Происхождение млекопитающих
Предками млекопитающих, несомненно, были древние палеозойские рептилии, еще не утратившие некоторых черт строения амфибий: кожные железы, расположение сочленений в конечностях (между голенью и предплюсной и между предплечьем и запястьем) и, быть может, двойной затылочный мыщелок. Такой предковой группой считают подкласс звероподобных (Theromorpha).
Особенно близок к млекопитающим один из отрядов этого подкласса - отряд зверозубых (Theriodontia), у представителей которого зубы были дифференцированы на те же категории, что и у млекопитающих, т. е. на резцы, клыки и коренные, и сидели они в альвеолах. Имелось вторичное костное нёбо - признак весьма характерный для млекопитающих. Квадратная кость и сочленовная кость нижней челюсти были сильно уменьшены. При этом сочленовная кость не была приращена к нижней челюсти. Зубная кость, наоборот, была очень крупной. Здесь следует учесть, что у млекопитающих нижняя челюсть состоит только из зубной кости, а сочленовная кость превращена в одну из слуховых косточек - молоточек".
Такие известные высшие зверозубые, как иностранцевия (из пермских отложений на Северной Двине) и циногнатус (из триасовых отложений Южной Африки), не могли быть предками зверей, так как были высокоспециализированными и при этом крупными (2-3 м длиной) животными. Малые размеры древнейших млекопитающих дают основание искать предков зверей среди более примитивных и мелких зверозубых. В этом отношении представляет интерес иктидозавр (из верхнего триаса Южной Африки). Это было животное размером с крысу, скелет которого удивительно сходен с таковым млекопитающих; только нижняя челюсть у него состояла из нескольких костей. Очевидно, от таких мелких зверозубых и произошли млекопитающие.
Зверозубые просуществовали недолгое время, лишь до верхнего триаса.
Находки древнейших млекопитающих приурочены к юрским отложениям. Это были разнообразные и в ряде случаев высокоспециализированные животные; следовательно, возникновение млекопитающих надо отнести к более раннему времени - к триасу. С конца этого периода и до первой половины третичного времени существовали так называемые многобугорчатые. Это были мелкие, размером с крысу, зверьки, и только немногие достигали размеров сурка. Свое название они получили в связи с наличием на коренных зубах многочисленных бугорков. Клыков у них не было, а резцы развивались, наоборот, очень сильно.
Многобугорчатые представляли специализированных растительноядных зверей, и их нельзя считать прямыми предками последующих групп млекопитающих. Лишь предположительно допускают, что наиболее ранние формы могли дать начало однопроходным. Основанием для такого предположения служит сходство строения их зубов с зубами зародыша утконоса (рис. 12).
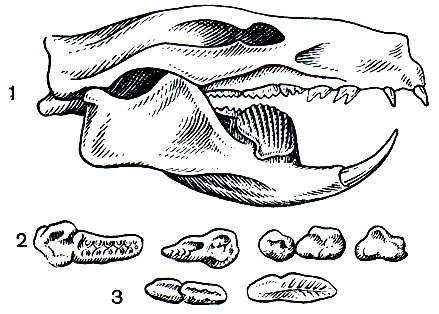
Рис. 12. Череп и зубы многобугорчатого Ptilodus (длина черепа около 8 см): 1 - череп; 2 - верхние зубы; 3 - нижние зубы
В отложениях с середины юры и до мела обнаруживаются многообразные трехбугорчатые, открывшие подлинную историю класса млекопитающих. Зубная система у них менее специализированная, чем у многобугорчатых; зубной ряд сплошной (рис. 13). Это были мелкие зверьки, близкие к насекомоядным; питались они частью животной, частью растительной пищей. Трехбугорчатые, в частности пантотерии,- наиболее вероятные предки сумчатых и плацентарных.
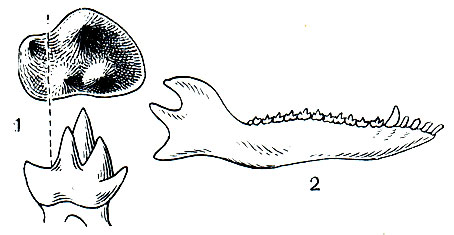
Рис. 13. Зубы (1) и нижняя челюсть (2) трехбугорчатого Amphitherium из средней юры
Сумчатые появились, видимо, в начале мелового периода. Однако первые ископаемые остатки их известны из верхнемеловых отложений Северной Америки. Это были мелкие зверьки типа опоссумов с бугорчато-режущими коренными и крупными клыками. В отложениях нижнетретичного времени они известны и в Евразии. Таким образом, родиной сумчатых следует считать северное полушарие. Однако еще до конца третичного времени они были вытеснены более высокоорганизованными плацентарными млекопитающими и в настоящее время сохранились только в Австралии, Новой Гвинее, Тасмании, Южной Америке, т. е. в южном полушарии, и лишь отчасти в Северной Америке (1 вид) и на острове Сулавеси (1 вид). Только в Австралии, куда из плацентарных проникли лишь рукокрылые и мышевидные грызуны, сумчатые дали большое разнообразие форм зверей, приспособленных к существованию в весьма различной жизненной обстановке.
Высшие, плацентарные млекопитающие, как и сумчатые, произошли от трех- бугорчатых в начале мелового периода и развивались самостоятельно, параллельно с сумчатыми. Первые ископаемые остатки, относимые к насекомоядным, известны из верхнемеловых отложений Монголии. Плацентарные образовали 31 отряд, из которых 17 распространены в настоящее время, а 14 полностью вымерли.
Древнейшим отрядом, давшим, по-видимому, начало всем остальным отрядам, -будут насекомоядные, известные из позднемеловых отложений. Это были мелкие зверьки, ведшие частью наземный, частью древесный образ жизни. Постепенное приспособление некоторых древесных насекомоядных вначале к планированию, а затем и к полету положило начало отряду рукокрылых. Увеличение плотоядности и переход на питание все более крупными животными послужили предпосылкой для формирования в начале третичного времени отряда древних хищников, или креодонтов. Но уже в олигоцене, когда медлительных травоядных начала третичного времени заменили более подвижные формы, креодонты были вытеснены новыми специализированными хищниками типа современных (Carnivora). В олигоцене уже существовали предковые группы вивер, куниц, собак, кошек. В миоцене и плиоцене были обыкновенны оригинальные саблезубые кошки с огромными изогнутыми клыками.
Очень рано, на грани между эоценом и олигоценом, хищники разделились на две неравноценные по обилию видов ветви: большую - наземных хищников и меньшую - ластоногих.
От древних хищников - креодонтов, перешедших на питание растительностью, в самом начале третичного времени (в палеоцене) возникли первичные копытные, или кондилятры. Это были сравнительно мелкие (размером от куницы до волка) всеядные животные с умеренно длинными клыками, бугорчатыми коренными зубами, с пятипалыми конечностями, в которых средний палец был развит сильнее остальных, а первый и пятый пальцы ослаблены. От кондилятр возникли две ветви современных копытных: непарнокопытные и парнокопытные. В эоцене возникли хоботные (корни их происхождения неизвестны). Также неясно происхождение и китообразных. От древних насекомоядных берут начало грызуны, неполнозубые, трубкозубые и приматы.
Наиболее древние ископаемые обезьяны найдены в палеоцене. Древесные обезьяны нижнего олигоцена (проплиопитекусы) дали начало гиббонам и крупным обезьянам из миоцена Индии (рамапитекусам), близким к антропоидам. Остатки человекообразных обезьян (орангутана, гориллы, шимпанзе) известны из плиоценовых отложений Африки, Индии, Китая. Некоторые из них (например, австралопитек, плезиантропус и парантропус) по строению стояли близко к человеку, и какая-то ветвь их дала начало всем его современным расам.
Следует подчеркнуть большую самостоятельность двух основных ветвей в филогении млекопитающих. К одной из них относятся только виды однопроходных (утконос, ехидна и проехидна). Эта ветвь берет начало от упоминавшихся ранее многобугорчатых и никогда не имела большого развития. К другой ветви, корнями уходящей к трехбугорчатым, относятся все остальные млекопитающие (сумчатые и плацентарные).
Современное представление о системе млекопитающих выглядит так:
Класс Млекопитающие (Mammalia)
Подкласс Первозвери (Prototheria)
Отряд Однопроходные (Monotremata)
Подкласс Настоящие звери (Theria)
Инфракласс Низшие звери (Metatheria)
Отряд Сумчатые (Marsupialia)
Инфракласс Высшие звери (Eutheria)
17 отрядов современных и
14 отрядов вымерших.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'