и по внешнему виду напоминает асцидий. Но по внутреннему строению октакнемиды близки к сальпам. В зоне прикрепления к субстрату туника дает тонкие волосовидные выросты, но, видимо, эти животные слабо укреплены в грунте и могут плавать над дном на небольшие расстояния. Некоторые ученые считают их особым, сильно уклонившимся подклассом асцидий, другие же склонны рассматривать их как вторично осевших на дно сальп. Octacnemidae - глубоководные животные, найденные в тропических районах Тихого океана и у берегов Патагонии, а также в Атлантическом океане южнее Гренландии, главным образом на глубине 2000-4000 тыс. м.
Строение аппендикулярий. Аппендикудярии - очень мелкие прозрачные свободноплавающие животные. В отличие от остальных оболочников они никогда не образуют колоний. Размеры их тела колеблются от 0,3 до 2,5 см. Личинки аппендикулярий не подвергаются в своем развитии регрессивному метаморфозу, т. е. упрощению строения тела и утрате целого ряда важных органов, например хорды и органов чувств, вызванному превращением свободноплавающей личинки в неподвижную взрослую форму, как это имеет место у асцидий. Взрослая аппендикулярия по своему строению очень похожа на личинку асцидий. Как уже говорилось, такая важная черта строения их тела, как наличие хорды, которая всех оболочников ставит в одну группу с хордовыми животными, сохраняется у аппендикулярий в течение всей жизни, и именно этим они отличаются от всех остальных оболочников, по внешнему виду абсолютно непохожих на своих ближайших родственников.
Тело аппендикулярий распадается на туловище и хвост (рис. 178, А). Общий вид животного напоминает головастика лягушек. Хвост, длина которого в несколько раз превышает длину округлого тельца животного, прикрепляется к брюшной стороне в виде длинной тонкой пластинки. Аппендикулярия держит его повернутым на 90° вокруг его длинной оси и подогнутым на брюшную сторону. Вдоль середины хвоста по всей его длине проходит хорда - эластичный тяж, состоящий из ряда крупных клеток. По бокам от хорды имеются 2 мышечные ленты, каждая из которых образована всего лишь десятком гигантских клеток.
На переднем конце туловища лежит рот, ведущий в объемистую глотку (рис. 178, Б). Глотка сообщается непосредственно с внешней средой двумя продолговатоовальными жаберными отверстиями, или стигмами. Околожаберной полости с клоакой, как у асцидий, здесь нет. По брюшной стороне глотки проходит эндостиль, на противоположной, спинной, стороне заметен продольный спинной вырост. Эндостиль гонит комочки пищи к пищеварительному отделу кишечника, который напоминает подковообразную изогнутую трубку и состоит из пищевода, короткого желудка и короткой задней кишки, открывающейся наружу анальным отверстием на брюшной стороне тела.
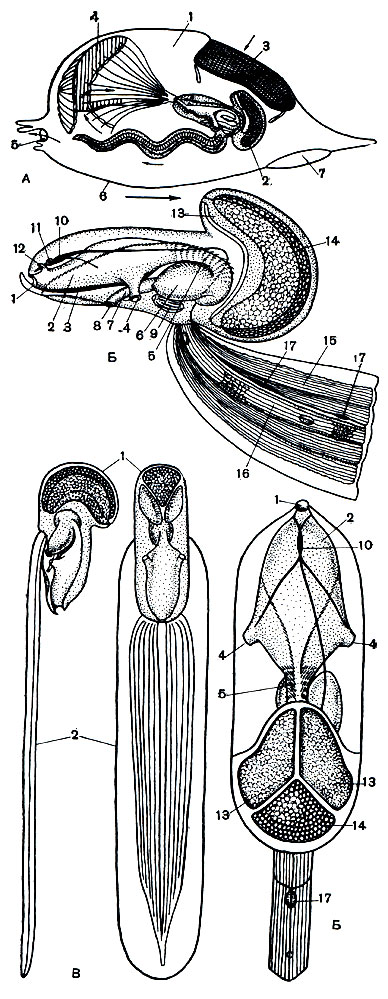
Рис. 178. Строение аппендикулярий. А - домик аппендикулярий: 1 - домик; 2 - аппендикулярия; 3 - фильтр у входного отверстия домика; 4 - воронкообразное приспособление для лова добычи; 5 - выходное отверстие домика; 6 - стенка домика; 7 - отверстие для бегства. Б - строение аппендикулярий (вид сбоку и со спинной стороны): 1 - рот; 2 - глотка; 3 - эндостиль; 4 - жаберное отверстие; 5 - пищевод; 6 - желудок; 7 - кишка; 8 - задний проход; 9 - сердце; 10 - нервный ганглий и спинной нерв; 11 - статоцист; 12 - мерцательная ямка; 13 - семенник; 14 - яичник с яйцами; 15 - мышцы хвоста; 16 - хорда; 17 - нервные ганглии хвоста. В - внешний вид животного без домика (вид сбоку и со спинной стороны): 1 - тельце; 2 - хвост. Стрелки показывают направление тока воды и движения домика
На брюшной стороне тела под желудком лежит сердце. Оно имеет форму продолговатоовального баллона, плотно прилегающего своей спинной стороной к желудку. К переднему отделу туловища от сердца идут кровеносные сосуды - брюшной и спинной. В переднем отделе глотки они соединяются при помощи кольцевого сосуда. Имеется система лакун, по которым, как и по кровеносным сосудам, идет циркуляция крови. Кроме того, по спинной и брюшной сторонам хвоста также проходит по кровеносному сосуду. Сердце аппендикулярий, как и у остальных оболочников, периодически меняет направление тока крови, сокращаясь в течение нескольких минут то в одном, то в другом направлении. При этом оно работает очень быстро, делая до 250 сокращений в минуту.
Нервная система состоит из крупного надглоточного мозгового ганглия, от него отходит назад спинной нервный ствол, достигающий конца хвоста и проходящий над хордой. У самого основания хвоста нервный ствол образует вздутие- небольшой нервный узелок. Несколько таких же нервных узелков, или ганглиев, имеется на протяжении всего хвоста. К спинной стороне мозгового ганглия тесно примыкает маленький орган равновесия - статоцист, а на спинной стороне глотки имеется небольшая ямка. Ее обычно принимают за орган обоняния. Других органов чувств у аппендикулярий нет. Специальные органы выделения отсутствуют.
Аппендикулярий - гермафродиты, у них имеются и женские и мужские половые органы. В задней части туловища располагается яичник, тесно сжатый с двух сторон семенниками. Сперматозоиды выводятся из семенников наружу через отверстия на спинной стороне тела, а яйца попадают в воду только после разрыва стенок тела. Таким образом, после откладки яиц аппендикулярий погибают.
Все аппендикулярий строят чрезвычайно характерные домики, являющиеся результатом выделения их кожного эпителия (рис. 178, В). Этот несколько заостренный спереди домик - толстостенный, студенистый и совершенно прозрачный - сначала вплотную прилегает к телу, а затем отстает от него так, что животное может свободно двигаться внутри домика. Домик и есть туника, но у аппендикулярий она не содержит целлюлозы, а состоит из хитина, вещества, близкого по строению к роговому. На переднем и заднем конце домик снабжен несколькими отверстиями. Находясь внутри, аппендикулярия производит хвостом волнообразные движения, благодаря чему внутри домика образуется ток воды, и вода, выходя из домика, заставляет его двигаться в противоположную сторону. На той же стороне домика, в которую он движется, наверху имеются два отверстия, затянутые очень частой решеткой с длинными узкими щелями. Ширина этих щелей 9-46 мк, а длина 65-127 мк. Решетка представляет собой фильтр для поступающих с водой внутрь домика пищевых частиц. Аппендикулярий питаются только тем мельчайшим планктоном, который проходит через отверстия решетки. Обычно это организмы размером 3-20 мк. Более же крупные частички, рачки, радиолярии и диатомовые водоросли, не могут проникнуть внутрь домика.
Ток воды, поступив внутрь домика, попадает в новую решетку, по форме напоминающую вершу и заканчивающуюся на конце мешковидным каналом, за который держится своим ртом аппендикулярия. Бактерии, мельчайшие жгутиковые, корненожки и другие организмы, прошедшие через первый фильтр, собираются на дне канала, и аппендикулярия питается ими, делая время от времени глотательные движения. Но тонкий передний фильтр быстро засоряется. У некоторых видов, например у Oikopleura rufescens, он перестает работать уже через 4 часа. Тогда аппендикулярия покидает испорченный домик и выделяет вместо него новый. На постройку нового домика уходит всего лишь около 1 часа, и снова она начинает отфильтровывать мельчайший наннопланктон. За время своей работы домик успевает пропустить примерно 100 см3 воды. Для того чтобы покинуть домик, аппендикулярия пользуется так называемой "калиткой для бегства". Стенка домика в одном месте очень сильно утончена и превращена в тонкую пленку. Пробив ее ударом хвоста, животное быстро покидает домик, для того чтобы тут же выстроить новый. Домик аппендикулярий очень легко разрушается при фиксации или при механическом воздействии, и его можно видеть только у живых организмов.
Характерной чертой аппендикулярии является постоянство клеточного состава, т. е. постоянство количества клеток, из которых строится все тело животного. Причем разные органы строятся также из определенного числа клеток. Такое же явление известно для коловраток и нематод. У коловраток, например, число клеточных ядер и особенно их расположение всегда постоянны для определенного вида. Один вид состоит из 900 клеток, другой - из 959. Это происходит в результате того, что каждый орган формируется из небольшого числа клеток, после чего размножение клеток в нем на всю жизнь останавливается. У нематод не все органы обладают постоянством клеточного состава, а только мышцы, нервная система, задняя кишка и некоторые другие. Количество клеток в них невелико, но зато размеры клеток могут быть огромными.
Размножение и развитие оболочников. Размножение оболочников представляет собой удивительный пример того, какие необычайно сложные и фантастические жизненные циклы могут существовать в природе. Всем оболочникам, кроме аппендикулярии, свойствен как половой, так и бесполый способ размножения. В первом случае новый организм образуется из оплодотворенного яйца. Но у оболочников при этом развитие до взрослой особи происходит с глубокими преобразованиями в строении личинки в сторону ее значительного упрощения. При бесполом же размножении новые организмы как бы отпочковываются от материнской особи, получая от нее зачатки всех основных органов.
Все половые особи оболочников - гермафродиты, т. е. они обладают как мужскими, так и женскими половыми железами. Созревание мужских и женских половых продуктов всегда происходит в разное время, и поэтому невозможно самооплодотворение. Мы уже знаем, что у асцидий, сальп и пиросом протоки половых желез открываются в клоакальную полость, а у аппендикулярии сперматозоиды выходят в воду через протоки, открывающиеся на спинной стороне тела, тогда как яйца могут выйти наружу только после разрыва стенок, что ведет к гибели животного. Оплодотворение у оболочников, кроме сальп и пиросом, наружное. Это значит, что сперматозоид встречается с яйцеклеткой в воде и там оплодотворяет ее. У сальп и пиросом образуется только по одному яйцу, которое оплодотворяется и развивается в теле матери. У некоторых асцидий оплодотворение яиц тоже происходит в клоакальной полости матери, куда с током воды через сифоны проникают сперматозоиды других особей, а оплодотворенные яйца выводятся через анальный сифон. Иногда же зародыши и развиваются в клоаке и лишь потом выходят наружу, т. е. имеет место своего рода живорождение.
Размножение и развитие аппендикулярии. У аппендикулярии живорождение неизвестно. Отложенное яйцо (около 0,1 мм в диаметре) начинает дробиться целиком, и вначале дробление идет равномерно. Все стадии своего эмбрионального развития - бластулу, гаструлу и др.- аппендикулярии проходят очень быстро, и в результате развивается массивный зародыш. У него уже имеется тельце с глоточной полостью и мозговым пузырьком и хвостовой придаток, в котором располагаются в ряд друг за другом 20 клеток хорды. К ним примыкают мышечные клетки. Затем из четырех клеток формируется и нервная трубка, залегающая вдоль всего хвоста над хордой.
На такой стадии личинка покидает яйцевую оболочку. Она еще развита очень мало, но в то же время у нее имеются зачатки всех органов. Пищеварительная полость зачаточна. Нет ни рта, ни анального отверстия, но мозговой пузырек со статоцистом - органом равновесия - уже развит. Хвост личинки располагается в продолжение передне-задней оси ее тела, и его правая и левая стороны обращены соответственно вправо и влево.
Далее следует превращение личинки во взрослую аппендикулярию. Формируется кишечная петля, которая растет к брюшной стенке тела, где она открывается наружу анусом. В это же время глотка растет вперед, достигает внешней поверхности и прорывается ротовым отверстием. Образуются бронхиальные трубки, которые открываются с двух сторон тела жаберными отверстиями наружу и также соединяют полость глотки с внешней средой. Развитие пищеварительной петли сопровождается оттеснением хвоста с самого конца тела на его брюшную сторону. В это же время хвост поворачивается вокруг своей оси на 90° влево, так что его спинной гребень оказывается с левой стороны, а правая и левая стороны хвоста обращены теперь вверх и вниз. Нервная трубка вытягивается в нервный тяж, формируются нервные узелки, и личинка превращается во взрослую аппендикулярию.
Все развитие и метаморфоз личинок аппендикулярий характеризуется большой скоростью всех процессов, происходящих во время этого развития. Личинка вылупляется из яйца еще до окончания своего формирования. Такая скорость развития не вызывается воздействием каждый раз каких-то внешних причин. Она определяется внутренней природой этих животных и является наследственной.
Как мы увидим дальше, взрослые аппендикулярий по своему строению очень похожи на личинок асцидий. Только некоторые детали строения отличают их друг от друга. Существует точка зрения, что аппендикулярий всю свою жизнь остаются на личиночной стадии развития, но личинка их приобрела способность полового размножения. Такое явление известно в науке под названием неотении. Широко известен пример амфибии амбистомы, личинки которой, называемые аксолотлями, способны к половому размножению. Живя в неволе, аксолотли никогда не превращаются в амбистом. Они имеют жабры и хвостовой плавник и живут в воде, прекрасно размножаясь и давая потомство, подобное себе. Но если их кормить препаратом щитовидной железы, аксолотли заканчивают превращение, теряют жабры и, выходя на сушу, превращаются во взрослых амбистом. Неотения отмечена и у других земноводных- тритонов, лягушек, жаб. Из беспозвоночных животных она встречается у некоторых червей, ракообразных, пауков и насекомых.
Половое размножение на личиночных стадиях иногда бывает выгодным для животных. Неотения может быть не у всех особей данного вида, а лишь у тех, которые живут в особых, возможно малоблагоприятных для них, условиях, например при пониженной температуре. В результате получается возможность размножения в необычной обстановке. При этом животное не затрачивает много энергии на завершение полного превращения личинки во взрослую особь и скорость созревания увеличивается.
Неотения, вероятно, сыграла большую роль в эволюции животных. Одна из наиболее серьезных теорий о происхождении всего ствола вторичноротых животных - Deuterostomia, к которым относятся и все хордовые животные, в том числе и позвоночные, выводит их из свободноплавающих кишечнополостных ктенофор или гребневиков. Некоторые ученые считают, что предки кишечнополостных были сидячими формами, а гребневики произошли от плавающих в воде личинок древнейших кишечнополостных, приобретших способность полового размножения в результате прогрессивной неотении.
Размножение и развитие асцидий. Развитие асцидий происходит более сложным путем. Когда из яйцевой оболочки выходит личинка, она вполне похожа на взрослую аппендикулярию (рис. 179, А). Она так жег как и аппендикулярий, напоминает по внешнему виду головастика, удлиненно-овальное тельце которого несколько сжато с боков. Хвостик вытянут и окружен тонким плавником. По оси хвоста проходит хорда. Нервная система личинки образована нервной трубкой, залегающей над хордой в хвосте и образующей на переднем конце тела мозговой пузырек со статоцистом. В отличие от аппендикулярий у асцидий есть еще пигментированный глазок, способный реагировать на свет. На передней части спинной стороны располагается рот, ведущий в глотку, стенки которой пронизаны несколькими рядами жаберных щелей. Но, в отличие от аппендикулярий, жаберные щели даже у личинок асцидии открываются не прямо наружу, а в особую околожаберную полость, зачатки которой в виде двух впячивающихся с поверхности тела мешочков хорошо видны с каждой стороны тела. Они называются нерибранхиальными впячиваниями. На переднем конце тела личинки видны три липких прикрепительных сосочка.
Вначале личинки свободно плавают в воде, двигаясь при помощи своего хвоста. Размеры их тела достигают одного или нескольких миллиметров. Специальные наблюдения показали, что личинки плавают в воде недолго - 6-8 часов. За это время они могут преодолевать расстояния до 1 км, хотя большинство их оседает на дно сравнительно недалеко от родителей. Однако и в этом случае наличие свободноплавающей личинки способствует расселению неподвижных асцидий на значительные расстояния и помогает им распространяться по всем морям и океанам.
Оседая на дно, личинка прикрепляется при помощи своих липких сосочков к различным твердым предметам. Таким образом, личинка садится передним концом тела, и с этого момента она начинает вести неподвижный, прикрепленный образ жизни. В связи с этим происходит коренная перестройка и значительное упрощение строения тела (рис. 179, Б-Ж). Хвост вместе с хордой постепенно исчезает. Тело принимает мешковидную форму. Статоцист и глаз исчезают, а вместо мозгового пузырька остается только нервный ганглий и околонервная железка. Оба перибранхиальных впячивания начинают сильно разрастаться по бокам глотки и окружают ее. Два отверстия этих полостей постепенно сближаются и наконец сливаются на спинной стороне в одно клоакальное отверстие. В эту полость и открываются образующиеся вновь жаберные щели. Кишка также открывается в клоаку.
Садясь на дно своей передней частью, на которой находится рот, личинка асцидии попадает в очень невыгодное положение в смысле захвата пищи. Поэтому у осевшей личинки наступает еще одно важное изменение в общем плане строения тела: рот ее начинает медленно перемещаться снизу вверх и в конце концов располагается на самом верхнем конце тела (рис. 179, Г-Ж). Перемещение происходит по спинной стороне животного и влечет за собой смещение всех внутренних органов. Перемещающаяся глотка толкает перед собой мозговой ганглий, который в конце концов ложится на спинной стороне тела между ртом и клоакой. На этом заканчивается превращение, в результате которого животное оказывается совершенно непохожим по внешнему виду на собственную личинку.
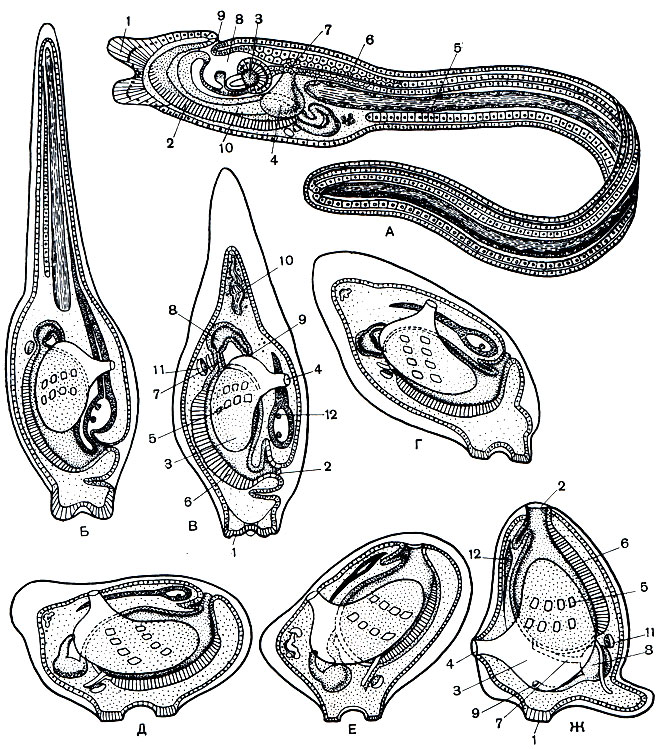
Рис. 179. Строение и превращение личинки асцидий. А - строение личинки асцидий: 1 - прикрепительные сосочки; 2 - эндостиль; 3 - глаза; 4 - перибранхиальная полость; 5 - хорда; 6 - нервная трубка; 7 - отверстие перибранхиальной полости; 8 - мозговой пузырь; 9 - рот; 10 - глотка. В-Ж - превращение личинки асцидии во взрослую особь: 1 - прикрепительные сосочки; 2 - рот; 3 - перибранхиальная полость; 4 - отверстие перибранхиальной полости; 5 - жаберные щели в глотке; 6 - эндостиль; 7 - пищевод; 8 - желудок; 9 - задний проход; 10 - рудимент хвоста; 11 - сердце; 12 - нервная трубка с остатками мозгового пузыря
Образовавшаяся таким образом асцидия может размножаться и иным, бесполым путем, посредством почкования. В наиболее простом случае от брюшной стороны тела у его основания вырастает колбасовидное выпячивание, или почкородный столон (рис. 180). Этот столон окружен наружным покровом тела асцидий (эктодермой), в него продолжается полость тела животного и, кроме того, слепое выпячивание задней части глотки. Длинный отросток в столон дает и сердце. Таким образом, в почкородный столон заходят зачатки важнейших систем органов. На поверхности столона образуются небольшие бугорки, или почки, в которые все перечисленные выше зачатки органов также дают свои отростки. Путем сложной перестройки эти зачатки образуют новые органы почки. Из выроста глотки развивается новый кишечник, из сердечного выроста - новый сердечный мешок. В покровах тела почки прорывается ротовое отверстие. Путем впячивания эктодермы снаружи внутрь образуется клоака и перибранхиальные полости. У одиночных форм такая почка, вырастая, отрывается от столона и дает начало новой одиночной асцидии, а у колониальных - почка остается сидеть на столоне, вырастает, вновь начинает почковаться, и в конце концов образуется новая колония асцидий. Интересно, что почки у колониальных форм с общей студенистой туникой всегда отделяются внутри нее, но не остаются на месте, где они образовались, а передвигаются сквозь толщу туники на свое окончательное место. Их почка всегда прокладывает себе дорогу к поверхности туники, куда и открываются ее ротовое и анальное отверстия. У одних видов эти отверстия открываются независимо от отверстий других почек, у других - наружу открывается только один рот, тогда как клоакальное отверстие открывается в общую для нескольких зооидов клоаку (рис. 174, Б). Иногда при этом могут образовываться длинные каналы. У многих видов зооиды образуют тесный кружок вокруг общей клоаки, а те, которые не поместились в нем, выталкиваются прочь и дают начало новому кружку зооидов и новой клоаке. Такое скопление зооидов образует так называемый кормидий.
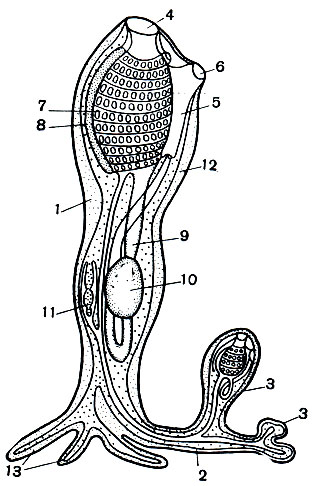
Рис. 180. Размножение сложных асцидий: 1 - асцидиозооид; 2 - столон; 3 - почки; 4 - ротовой сифон; 5 - клоака; 6 - клоакальный сифон; 7 - глотка с жаберными щелями; 8 - эндостиль; 9 - кишка; 10 - желудок; 11 - сердце; 12 - туника; 13 - корневые отростки столона с зачатками тканей
Иногда такие кормидий устроены очень сложно и имеют даже общую колониальную сосудистую систему. Кормидий окружается кольцевым кровеносным сосудом, в который впадает от каждого зооида по два сосудика. Кроме того, такие сосудистые системы отдельных кормидиев сообщаются и между собой, и возникает сложная общеколониальная кровеносная сосудистая система, так что все асцидиозооиды оказываются связанными между собой. Как мы видим, связь между отдельными членами колоний у разных сложных асцидий может быть или очень простой, когда отдельные особи вполне самостоятельны и только погружены в общую тунику, а почки обладают, кроме того, способностью передвигаться в ней, или же сложной, с единой кровеносной системой.
Кроме почкования посредством столона, возможны также и другие виды почкования - так называемые мантийное, пилорическое, постабдоминальное,- в зависимости от тех частей тела, которые дали начало почке. При мантийном почковании почка возникает в виде бокового выступа стенки тела в области глотки. Она состоит всего лишь из двух слоев: наружного - эктодермы и внутреннего - выроста око ложаберной полости, из которого формируются в дальнейшем все органы нового организма. Как и на столоне, почка постепенно округляется и отделяется от материнской особи тонкой перетяжкой, которая затем превращается в стебелек. Такое почкование начинается еще на стадии личинки и особенно ускоряется после того, как личинка садится на дно. Личинка, дающая начало почке (она в данном случае называется оозооид), погибает, а развивающаяся почка (или бластозоид) дает начало новой колонии. У других асцидий почка образуется на брюшной поверхности кишечного отдела тела, также очень рано, когда личинка еще не вылупилась. В этом случае в состав почки, покрытой эпидермисом, входят ответвления нижнего конца эпикарда, т. е. наружной стенки сердца. Первичная почка удлиняется, подразделяется на 4-5 частей, каждая из которых превращается в самостоятельный организм, а личинка - оозооид,- давшая начало этим почкам, распадается и служит для них питательной массой. Иногда в почку могут входить части пищеварительной системы желудка и задней кишки. Такой способ почкования называется пилорическим. Интересно, что в этом очень сложном случае почкования целый организм возникает в результате слияния двух почек в одну. Например, у Trididemnum в состав первой почки входят выросты пищевода, а во вторую - выросты эпикарда. После того как обе почки сольются, из первой образуется пищевод, желудок и кишечник дочернего организма, а также сердце, а из второй - глотка, пронизанная жабрами, и нервная система. После этого дочерний организм, обладающий уже полным набором органов, отшнуровывается от материнского. Однако и другие части тела могут дать начало почке. В некоторых случаях даже выросты хорды личинки могут входить в почку и из них формируется нервная система и половые железы дочерней особи. Иногда же процессы почкования бывают настолько похожи на простое деление организма на части, что трудно сказать, какой способ размножения имеется в данном случае. Кишечный отдел тела при этом сильно удлиняется, в нем накапливаются питательные вещества, которые получаются в результате распада грудного отдела. Затем происходит разделение брюшного отдела на несколько фрагментов, обычно называемых почками, из которых возникают новые особи. У Amaroucium вскоре после прикрепления личинки на заднем конце ее тела образуется длинный вырост. Он увеличивается в размере, и у асцидии в результате этого сильно развивается задняя часть тела - постабдомен, в который смещается сердце. Когда длина постабдомена намного начинает превышать длину тела личинки, происходит его отрыв от материнской особи и разделение на 3-4 части, из которых образуются молодые почки - бластозоиды. Они перемещаются из постабдомена вперед и располагаются рядом с материнским организмом, сердце в котором образуется заново. Развитие бластозооидов происходит неравномерно, и, когда одни из них уже закончили его, другие только начинают развиваться.
Процессы почкования у асцидий чрезвычайно разнообразны. Иногда даже близкие виды одного рода обладают разными способами почкования. Некоторые асцидии способны образовывать покоящиеся, остановившиеся в своем развитии почки, которые позволяют им переживать неблагоприятные условия.
При почковании у асцидий наблюдается следующее интересное явление. Как известно, в процессе эмбрионального развития различные органы организма животного возникают из разных, но совершенно определенных частей эмбриона (зародышевых листков) или слоев тела зародыша, составляющих его стенку на самых первых стадиях развития.
У большинства организмов имеется три зародышевых листка: наружный, или эктодерма, внутренний, или энтодерма, и средний, или мезодерма. У зародыша эктодерма покрывает тело, а энтодерма выстилает внутреннюю кишечную полость и обеспечивает его питание. Мезодерма осуществляет связь между ними. В процессе развития из эктодермы, как общее правило, образуется нервная система, кожные покровы, а у асцидий и околожаберные мешки, из энтодермы - пищеварительная система и органы дыхания, из мезодермы - мышцы, скелет и половые органы. При различных же способах почкования у асцидий это правило нарушается. Например, при мантийном почковании всем внутренним органам (в том числе и желудку и кишечнику, возникающим из энтодермы эмбриона) дает начало вырост околожаберной полости, являющийся по своему происхождению образованием эктодермическим. И наоборот, в случае, когда в состав почки входит вырост эпикарда (а сердце у асцидии в процессе эмбрионального развития образуется как вырост энтодермической глотки), большая часть внутренних органов, в том числе и нервная система и околожаберные мешки, образуется как производное энтодермы.
Размножение и развитие пиросом. Пиросомы также обладают бесполым способом размножения путем почкования. Но у них почкование происходит с участием специального постоянного выроста тела - почкородного столона. Оно характеризуется также и тем, что происходит на очень ранних стадиях развития. Яйца пиросом очень крупны, до 0,7 мм и даже до 2,5 мм, и богаты желтком. В процессе их развития образуется первый индивид - так называемый циатозооид. Циатозооид соответствует оозооиду асцидий, т. е. он является бесполой материнской особью, развившейся из яйца. Он очень рано перестает развиваться и разрушается. Всю основную часть яйцеклетки занимает питательный желток, на котором и развивается циатозооид.
У описанного недавно вида Pyrosoma vitjazi на желточной массе располагается циатозооид, который представляет собой вполне развитую асцидию размером в среднем около 1 мм (рис. 181, А). Есть даже маленькое ротовое отверстие, которое открывается наружу под яйцевой оболочкой. В глотке расположено 10-13 пар жаберных щелей и 4-5 пар кровеносных сосудов. Кишечник сформирован полностью и открывается в клоаку, сифон, который имеет форму широкой воронки. Имеется также нервный ганглий с околонервной железой и сердце, которое энергично пульсирует. Кстати, все это говорит о происхождении пиросом от асцидий. У других же видов в период максимального развития циатозооида можно различить лишь зачатки глотки с двумя жаберными щелями, зачатки двух околожаберных полостей, клоакальный сифон, нервный ганглий с околонервной железкой и сердце. Рот и пищеварительный отдел кишечника отсутствуют, хотя эндостиль и намечается. Развита также клоака с широким отверстием, открывающаяся в пространство под яйцевыми оболочками. На этой стадии еще в яйцевой оболочке у пиросом уже начинаются процессы бесполого развития. На заднем конце циатозооида образуется столон - эктодерма дает вырост, в который заходят продолжения эндостиля, околосердечной сумки и околожаберных полостей. Из эктодермы столона в будущей почке возникает нервный тяж, независимый от нервной системы самого циатозооида. В это время столон разделяется поперечными перетяжками на четыре участка, из которых и развиваются первые почки-бластозооиды, которые являются уже членами новой колонии, т. е. асцидиозооидами. Столон постепенно становится поперек оси тела циатозооида и желтка и закручивается вокруг них (рис. 181, Б-Е). Причем каждая почка становится перпендикулярно к оси тела циатозооида. По мере развития почек материнская особь - циатозооид - разрушается, а желточная масса постепенно используется в качестве питания для первых четырех почек-асцидиозооидов - родоначальников новой колонии. Четыре первичных асцидиозооида принимают геометрически правильное крестообразное положение и образуют общую клоакальную полость. Это настоящая маленькая колония (рис. 181, Е-Ж). В таком виде колония выходит из тела матери и освобождается от яйцевой оболочки. Первичные асцидиозооиды в свою очередь образуют столоны на своих задних концах, которые, перешнуровываясь, дают начало уже вторичным асцидиозооидам, и т. д. Как только асцидиозооид изолируется, на его конце образуется новый столон и каждый столон образует цепочку из четырех новых почек. Колония прогрессивно растет. Каждый асцидиозооид становится половозрелым и имеет мужские и женские половые железы.
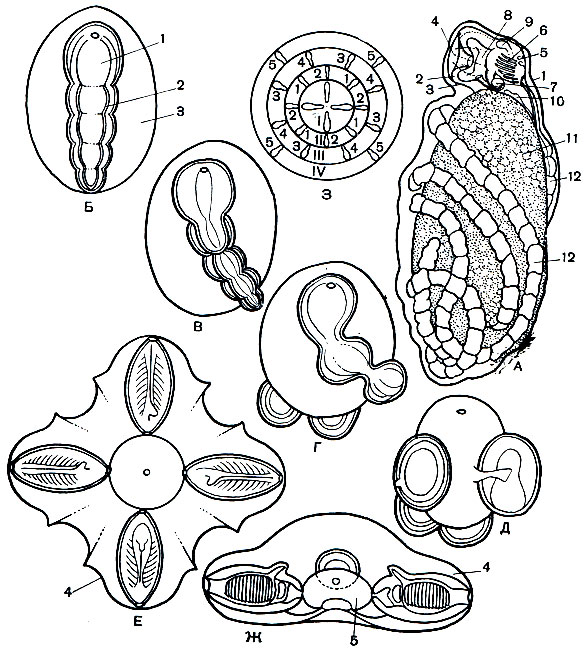
Рис. 181. Размножение и развитие пиросом. А - строение циатозооида: 1 - яйцевая оболочка; 2 - циатозооид; 3 - туника; 4 - клоакальный сифон; 5 - ротовой сифон; 6 - глотка с жаберными щелями и эндостилем; 7 - пищеварительный отдел кишечника; 8 - клоака; 9 - нервный ганглий; 10 - сердце; 11 - желток; 12 - первичный столон с почками. Б-Ж - схема образования первичной колонии: 1 - циатозооид; 2 - столон с четырьмя первичными почками; 3 - желток; 4 - первичная колония с четырьмя асцидиозооидами; 5 - полость колонии. 3 - последовательность развития асцидиозооидов в колонии пиросом (в центре - четыре первичных асцидиозооида): 1-5 - вторичные асцидиозооиды в порядке их образования. I-IV - кольцевые ряды асцидиозооидов
У одной группы пиросом асцидиозооиды сохраняют связь с материнской особью и остаются на том месте, где они возникли. В процессе образования почек столон удлиняется и почки соединяются между собой тяжами. Асцидиозооиды располагаются друг за другом по направлению к закрытому, переднему, концу колонии, в то время как первичные асцидиозооиды отодвигаются к ее задней, открытой, части.
У другой же группы пиросом, куда входит большинство их видов, почки не остаются на месте. Как только они достигают определенной стадии развития, они отделяются от столона, который никогда не удлиняется. В это же время они подхватываются особыми клетками - фороцитами. Фороциты - это крупные, похожие на амеб клетки. Они обладают способностью передвигаться в толще туники при помощи своих ложноножек, или псевдоподий, так же, как это делают амебы. Подхватывая почку, фороциты переносят ее через тунику, покрывающую колонию, на строго определенное место под первичными асцидиозооидами, и, как только конечный асцидиозооид отрывается от столона, фороциты переносят его вдоль левой стороны к спинной части его производителя, где он окончательно устанавливается таким образом, что старые асцидиозооиды все дальше отодвигаются к вершине колонии, а молодые оказываются на ее заднем конце.
Каждое новое поколение асцидиозооидов переносится с геометрической правильностью в строго определенное место по отношению к предыдущему поколению и располагается этажами (рис. 181, 3). После образования трех первых этажей между ними начинают возникать вторичные, затем третичные и т. д. этажи. Первичные этажи имеют по 8 асцидиозооидов, вторичные - по 16, третичные - по 32 и т. д. в геометрической прогрессии. Диаметр колонии увеличивается. Однако с ростом колонии четкость этих процессов нарушается, некоторые асцидиозооиды путаются и попадают в чужие этажи. У тех же самых индивидуумов в колонии пиросом, которые размножались почкованием, в дальнейшем развиваются половые железы, и они приступают к половому размножению. Как мы уже знаем, у каждого из многих асцидиозооидов пиросом, образовавшихся при помощи почкования, развивается только по одному крупному яйцу.
По способу образования колоний, а именно по тому, сохраняется ли у асцидиозооидов связь с материнским организмом в течение долгого времени или нет, пиросомы делятся на две группы - Pyrosoma fixata и Pyrosoma ambulata. Первые считаются более примитивными, так как перенос почек при помощи фороцитов является более сложным и более поздним приобретением пиросом.
Образование первичной колонии из четырех членов считалось настолько постоянным для пиросом, что этот признак вошел даже в характеристику всего отряда Pyrosomida. Однако в последнее время были получены новые данные по развитию пиросом. Оказалось, например, что у Pyrosoma vitjazi почкородный столон может достигать очень большой длины, а количество почек, одновременно образующихся на нем,- около 100. Такой столон образует под яйцевой оболочкой неправильные петли (рис. 181, А). К сожалению, пока еще остается неизвестным, каким образом происходит у них формирование колонии.
Размножение и развитие бочоночников и сальп. У бочоночников процессы размножения еще более сложны и интересны. Из яйца у них развивается хвостатая, как у асцидий, личинка, имеющая в хвостовом отделе хорду (рис. 182, А). Однако хвост в скором времени исчезает, а туловище личинки сильно разрастается и превращается во взрослого бочоночника, который по своему строению заметно отличается от половой особи, которую мы описали выше. Вместо восьми мускульных обручей у него имеется девять, есть маленький мешковидный орган равновесия - статоцист, жаберных щелей вдвое меньше, чем у половой особи. У него совершенно отсутствуют половые железы и, наконец, посередине брюшной стороны тела и на спинной стороне его заднего конца развиваются два особых выроста - столоны (рис. 182, Б). Эта бесполая особь носит особое название - кормилка. В нитевидный брюшной столон кормилки, который является почко-родным столоном, заходят выросты многих органов животного - продолжение полости тела, глотки, сердца и др.,- всего восемь разных зачатков. Этот столон очень рано начинает отпочковывать крошечные первичные почки, или так называемые предпочки. В это время у его основания толпится множество уже знакомых нам крупных фороцитов. Фороциты по двое, по трое подхватывают почки и несут их сначала по правому боку кормилки, а потом по ее спинной стороне к спинному столону (рис. 182, В, Г). Если при этом почки собьются с дороги, они погибают. Пока почки передвигаются и переходят на спинной столон, они все время продолжают делиться. Оказывается, что почки, образовавшиеся на брюшном столоне, не могут развиваться и жить на нем.
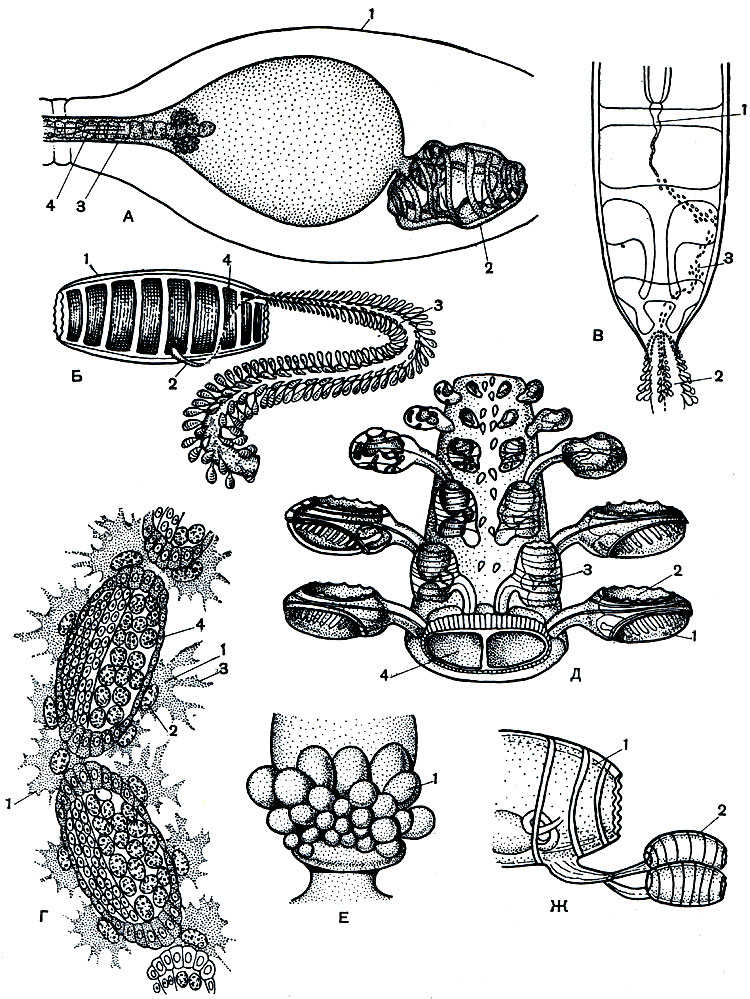
Рис. 182. Размножение и развитие бочоночников. А - личинка бочоночника: 1 - яйцевая оболочка; 2 - туловище личинки; з - хвост; 4 - хорда. Б - внешний вид почкующегося бесполого бочоночника (кормилки) со спинным столоном: 1 - кормилка; 2 - брюшной столон; 3 - спинной столон с почками; 4 - передвигающиеся почки. В - перенос почек фороцитами с брюшного столона на спинной: 1 - брюшной столон; 2 - спинной столон; 3 - путь почек по телу кормилки. Г - фороциты, несущие почки: 1 - фороциты; 2 - ядро клетки; 3 - ложноножки; 4 - почка с зачатками разных органов. Д - фрагмент спинного столона: 1 - боковые почки, или гастрозоиды; 2 - рот; 3 - срединные почки, или форозоиды; 4 - кровеносные лакуны. Е - стебелек форозоида с половыми предпочками, или гонозоидами: 1 - гонозоиды. Ж - свободноплавающий форозоид с гонозоидами: 1 - форозоид; 2 - гонозоиды
Первые порции почек усаживаются фороцитами на спинном столоне в два боковых ряда на его спинной стороне. Эти боковые почки очень быстро развиваются здесь в маленьких бочоночников, имеющих форму ложки, с огромным ртом, хорошо развитыми жабрами и кишечником (рис. 182, Д). Другие же органы у них атрофируются. Они прикрепляются к спинному столону кормилки собственным спинным столоном, имеющим форму отростка. Спинной столон кормилки в это время сильно растет - удлиняется и расширяется. В конце концов он может достигать 20-40 см длины. Он представляет собой длинный вырост тела, в который заходят две большие кровеносные лакуны кормилки.
Между тем подползают все новые и новые фороциты с почками, но теперь эти почки усаживаются уже не по бокам, а посередине столона, между двумя рядами описанных выше особей. Эти почки называются срединными или форозоидами. Они мельче боковых, и из них развиваются бочоночники, похожие на половозрелых особей, но не имеющие половых гонад. Эти бочоночники прирастают к столону кормилки особым тоненьким стебельком.
Все это время кормилка снабжает питательными веществами всю колонию. Они поступают сюда через кровеносные лакуны спинного столона и через стебельки почек. Но постепенно кормилка истощается. Она превращается в пустой мускулистый бочонок, который служит лишь для передвижения уже значительной колонии, образовавшейся на спинном столоне.
По поверхности этого бочонка продолжают двигаться все новые и новые почки, которые продолжает образовывать брюшной столон. С того момента, как кормилка превращается в пустой мешок, роль ее в питании колонии берут на себя большеротые боковые особи, которые называются гастрозоидами (питающими зооидами). Они захватывают и переваривают пищу. Усвоенные ими питательные вещества не только используются ими самими, но и передаются срединным почкам. А фороциты все еще подносят на спинной столон новые поколения почек. Теперь эти почки усаживаются уже не на самом столоне, а на тех стебельках, которыми прикрепляются срединные почки (рис. 182, Е). Вот эти-то почки и превращаются в настоящих половых бочоночников. После того как на стебельке срединной почки, или форозоида, укрепилась половая предпочка, он отрывается вместе со своим стебельком от общего столона и становится свободноплавающей маленькой самостоятельной колонией (рис. 182, Ж). Задача форозоида состоит в том, чтобы обеспечить развитие половой предпочки. Иногда его называют кормилкой второго порядка. Во время свободного периода жизни форозоидов половая предпочка, осевшая на его стебельке, делится на несколько половых почек-гонозоидов. Каждая такая почка вырастает в типичного полового бочоночника, который уже был описан в предыдущей части. Достигая зрелости, гонозоиды в свою очередь отделяются от своего форозоида и начинают вести жизнь самостоятельных одиночных бочоночников, способных к половому размножению. Надо сказать, что и у гастрозоидов и у форозоидов гонады также закладываются в процессе их развития, но затем они исчезают. Эти особи только помогают развитию третьего настоящего полового поколения.
После того как от спинного столона кормилки оторвутся все срединные почки, кормилка вместе с боковыми почками отмирает. Количество индивидуумов, образующихся на одной кормилке, чрезвычайно велико. Оно равняется нескольким десяткам тысяч.
Как мы видим, цикл развития бочоночников чрезвычайно сложен и характеризуется сменой полового и бесполого поколений. Краткая схема его такова: 1. Половая особь развивается на брюшном стебельке форозоида. 2. Половая особь откладывает яйца, и в результате их развития получается бесполая хвостатая личинка. 3. Из личинки непосредственно развивается бесполая кормилка. 4. На спинном столоне кормилки развивается поколение бесполых боковых гастрозоидов. 5. Новое поколение бесполых срединных форозоидов. 6. Появление и развитие на брюшном столоне оторвавшегося от кормилки форозоида половых гонозоидов. 7. Образование половой особи из гонозоида. 8. Откладывание яиц.
В развитии сальп также происходит смена поколений, однако она не имеет столь удивительной сложности, как у бочоночников. Личинки сальп не обладают хвостом, содержащим хорду. Развиваясь из единственного яйца в теле матери, в ее клоачной полости, зародыш сальп вступает в тесную связь со стенками материнского яичника, через которые к нему поступают питательные вещества. Это место соединения тела зародыша с тканями матери называется детским местом или плацентой. Свободноживущая личиночная стадия у сальп отсутствует, и у их зародыша имеется только рудимент (остаток, не получивший полного развития) хвоста и хорды. Это так называемый элеобласт, состоящий из крупных богатых жиром клеток (рис. 183, А). Только что развившийся зародыш, по существу еще эмбрион, вышедший через клоакальный сифон в воду, имеет на брюшной стороне вблизи сердца и между остатком плаценты и элеобластом маленький почкородный столон. У взрослых форм столон достигает значительной длины и обычно закручен спирально. Эта одиночная сальпа также представляет из себя такую же кормилку, как и образовавшийся из личинки бочоночник (рис. 183, Б). На столоне из боковых утолщений образуются многочисленные почки, расположенные двумя параллельными рядами. Обычно сначала захватывается почкованием какой-то определенный участок столона, дающий начало определенному числу одновозрастных почек. Количество их бывает различно - у разных видов от нескольких единиц до нескольких сотен. Затем начинает почковаться второй участок, третий и т. д. Все почки - бластозоиды - каждого отдельного участка или звена развиваются одновременно и равны по величине. В то время как в первом участке они достигают уже значительного развития, бластозоиды второго участка развиты гораздо менее и т. д., а на последнем участке столона почки только намечаются (рис. 183, В).
В процессе своего развития бластозоиды испытывают перегруппировку, оставаясь при этом связанными друг с другом столоном. Каждая пара зооидов приобретает определенное положение но отношению к другой паре. Она поворачивается своими свободными концами в противоположные стороны. Кроме того, в каждой особи происходит, как у асцидий, смещение органов, приводящее к изменению их первоначального взаиморасположения. На образование почек идет все вещество столона. У сальп все развитие почек протекает на брюшном столоне, и они не нуждаются в специальном спинном столоне. Почки отрываются от него не поодиночке, а целыми цепочками, соответственно тому как они возникли, и образуют временные колонии (рис. 183, Г). Все особи в них совершенно равноценны, и каждая развивается в половозрелое животное.
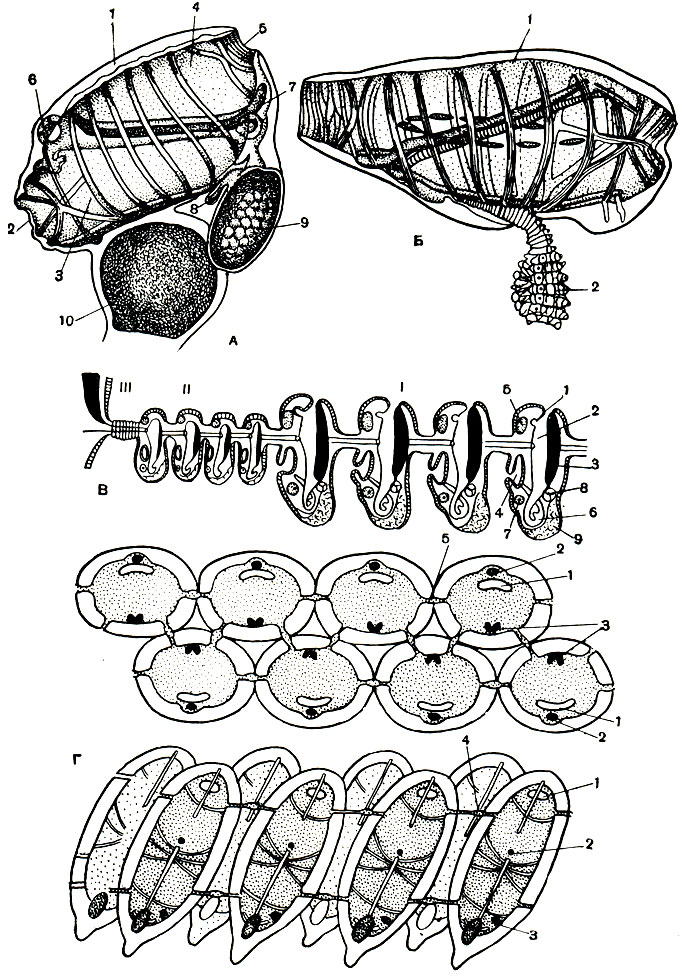
Рис. 183. Размножение и развитие сальп. А - личинка сальп: 1 - тельце личинки; 2 - ротовой сифон; 3 - глотка; 4 - клоака; 5 - клоакальный сифон; 6 - ганглий; 7 - кишка; 8 - столон; 9 - рудимент хорды, или элеобласт; 10 - плацента. Б - внешний вид почкующейся бесполой сальпы: 1 - сальпа; 2 - брюшной столон с почками. В - схема почкования столона: I-III - участки столона, на которых одновременно развиваются почки; 1 - рот; 2 - глотка; 3 - эндостиль; 4 - клоака; 5 - нервный ганглий; 6 - кишечник; 7 - половые железы; 8 - сердце; 9 - элеобласт. Г - схема расположения бластозооидов сальп в колонии. Вид сверху и сбоку: 1 - ротовой сифон; 2 - нервный ганглий; 3 - зародыш; 4 - прикрепительные папиллы
Интересно, что, в то время как нервная трубка, половой тяж, околожаберные полости и пр. у разных индивидуумов уже разделились, глотка остается общей в пределах одной цепочки. Таким образом, члены цепочки сначала органически соединены между собой столоном. Но отрывающиеся зрелые отрезки цепочки состоят из особей, соединенных между собой только слипшимися прикрепительными сосочками. Каждая особь имеет по восемь таких присосок, что обусловливает связь всей колонии. Эта связь и анатомически и физиологически крайне слаба. -
Колониальность таких цепочек в сущности едва выражена. Линейно вытянутые цепочки-колонии сальп - могут состоять из сотен отдельных индивидуумов. Однако у некоторых видов колонии могут иметь форму кольца. В этом случае особи соединены между собой выростами туники, направленными, как спицы в колесе, к центру кольца, по которому расположены члены колонии. Такие колонии состоят лишь из нескольких членов: у Cyclosalpa pinnata, например, из восьми-девяти особей (табл. 29).

Таблица 29. Оболочники. Одиночные асцидии: 1 - Phallusia mammillata; 2 - Halocynthia roretzi; 4 - Styela clavata; 5 - Ascidia virginea; 6 - Boltenia ovifera; 8 - Distomus variolosus; 9 - Boltenia echinata; 11 - Clavelina lepadiformis; 14 - Halocynthia aurantium; 16 - Giona intestinalis; 17 - Chelyosoma macleayanum; 18 - Pyura setosa; 19 - Pyura discoveryi; 20 - Styela plicata. Колониальные асцидии: 3 - Gynandrocarpa placenta; 7 - Sarcobotrylloides racemosum; 10 - Ecteinascidia tortugensis; 12 - Sycozoa umbellata; 13, 15 - Botryllus schlossery

Таблица 29. Оболочники. Одиночные асцидии: 1 - Phallusia mammillata; 2 - Halocynthia roretzi; 4 - Styela clavata; 5 - Ascidia virginea; 6 - Boltenia ovifera; 8 - Distomus variolosus; 9 - Boltenia echinata; 11 - Clavelina lepadiformis; 14 - Halocynthia aurantium; 16 - Giona intestinalis; 17 - Chelyosoma macleayanum; 18 - Pyura setosa; 19 - Pyura discoveryi; 20 - Styela plicata. Колониальные асцидии: 3 - Gynandrocarpa placenta; 7 - Sarcobotrylloides racemosum; 10 - Ecteinascidia tortugensis; 12 - Sycozoa umbellata; 13, 15 - Botryllus schlossery
Если теперь сравнить способы бесполого размножения разных туникат, то, несмотря на большую сложность и разнородность этого процесса в разных группах, нельзя не заметить и общих черт. А именно: у всех них наиболее распространенным способом размножения является деление почкородного столона на большее или меньшее число участков, дающих начало отдельным особям. Такие столоны есть и у асцидий, и у пиросом, и у сальп.
Колонии всех туникат возникают как следствие бесполого способа размножения. Но если у асцидий они появляются просто в результате почкования и каждый зооид в колонии может развиваться как бесполым, так и половым путем, то у пиросом и особенно у сальп их появление связано со сложным чередованием полового и бесполого поколений.
Образ жизни туникат. Посмотрим теперь, как живут разные оболочники и какое практическое значение они имеют. Мы уже говорили выше, что часть их живет на дне моря, а часть в толще воды. Асцидий являются донными животными. Всю свою жизнь взрослые формы проводят неподвижно, прикрепившись к какому-нибудь твердому предмету на дне и прогоняя через свою пронизанную жабрами глотку воду, чтобы отфильтровать из нее мельчайшие клетки фитопланктона или мелких животных и частицы органического вещества, которыми асцидий питаются. Они не могут передвигаться, и, только испугавшись чего-либо или проглотив что-нибудь слишком крупное, асцидия может сжаться в комочек. При этом вода с силой выбрасывается из сифона.
Как правило, асцидий просто прирастают к камням или другим твердым предметам нижней частью своей туники. Но иногда тело их может приподниматься над поверхностью грунта на тонком стебельке. Такое приспособление позволяет животным "облавливать" больший объем воды и не тонуть в мягком грунте. Оно особенно характерно для глубоководных асцидий, которые живут на тонких илах, покрывающих дно океана на большой глубине. Для того чтобы не тонуть в грунте, у них может быть еще и другое приспособление. Отростки туники, которыми в обычном случае асцидий прикрепляются к камням, разрастаются и образуют своего рода "парашют", удерживающий животное на поверхности дна. Такие "парашюты" могут появляться и у типичных обитателей твердых грунтов, обычно селящихся на камнях, при переходе их к жизни на мягких илистых грунтах. Корневидные выросты тела позволяют особям одного и того же вида выйти в новую и не обычную для них среду обитания и расширить границы своего ареала, если другие условия благоприятны для их развития.
В последнее время асцидий были обнаружены среди очень специфической фауны, населяющей тончайшие ходы между песчинками грунта. Такая фауна называется интерстициальной. Теперь уже известно семь видов асцидий, избравших местом своего обитания столь необычный биотоп. Это чрезвычайно мелкие животные - размер их тела всего лишь 0,8-2 мм в диаметре. Некоторые из них подвижны.
Одиночные асцидий образуют иногда большие сростки, которые разрастаются в целые друзы и селятся большими скоплениями. Как уже говорилось, очень многие виды асцидий колониальны. Чаще других встречаются массивные студенистые колонии, отдельные члены которых погружены в общую довольно толстую тунику. Такие колонии образуют корковые наросты на камнях или же встречаются в виде своеобразных шаров, лепешек и выростов на ножках, иногда напоминающих по форме грибы. В других случаях отдельные особи колоний могут быть почти самостоятельными.
Некоторые асцидии, например Claveiina, обладают способностью легко восстанавливать, или регенерировать, свое тело из разных его частей. Каждый из трех отделов тела колониальной клавелины - грудной отдел с жаберной корзиной, отдел тела, содержащий внутренности, и столон,- будучи вырезанным, способен воссоздать целую асцидию. Удивительно, что даже из столона вырастает цельный организм с сифонами, всеми внутренностями и нервным ганглием. Если выделить у клавелины кусок жаберной корзины одновременно проводимыми двумя поперечными разрезами, то на переднем конце превратившегося в округлый комочек фрагмента животного образуется новая глотка с жаберными щелями и сифоны, а на заднем - столон. Если же сделать сначала разрез сзади, а потом спереди, то удивительным образом глотка с сифонами образуется на заднем конце, а сифон на переднем и переднезадняя ось тела животного поворачивается на 180°. Некоторые асцидии способны в каких-то определенных случаях сами отбрасывать части своего тела, т. е. они способны к аутотомии. И подобно тому как заново вырастает у ящерицы ее оторвавшийся хвост, из оставшегося куска тела вырастает новая асцидия. Способность асцидии к восстановлению утраченных частей тела особенно ярко выражена во взрослом состоянии у тех видов, которые могут размножаться путем почкования. Виды же, размножающиеся лишь половым путем, например одиночная Ciona intestinalis, обладают регенеративной способностью в гораздо меньшей степени.
Процессы регенерации и бесполого размножения имеют много сходных черт, и, например, Ч. Дарвин утверждал, что эти процессы имеют единую основу. Способность восстанавливать утраченные части тела особенно сильно развита у простейших, кишечнополостных, червей и оболочников, т. е. у тех групп животных, которым особенно свойственно бесполое размножение. И в каком-то смысле само бесполое размножение можно рассматривать как проявляющуюся в естественных условиях существования и локализованную в определенных частях тела животного способность к регенерации его из фрагмента тела.
Асцидии широко распространены как в холодных морях, так и в теплых. Они есть и в Северном Ледовитом океане, и в Антарктике. Они были найдены даже непосредственно на побережье Антарктиды при обследовании советскими учеными одного из фиордов "оазиса" Бангера. Фиорд был отгорожен от моря нагромождениями многолетнего льда, и поверхностная вода в нем была сильно опреснена. На каменистом и безжизненном дне этого фиорда удалось обнаружить лишь комочки диатомовых и нити зеленых водорослей. Однако в самом куту залива были найдены остатки морской звезды и большое количество крупных, до 14 см длиной, розовато-прозрачных студенистых асцидии. Животные были оторваны от дна, вероятно, штормом и прибиты сюда течением, но желудки и кишечники их были целиком заполнены зеленой массой несколько переваренного фитопланктона. Вероятно, они питались незадолго до того, как были выловлены из воды у самого берега.
Особенно разнообразны асцидии в тропической зоне. Имеются данные о том, что число видов оболочников в тропиках примерно в 10 раз больше, чем в умеренных и полярных областях. Интересно, что в холодных морях асцидии значительно крупнее, чем в теплых, и поселения их более многочисленны. Они, так же как и другие морские животные, подчиняются общему правилу, по которому в умеренных и холодных морях обитает меньшее количество видов, но они образуют значительно большие поселения и биомасса их на 1 м2 поверхности дна во много раз больше, чем в тропиках.
Большинство асцидии живет в самой поверхностной литоральной или приливо-отливной зоне океана и в верхних горизонтах материковой отмели или сублиторали до глубины 200 м. С увеличением глубины общее количество видов их уменьшается. В настоящее время глубже 2000 м известно 56 видов асцидии. Максимальная глубина их обитания, на которой были найдены эти животные,- 7230 м. На этой глубине асцидии были обнару жены во время работ советской океанологической экспедиции на корабле "Витязь" в Тихом океане. Это были представители характерного глубоководного рода Culeolus. Округлое тело этой асцидий с очень широко открытыми сифонами, совершенно не выступающими над поверхностью туники, сидит на конце длинного и тонкого стебелька, которым кулеолюс может прикрепляться к мелким камушкам, спикулам стеклянных губок и другим предметам на дне. Стебелек не может выдержать веса довольно крупного тела, и, вероятно, оно плавает, колеблясь над дном, уносимое слабым течением. Окраска его беловато-серая, бесцветная, как у большинства глубоководных животных (рис. 184).

Рис. 184. Глубоководные оболочники: 1 - Guleolus murrayi; 2 - Ascopera gigantea; 3 - Octacnemus bythius
Асцидий избегают опресненных участков морей и океанов. Громадное большинство их живет при нормальной океанической солености около 350/00.
Как уже говорилось, наибольшее количество видов асцидий обитает в океане на небольшой глубине. Здесь же они образуют и наиболее массовые поселения, особенно там, где в толще воды достаточно взвешенных частиц - планктона и детрита,- служащих им пищей. Асцидий селятся не только на камнях и других жестких естественных предметах. Излюбленным местом их поселения являются также днища кораблей, поверхность разных подводных сооружений и т. д. Поселяясь иногда в огромных количествах вместе с другими организмами-обрастателями, асцидий могут причинять большой вред хозяйству. Известно, например, что, поселяясь на внутренних стенках водоводов, они развиваются в таком количестве, что сильно сужают диаметр труб и засоряют их. При массовом отмирании в определенные сезоны года они настолько забивают фильтрационные устройства, что подача воды может прекращаться совсем и промышленные предприятия терпят значительный ущерб.
Одна из наиболее широко распространенных асцидий - Ciona intestinalis,- обрастая днища кораблей, может поселяться в таких огромных количествах, что скорость корабля значительно снижается. Потери транспортного судоходства в результате обрастаний очень велики и могут составлять миллионы рублей в год.
Однако способность асцидий образовывать массовые скопления благодаря одной удивительной их особенности может представлять известный интерес для людей. Дело в том, что в крови асцидий вместо железа содержится ванадий, который выполняет ту же роль, что и железо,-служит для переноса кислорода.
Ванадий- редкий элемент, имеющий большое практическое значение, растворен в морской воде в чрезвычайно малых количествах. Асцидии обладают способностью концентрировать его в своем теле. Количество ванадия составляет 0,04-0,7% веса золы животного. Следует вспомнить еще, что и туника асцидий содержит другое ценное вещество- целлюлозу. Количество ее, например, в одном экземпляре наиболее широко распространенного вида Ciona intestinalis равно 2-3 мг. Эти асцидии поселяются иногда в огромных количествах. Число особей на 1 м2 поверхности достигает 2500-10 000 экземпляров, а их сырой вес - 140 кг на 1 м2.
Появляется возможность обсуждения, как можно практически использовать асцидий в качестве источника этих веществ. Не везде есть древесина, из которой добывается целлюлоза, а залежи ванадия немногочисленны и рассеянны. Если же устроить подводные "морские огороды", то асцидий в огромных количествах можно выращивать на специальных пластинах. Подсчитано, что с 1 га площади моря можно получать от 5 до 30 кг ванадия и от 50 до 300 кг целлюлозы.
В толще воды океана обитают пелагические оболочники - аппендикулярии, пиросомы и сальпы. Это мир прозрачных фантастических существ, обитающих в основном в теплых морях и в тропической зоне океана. Большинство их видов настолько сильно приурочено в своем распространении к теплым водам, что они могут служить индикаторами изменений гидрологических условий в различных районах океана. Например, появление или исчезновение пелагических оболочников, в частности сальп, в Северном море в определенные периоды связано с большим или меньшим поступлением в эти районы теплых атлантических вод. То же самое явление неоднократно отмечалось в районе Исландии, Ла-Манша, около полуострова Ньюфаундленд и было связано как с ежемесячными, так и с сезонными изменениями в распространении теплых атлантических и холодных арктических вод. В эти районы заходят только три вида сальп - Salpa fusiformis, Jhlea asymmetrica и наиболее широко распространенная в океане Thalia democratica. Появление всех этих видов в большом количестве у побережий Британских островов, Исландии, Фарерских островов и в Северном море редко и связано с потеплением вод. У побережья Японии пелагические туникаты являются показателем пульсаций течения Куросио.
Пиросомы и сальпы особенно чувствительны к холодным водам и предпочитают не покидать тропическую зону океана, в которой они распространены очень широко. Области географического распространения большинства видов сальп, например, охватывают теплые воды всего Мирового океана, где их встречается более 20 видов. Правда, описаны два вида сальп, обитающих в Антарктике. Это Salpa thompsoni, распространенная во всех антарктических водах и не выходящая за пределы 40° ю. ш., т. е. зоны субтропического опускания холодных антарктических вод, и Salpa gerlachei, обитающая только в море Росса. Аппендикулярии распространены более широко, имеется около десяти видов, обитающих, например, в морях Северного Ледовитого океана, но и они более разнообразны и многочисленны в тропических районах.
Пелагические оболочники встречаются при нормальной океанической солености в 34-360/00. Известно, например, что в районе впадения реки Конго, где температурные условия являются весьма благоприятными для сальп, они отсутствуют вследствие того, что соленость в этом месте африканского побережья равна всего лишь 30,40/00. С другой стороны, сальпы отсутствуют и в восточной части Средиземного моря у Сирии, где соленость, наоборот, слишком высока - 400/00.
Все планктонные формы туникат - жители поверхностных слоев воды, главным образом от 0 до 200 м. Пиросомы, видимо, не идут глубже 1000 м. Сальпы и аппендикулярии в основной массе тоже не опускаются глубже нескольких сотен метров. Однако в литературе имеются указания о нахождении пиросом на глубине 3000 м, бочоночников - 3300 м и сальп даже до 5000 м. Но трудно сказать, обитают ли на такой большой глубине живые сальпы, или же это были просто их мертвые, но хорошо сохранившиеся оболочки.
На "Витязе" в ловах, сделанных закрывающейся сеткой, пиросомы не были обнаружены глубже 1000 м, а бочоночники - 2000-4000 м.
Все пелагические оболочники в общем широко распространены в океане. Часто они попадаются в сети зоолога единичными экземплярами, но столь же характерны для них и большие скопления. Аппендикулярии попадаются в значительном количестве - 600-800 экземпляров в лове с глубины до 100 м. У берегов Ньюфаундленда количество их значительно больше, иногда свыше 2500 экземпляров в таком лове. Это составляет приблизительно 50 экземпляров в 1 л3 воды. Но благодаря тому что аппендикулярии очень мелки, биомасса их при этом бывает незначительна. Обычно она составляет 20-30 мг на 1 м2 в холодноводных районах и до 50 мг на 1 м2 в тропических районах.
Что касается сальп, то они способны собираться иногда в огромнейших количествах. Известны случаи, когда скопления сальп останавливали даже большие корабли. Вот как описывает один такой случай участник советской антарктической экспедиции зоолог К. В. Беклемишев: "Зимой 1956-1957 гг. теплоход "Кооперация" ( водоизмещением более 5000 т) доставлял вторую смену зимовщиков в Антарктику, в поселок Мирный. Ясным ветреным утром 21 декабря 1956 г. в южной части Атлантического океана с палубы судна на поверхности воды было замечено 7-8 рыжеватых полос, вытянувшихся по ветру почти параллельно курсу корабля. Когда судно приблизилось, полосы уже не казались рыжими, но вода в них все же была не синей (как вокруг), а беловато-мутной от присутствия массы каких-то существ. Ширина каждой полосы была больше метра. Расстояние между ними - от нескольких метров до нескольких десятков метров. Длина полос - около 3 км. Едва "Кооперация" начала пересекать под острым углом эти полосы, как вдруг машина остановилась и судно легло в дрейф. Оказалось, что планктон забил машинные фильтры и прекратилась подача воды к двигателю. Во избежание аварии машину пришлось, застопорить, чтобы прочистить фильтры.
Взяв пробу воды, мы обнаружили в ней массу продолговатых прозрачных существ величиной около 1-2 см, называемых Thalia longicaudata и относящихся к отряду сальп. В 1 м3 воды их было не менее 2500 экземпляров. Ясно, что решетки фильтров были ими совершенно заполнены. Водоразборные кингстоны "Кооперации" расположены на глубине 5 м и 5,6 м. Следовательно, сальпы находились в большом количестве не только на поверхности, но и на глубине по крайней мере 6 м".
Массовое развитие оболочников и их доминирование в планктоне, по-видимому, характерное явление для краев тропической области. Скопления сальп отмечаются в северной части Тихого океана, массовое их развитие известно в зоне смешения вод течений Куросио и Оясио, у западного Алжира, к западу от Британских островов, у Исландии, в северо-западной Атлантике в прибрежных районах, вблизи южной границы тропической области в Тихом океане, у юго-восточной Австралии. Иногда сальпы могут преобладать в планктоне, в котором уже нет других типичных тропических представителей.
Что касается пиросом, то в столь огромных количествах, как описывалось выше для сальп, они, видимо, не встречаются. Однако в некоторых краевых районах тропической области обнаружены и их скопления. В Индийском океане на 40-45° ю. ш. во время работы советской антарктической экспедиции было встречено огромное количество крупных пиросом. Пиросомы располагались на самой поверхности воды пятнами. В каждом пятне насчитывалось от 10 до 40 колоний, которые ярко светились голубым светом. Между пятнами расстояние было 100 м и больше. В среднем на 1 м2 поверхности воды приходилось 1-2 колонии. Аналогичные скопления пиросом отмечались и у берегов Новой Зеландии.
Пиросомы известны как исключительно пелагические животные. Однако сравнительно недавно в проливе Кука у Новой Зеландии удалось получить несколько фотографий с глубины 160-170 м, на которых ясно были видны большие скопления Pyrosoma atlanticum, колонии которых просто лежали на поверхности дна.
Другие же особи плавали в непосредственной близости от дна. Было дневное время, и, возможно, животные ушли на большую глубину, чтобы спрятаться от прямых солнечных лучей, как это делают многие планктонные организмы.
Судя по всему, они чувствовали себя хорошо, так как условия среды были благоприятны для них. В мае эта пиросома обычна в поверхностных водах пролива Кука. Интересно, что в том же районе в октябре дно на глубине 100 м бывает покрыто мертвыми, разлагающимися пиросомами. Вероятно, это массовое отмирание пиросом связано с сезонными явлениями. Оно в какой-то степени дает представление о том, в каких количествах могут встречаться эти животные в море.
Пиросомы, что в переводе на русский язык означает "огнетелки", получили свое название от свойственной им способности к свечению. Было установлено, что свет, возникающий в клетках органов свечения пиросом, вызывается особыми симбиотическими бактериями. Они поселяются внутри клеток светящихся органов и, видимо, там и размножаются, поскольку неоднократно наблюдались бактерии со спорами внутри их. Светящиеся бактерии передаются из поколения в поколение. Током крови они переносятся к яйцам пиросом, находящимся на последней стадии развития, и заражают их. Затем они поселяются между бластомерами дробящегося яйца и проникают в зародыш. Светящиеся бактерии проникают вместе с током крови и в почки пиросом. Таким образом, молодые пиросомы получают светящихся бактерий в наследство от материнских особей. Однако не все ученые согласны с тем, что пиросомы светятся благодаря бактериям-симбионтам. Дело в том, что свечение бактерий характеризуется своей непрерывностью, а пиросомы испускают свет только после какого-нибудь раздражения. Свет асцидиозооидов в колонии может быть удивительно интенсивным и очень красивым. Кроме пиросом, светятся сальпы и аппендикулярии.
Ночью в тропическом океане вслед за идущим кораблем остается светящийся след. Волны, бьющиеся о борта суднат тоже вспыхивают холодным пламенем - серебристым, голубоватым или зеленовато-белым. В море светятся не только пиросомы. Известны многие сотни видов светящихся организмов - различные медузы, ракообразные, моллюски, рыбы. Нередко вода в океане горит ровным немерцающим пламенем от мириад светящихся бактерий. Светятся даже донные организмы. Мягкие кораллы горгонии в темноте горят и переливаются, то ослабляя, то усиливая свечение, разными огнями - фиолетовым, пурпурным, красным и оранжевым, голубым и всеми оттенками зеленого цвета. Иногда их свет похож на раскаленное добела железо. Среди всех этих животных огнетелки по яркости своего свечения, безусловно, занимают первое место. Иногда в общей светящейся массе воды как отдельные яркие шары вспыхивают более крупные организмы. Как правило, это бывают пиросомы, медузы или сальпы. Арабы называют их "морскими фонарями" и говорят, что их свет похож на свет месяца, слегка закрытого облаками. Овальные пятна света на небольшой глубине часто упоминаются при описании морского свечения. Например, в выписке из журнала теплохода "Алинбэк", приводимой Н. И.Тарасовым в его книге "Свечение моря", в июле 1938 г. в южной части Тихого океана были отмечены пятна света по преимуществу правильной прямоугольной формы, величина которых была приблизительно 45 х 10 см. Свет пятен был очень ярким, зеленовато-голубым. Особенно заметным стало это явление при наступившем шторме. Этот свет испускали пиросомы. Большой знаток в области свечения моря Н. И. Тарасов пишет, что колония пиросом может светиться до трех минут, после чего свечение прекращается сразу и полностью. Свет пиросом обычно голубой, но у утомленных, перевозбужденных и гибнущих животных он становится оранжевым и даже красным. Однако не все пиросомы могут светиться. Описанные выше гигантские пиросомы из Индийского океана так же, как и новый вид Pyrosoma vitjazi, не имеют органов свечения. Но не исключено, что способность к свечению у пиросом непостоянна и связана с определенными стадиями развития их колоний.
Как уже говорилось, сальпы и аппендикулярии тоже могут светиться. Свечение некоторых сальп заметно даже днем. Знаменитый русский мореплаватель и ученый Ф. Ф. Беллинсгаузен, проходя в июне 1821 г. мимо Азорских островов и наблюдая свечение моря, писал, что "море было усеяно светящимися морскими животными, они прозрачны, цилиндрообразны, длиною в два с половиною и в два дюйма, плавают соединенные одно с другим в параллельном положении, составляя, таким образом, род ленты, длина коей нередко аршин". В этом описании легко узнать сальп, которые встречаются в море как поодиночке, так и колониями. Чаще светятся только одиночные формы.
Если у сальп и пиросом имеются специальные органы свечения, то у аппендикулярий светится все туловище и некоторые места студенистого домика, в котором они живут. При разрыве домика внезапно возникает вспышка зеленого свечения по всему туловищу. Светятся, вероятно, желтые капельки особых секреторных выделений, имеющиеся на поверхности тела и внутри домика. Аппендикулярии, как уже говорилось, распространены шире, чем другие оболочники, и чаще встречаются в холодных водах. Нередко именно они обусловливают свечение воды в северной части Берингова моря, а также и в Черном море.
Свечение моря - необыкновенно красивое зрелище. Можно часами любоваться искрящимся буруном воды за кормой идущего корабля. Нам неоднократно приходилось работать в ночные часы во время экспедиции "Витязя" в Индийском океане. Большие планктонные сети, приходившие из глубины моря, часто бывали похожи на мерцающие голубоватым пламенем большие конусы, а куты их, в которых скапливается морской планктон, напоминали какие-то волшебные фонари, дающие такой яркий свет, что при нем вполне можно было читать. Вода стекала с сетей и с рук, падала на палубу огненными каплями.
Но свечение моря имеет и очень большое практическое значение, не всегда благоприятное для человека. Иногда оно сильно мешает кораблевождению, слепит и ухудшает видимость в море. Яркие вспышки его могут быть приняты даже за свет несуществующих маяков, не говоря о том, что светящийся след демаскирует в ночное время военные корабли и подводные лодки и наводит на цель флот и самолеты противника. Свечение моря часто мешает морскому промыслу, отпугивая рыб и морских зверей от облитых серебристым сиянием сетей. Но, правда, и большие скопления рыб могут быть легко обнаружены в темноте по вызванному ими свечению моря.
Оболочники иногда могут вступать в интересные взаимоотношения с другими пелагическими животными. Например, пустые оболочки сальп часто используются планктонными рачками гипериидами-фронимами в качестве надежного убежища для выведения потомства. Так же как и сальны, фронимы абсолютно прозрачны и незаметны в воде. Забравшись внутрь сальпы, самка фронимы выгрызает все внутри туники и остается в ней. В океане часто можно найти пустые оболочки сальп, в каждой из которых сидит по одному рачку. После того как в своеобразном родильном доме выведутся маленькие рачки, они прицепляются к внутренней поверхности туники и сидят на ней довольно продолжительное время. Мать же, усиленно работая своими плавательными ножками, прогоняет сквозь пустой бочонок воду, чтобы ее детям было достаточно кислорода. Самцы, видимо, никогда не поселяются внутри сальп. Все оболочники питаются взвешенными в воде мельчайшими одноклеточными водорослями, мелкими животными или же просто частицами органического вещества. Они являются активными фильтраторами. У аппендикулярии, например, выработалась специальная, очень сложно устроенная система фильтров и ловчих сетей для отлова планктона. Об устройстве их уже писалось выше. Некоторые сальпы обладают способностью скапливаться в огромные стаи.
При этом они могут настолько сильно выедать фитопланктон в тех районах моря, где они скапливаются, что серьезно конкурируют из-за пищи с другим зоопланктоном и являются причиной резкого уменьшения его количества. Известно, например, что у Британских островов могут образовываться большие скопления Salpa fusiformis, покрывающие площади до 20 тыс. квадратных миль. В районе своего скопления сальпы отфильтровывают фитопланктон в таком количестве, что почти нацело выедают его. При этом зоопланктон, в основном состоящий из мелких рачков Copepoda, также сильно уменьшается в количестве, поскольку Copepoda, как и сальпы, питаются плавающими микроскопическими водорослями.
Если такие скопления сальп в течение долгого времени держатся в одной и той же массе воды и такие воды сильно обедненные фито- и зоопланктоном, вторгаются в прибрежные области, они могут оказать серьезное воздействие на местное животное население. Выметанные личинки донных животных гибнут из-за отсутствия корма. Даже сельдь становится очень редкой в таких местах, возможно, из-за недостатка пищи или из-за большого количества растворенных в воде продуктов обмена оболочников. Однако столь большие скопления сальп - явление недолговечное, особенно в более холодноводных районах океана. При похолодании они исчезают.
Сами сальпы, так же как и пиросомы, могут иногда использоваться в пищу рыбами, но только очень немногими видами. К тому же туника их содержит очень незначительное количество усвояемых органических веществ. Известно, что в годы наиболее массового развития сальп в районе Оркнейских островов ими питалась треска. Летучие рыбы и желтоперые тунцы поедают сальп, а в желудках меч-рыбы были найдены пиросомы. Из кишечника другой рыбы - мунуса - размером в 53 см однажды было извлечено 28 пиросом. Аппендикулярии также встречаются иногда в желудках рыб, причем даже в значительных количествах. Сальпами и пиросомами могут, очевидно, питаться и те рыбы, которые поедают медуз и гребневиков. Интересно, что крупные пелагические черепахи каретты и некоторые антарктические птицы поедают одиночных сальп. Но большого значения в качестве пищевого объекта оболочники не имеют.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'