Отряд Миктофообразные (Myctophiformes) (В. Э. Беккер)
К миктофообразным рыбам относятся светящиеся анчоусы, ящероголовы, алепизавры и близкие к ним формы. Миктофообразные близки к сельдеобразным, но сильно специализованы. Брюшные плавники у них расположены в средней части брюха, грудные посажены низко; позади спинного плавника обычно имеется жировой плавник. Рот большой и, в отличие от сельдеобразных, окаймлен сверху одной парой костей (предчелюстными костями), а не двумя. Плавательный пузырь замкнутый или отсутствует. Миктофообразные - самые многочисленные и разнообразные глубоководные рыбы подповерхностного слоя, их насчитывается около 380 видов.
Все миктофообразные - морские, преимущественно глубоководные рыбы, только один вид - бомбилъ - живет в эстуариях. Различают две группы (подотряда) миктофообразных - миктофовидных и алепизавровидных.
Подотряд объединяет 10 семейств придонных и батипелагических рыб, распространенных от прибрежной и шельфовой зоны до больших глубин, порядка 5-6 тыс. м, а в пелагиали открытого океана от поверхности до глубины 2500 м. Наибольшее значение среди придонных семейств имеют ящероголовые и зеленоглазковые, а среди пелагических - светящиеся анчоусы. Это преимущественно бентосоядные или планктоноядные рыбы, размером от 5 до 50-60 см. Тело обычно покрыто хорошо развитой чешуей, скелет более или менее обызвествлен, у многих имеются хорошо развитые специальные светящиеся органы - фотофоры и разного рода светящиеся железы.
Семейство Аулоповые (Aulopidae)
Данное семейство, пожалуй, наименее специализировано в отряде миктофообразных. Аулоповые рыбы близки к светящимся анчоусам, но не имеют светящихся органов и обитают не в толще вод открытого океана, а у берегов, на сравнительно небольших глубинах. Распространены они довольно широко в тропических и умеренно теплых водах; очень обычны в Средиземном море, у восточных берегов Атлантического и западных берегов Тихого океана; известны также из Северо-Западной Атлантики и от побережья Калифорнии. Один вид - химе (Hime japonica) - известен из Японского моря.
Семейство аулоповых содержит один или два рода и около восьми видов. Размеры его наиболее крупных представителей не превышают 50-60 см. Тело этих рыб умеренно вытянуто, сильно сжато к хвосту, но почти круглое в сечении и довольно высокое в передней части, покрыто некрупной чешуей. Голова массивная, с большим ртом, задний край которого достигает середины глаза. На челюстях и нёбных костях имеются многочисленные мелкие зубы. Глаза большие и расположены очень близко к верхнему профилю головы. Спинной плавник обычно большой, его первые лучи иногда удлинены. На верхней стороне хвостового стебля всегда есть маленький жировой плавничок. Анальный плавник отнесен в заднюю часть тела и начинается далеко позади вертикали конца основания спинного. Характерны для аулоповых очень большие и широко расставленные брюшные плавники.
Для некоторых видов семейства характерен половой диморфизм, выражающийся в строении и размерах непарных плавников, особенно передней части спинного: у самцов второй и третий лучи этого плавника удлинены в виде нитей (например, у Aulopus filamentosus).
Как многие другие донные прибрежные рыбы теплых морей, аулоповые окрашены очень пестро и ярко. У химе (Hime japonica) общий тон тела светло-красный, с серебристым блеском, его верхняя половина пересекается темно-коричневыми, а нижняя - ярко-красными полосами и пятнами.
Биология аулоповых рыб не изучена. Судя по строению их рта и зубному вооружению, можно полагать, что питаются они главным образом некрупными донными беспозвоночными и, возможно, молодью других рыб. Обитают эти рыбы у дна на сравнительно небольших глубинах; так, например, уже упоминавшаяся химе живет на глубине 90-110 м в водах, омывающих Японию. У австралийского побережья эти рыбы обычны среди скал и рифов. В некоторых районах (например, в Японии) аулоповые имеют промысловое значение, а в Австралии известны и как объекты спортивного рыболовства. Мясо их вкусно и пригодно в пищу.

Рис. 93. Аулоп (Aulopus nanae)
Семейство Ящероголовые (Synodontidae)
У ящероголовых рыб тело прогонистое, почти цилиндрическое. Над концом анального плавника имеется небольшой жировой плавничок. Довольно большие глаза снабжены вертикальными жировыми веками и расположены у верхнего переднего края головы, очень близко друг к другу. Слегка приплюснутая, с большим ртом, покрытая чешуей голова этих рыб действительно очень похожа на голову ящерицы. Общий тон тела большинства видов светло-коричневый, иногда с зеленоватым или красноватым оттенком; на боках имеются обычно более темные или яркие пятна, широкие поперечные или узкие продольные полосы. Такая окраска делает этих рыб почти незаметными на фоне дна морских мелководий. Если что-либо привлекает внимание лежащей на дне рыбы, она приподнимает голову и переднюю часть тела, готовясь схватить добычу; если же оказывается, что ей грозит опасность, она мгновенно зарывается в песок, оставляя на его поверхности только глаза.
Ящероголовые рыбы широко распространены в тропических и теплых морях всех океанов, но, по-видимому, нигде не бывают особенно многочисленны. Некоторые из них известны с довольно больших глубин, до 300-350 м и более. Большинство видов, однако, обитает у берегов, в районах с илистым или песчаным грунтом, близ скал и рифов. Известны 3 или 4 рода с 33-34 видами. Большинство из них невелики и не превышают в длину 25-30 см. Представители некоторых видов, однако, достигают значительно больших размеров, до 40-50 см.
Особенно широко распространен короткорылый ящероголов (Trachinocephalus myops), встречающийся у западных берегов Тихого океана от Хоккайдо и Гавайских островов до Австралии и островов Океании, в Индийском океане вдоль берегов Южной Азии и Восточной Африки и в Атлантическом океане в водах острова Св. Елены и у островов Центральной Америки и Флориды. Род Зауриды (Saurida, 7 видов) заселяет прибрежные воды индотихоокеанской области, и один из его видов, заурида-эсо (S. undosquamis), недавно проник из Красного моря в Средиземное море. Наибольшее количество видов содержит род длиннорылых, или настоящих, ящероголовов (Synodus), 5 видов которого обитают у берегов Японии, 3 чисто австралийских, 9 живут у западных берегов Америки от Калифорнии до Перу, 5 - у восточных берегов Америки от мыса Код и Флориды до Бразилии и 1 (S. saurus) - у берегов Южной Европы, в Средиземном море. Ящероголовы встречаются как у открытых берегов океана, так и в окраинных морях и в связанных с морем континентальных солоноватых и соленых водоемах, обитая подчас в очень широком диапазоне температуры (от 8 до 32° С) и солености (от 14 до 37 и даже до 600/00) (например, Synodus foetens). Питаются ящероголовы мелкими беспозвоночными и молодью рыб, но некоторые из них ведут хищный образ жизни. Так, например, преимущественно рыбой питается уже упоминавшийся обыкновенный ящероголов (S. foetens), хотя в его рацион входят также мелкие ракообразные и черви. Этот вид переходит на питание рыбой довольно рано, достигнув длины всего 3,5 см. При этом он потребляет и своих собратьев, если они хоть немного уступают ему по величине; например, в желудке малька длиной 4 см был обнаружен трехсантиметровый малек этого же вида.

Рис. 94. Короткорылый ящероголов (Trachinocephalus myops)
Мясо ящероголовов невысоких вкусовых качеств, тем не менее некоторые виды имеют довольно существенное значение в промысле донных рыб в Желтом, Восточно-Китайском и Южно-Китайском морях. Таковы, например, достигающие 40-50 см длины и веса 1,1 кг зауриды: эсо (Saurida undosquamis), тумбилъ (S. tumbil), короткоперая (S. elongata), а также красный ящероголов (Synodus variegatus). Значительная часть японского улова этих рыб перерабатывается в рыбную пасту "камабоко".
Семейство Бомбилевые (Harpodontidae)
Бомбилевые рыбы представлены всего одним родом Бомбиль (Harpodon) с двумя видами, один из которых, по- видимому, глубоководный и известен только из вод, омывающих Японию, а другой обитает в прибрежных районах северной половины Индийского океана от Занзибара до австрало-азиатских морей и в водах Южного Китая.
Индоокеанский бомбилъ (Harpodon nehereus) - небольшая рыба длиной до 40 см. Тело у бомбиля прогонистое, сжатое с боков, в передней части лишенное чешуи, на ощупь мягкое и как бы дряблое. Во внешнем облике этой рыбы характерны закругленный передний профиль головы, огромный рот с многочисленными мелкими, загнутыми назад зубами, длинные грудные и брюшные плавники и необычный
хвостовой плавник, у которого посредине, в выемке между верхней и нижней лопастями, есть маленькая третья лопасть. Голова, спина и бока бомбиля полупрозрачные, светло-серые, с многочисленными звездообразными черными или коричневыми пятнышками. Брюхо светлое, серебристо-голубое. Плавники прозрачные, окрашены так же, как тело, но пятна на них расположены гуще, и от этого они кажутся более темными.
Бомбиль обитает главным образом в предустьевых пространствах моря, в эстуариях, заходит он и в реки. Почти по всей области своего распространения этот вид является важным объектом прибрежного рыболовства, но особенно велико его промысловое значение в дельте Ганга и у западных берегов Индии, в водах штата Бомбей. Бомбиль отличается исключительной жирностью, и его заготавливают впрок главным образом в сушеном виде, причем на западном побережье Индии он высушивается без предварительной засолки и известен под названием "бомбейская утка".
Семейство Батизавровые (Bathysauridae)
В этом семействе только один род Батизавры (Bathysaurus) с тремя видами. Батизавры близки к бомбилям и ящероголовам, но отличаются от них некоторыми особенностями строения и образом жизни. Тело батизавров удлиненное, сжатое с боков, особенно в задней части, а голова, наоборот, уплощена сверху вниз. Рот огромный, с многочисленными острыми и довольно длинными зубами. Глаза овальной формы, небольшие, направлены вверх и вбок и расположены не у переднего края головы, а примерно посредине ее длины. Окраска тела темно-коричневая, снизу почти черная. Светящихся органов нет. Батизавры - одни из самых крупных представителей миктофообразных: они достигают длины более 60 см.
Все батизавры - придонные глубоководные рыбы, живущие на больших глубинах - от 1600 до 4500 м. Распространены батизавры, по-видимому, очень широко; правда, в Индийском океане они до сих пор не обнаружены. Придонный образ жизни и большая глубина обитания явились причиной того, что сейчас, спустя почти 90 лет после описания первого вида, в музеях мира имеются лишь считанные экземпляры батизавров. Их поведение, размножение, питание и другие стороны биологии еще совершенно не известны.
Семейство Зеленоглазковые (Chlorophthalmidae)
В состав семейства входит три рода и около 20 видов морских донных рыб. Большинство видов, принадлежащих к родам Зеленоглазки (Chlorophthalmus) и парасудис (Parasudis), населяют прибрежные районы, где глубины составляют от 50 до 200 м и более, а три вида рода Батизayропс (Bathysauropsis) обитают на глубине более 2000 м. Этот глубоководный род является как бы промежуточным звеном между умеренно глубоководными зеленоглазками (Chlorophthalmus) и близкими глубоководными семействами ипноповых и батиптеровых.

Рис. 95. Бразильская зеленоглазка (Chlorophthalmus brasiliensis)
У всех зеленоглазок тело удлиненное, почти круглое в передней части и сжатое с боков к хвосту. Голова, в особенности рыло, удлинены и немного сплюснуты. Характерны очень большие зеленые глаза, смотрящие вбок и немного вверх. Все зеленоглазковые - гермафродиты.
Обитающая у американского берега Атлантического океана и в Средиземном море обыкновенная зеленоглазка (Chi. agassizi) держится в тех местах, где преобладают илистые грунты или мелкий песок, и питается мелкими донными беспозвоночными. Она распространена довольно широко, но не совершает дальних миграций. Поэтому на небольшом удалении друг от друга обитают немного различающиеся популяции этого вида, которые не смешиваются между собой. У берегов Японии обитает 5 видов зеленоглазок, в том числе один в Японском море. Другой прибрежный представитель семейства, обыкновенный парасудис (P. truculentus), достигающий длины 20-25 см, питается мелкой рыбой и в том числе обитающими в средних слоях воды видами светящихся анчоусов (семейство Myctophidae) и гоностомовых (семейство Gonostomidae). У восточного побережья Северной Америки этот вид, по-видимому, очень многочислен. Во всяком случае нередко за час траления в донный 12-метровый трал попадают сотни экземпляров этого вида. Специального промыслового значения он, однако, не имеет и попадается главным образом при ловле креветок.
Семейство Скопелозавровые (Scopelosauridae)
Это малоизученное семейство океанических рыб, содержащее только один род Скопелозавры (Scopelosaurus), в котором имеется около десяти видов. Внешне скопелозавровые напоминают маленьких барракуд и очень похожи на веретенниковых рыб (сем. Paralepididae). В действительности, однако, они наиболее близки к зеленоглазкам (сем. Chlorophthalmidae), от которых отличаются, кроме некоторых черт строения, тем, что ведут не придонный, а пелагический образ жизни.
Тело скопелозавровых рыб удлиненное, покрыто довольно крупной циклоидной чешуей; как и у многих близких форм, оно почти круглое в передней части и сжато с боков в задней. Спинной и брюшные плавники располагаются в средней части тела; анальный и маленький жировой плавничок отнесены далеко назад. Голова большая, превышает 1/4 длины тела, сужается кпереди. Челюсти и некоторые нёбные кости вооружены многочисленными мелкими и острыми зубами. Глаза большие; зрачок, так же как и все глазное яблоко, имеет овальную форму. Светящихся органов и плавательного пузыря у скопелозавровых рыб нет.
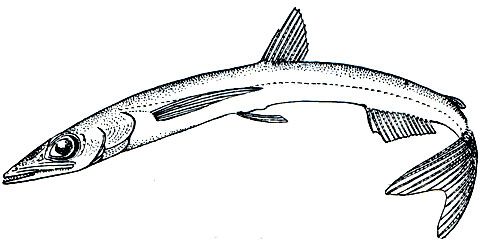
Рис. 96. Скопелозавр (Scopelosaurus lepidus)
Только один из известных взрослых экземпляров скопелозавров достигает длины 50 см; размеры остальных половозрелых рыб разных видов этого семейства не превышают 20 см. Большинство же скопелозавров, пойманных в различных экспедициях, неполовозрелы, а длина их составляет всего 3-5 см. Причина преобладания в уловах мелких рыб, видимо, состоит в том, что взрослые скопелозавры очень подвижны и стремительны и без труда уходят от любых орудий лова.
Темная окраска и строение глаз этих рыб позволяют полагать, что они живут глубже 1000 м, однако прямых доказательств этому очень мало. Только дважды представители этого семейства были пойманы сетями, которые поднимались с глубины более 1000 м. Большинство же скопелозавров поймано при ловах от 600 м и меньше. Несколько экземпляров были найдены на пляжах, а один засосан на палубу судна вместе с забортной водой. Все это говорит о том, что представители этого семейства могут быть встречены и у поверхности моря.
Распространены скопелозавровые очень широко. Они встречаются во всех трех океанах и известны от Исландии на севере до острова Маккуори на юге. Но наиболее часто они встречаются в умеренных и тропических водах.
Семейство Батиптеровые (Bathypteroidae)
Представители этого семейства небольшие, длиной всего 10-35 см, обитатели придонных слоев воды, близки к ипноповым, особенно к роду слепых рыб ипнопсов (Ipnops). В отличие от ипноповых, однако, у всех батиптеровых имеются нормально развитые, хотя и очень маленькие, глаза, расположенные по бокам головы, сразу над верхней челюстью. Тело у батиптеровых прогонистое, в задней части сжатое с боков, в передней - почти круглое. Общий тон его окраски темно-коричневый, до черного, причем спина и голова обычно окрашены темнее. Плавники темные или бесцветные. У некоторых видов края кожных карманов, в которых сидят чешуйки, белые. Наиболее характерная особенность внешнего облика батиптеровых - необычайно длинные лучи плавников. У разных видов в разных комбинациях бывают удлинены верхние или нижние лучи грудных плавников, наружные брюшных и нижние лучи хвостового плавника. У бентозавра (Benthosaurus grallator) чрезвычайно длинные лучи брюшных и хвостового плавников (длина которых превышает длину тела) образуют треножник, на который рыба опирается, опускаясь на дно. Именно в таком виде бентозавр и был сфотографирован из иллюминатора батискафа FNRS-3 на дне Средиземного моря. У некоторых видов рода батиптеров удлиненные верхние лучи грудных плавников при плавании направлены вперед и, по-видимому, служат органами осязания.

Рис. 97. Батиптер (Bathypterois atricolor)
Другая интересная особенность батиптеровых - их обоеполость (гермафродитизм). У некоторых видов в теле каждой рыбы молоки (сперма) и икра созревают одновременно, однако не известно, происходит ли при этом самооплодотворение или же икринки одной особи оплодотворяются молоками другой. Ничего не известно и о ранних стадиях их развития.
Семейство батиптеровых включает всего два рода - бентозавров (Benthosaurus) и батиптеров (Bathypterois) - и широко распространено в пределах тропической и умеренно теплой областей Мирового океана. Обитают батиптеровые у дна и, по-видимому, предпочитают мягкие, илистые грунты. Глубины, на которых живут разные виды, различны, но известные пределы вертикального распространения семейства в целом очень широки - от 480 до 5600 м.
Семейство Ипноповые (Ipnopidae)
Все представители этого семейства - морские, донные, глубоководные рыбы. Длина взрослых особей разных видов от 10 до 40 см. Наименьшая глубина, на которой они были пойманы, составляет около 800 м, наибольшая - почти 6000 м, причем большая часть нахождений приходится на глубины, превышающие 2000 м. Внешне ипноповые похожи на батиптеровых (семейство Bathypteroidae) и, вероятно, близки к ним. Тело у них удлиненное и покрыто чешуей. Общий тон его окраски темно-коричневый. Передняя часть головы, как и у батиптеровых, вытянута и сплюснута. Все плавники развиты хорошо; нижние лучи хвостового у некоторых видов удлинены. Светящихся органов нет. Наиболее яркая отличительная особенность ипноповых - необычайные изменения органов зрения. Глаза у представителей этого семейства либо вовсе отсутствуют (Ipnops), либо очень малы и скрыты под чешуей и кожей (Bathymicrops) и, конечно, не могут функционировать так, как глаза большинства других рыб.

Рис. 98. Ипнопс (Ipnops murrayi)
Верхняя сторона головы у видов рода Ипнопс (Ipnops) покрыта двумя очень тонкими костными пластинками. Под ними лежат так называемые головные органы, функции которых долгое время не были ясны. Первоначально полагали, что это светящиеся органы. Затем предполагалось, что это органы зрения. Но лишь недавно изучение тонкого строения головных органов ипнопса показало, что они представляют собой совершенно уникальные светочувствительные структуры, которые, как и глаза других позвоночных животных, имеют сетчатку, но, в отличие от них, лишены хрусталика. У представителей двух других родов глаза расположены по бокам головы, близко к ее переднему краю и, как уже говорилось, чрезвычайно малы и не способны воспринимать свет.
Эти и некоторые другие особенности строения заставляют думать, что ипноповые ведут необычный образ жизни. Но на этот счет сейчас можно лишь строить догадки, так как материал не слишком велик. Судя по опубликованным работам, до сего времени известно всего около 40 экземпляров этих рыб. Около 30 из них приходится на три вида рода Ipnops, а остальные представляют роды Bathymicrops с одним и Bathytyphlops с двумя видами. Можно сказать, что семейство ипноповых представляет собой одну из многочисленных загадок, которые еще предстоит решить морским ихтиологам.
Семейство Неоскопеловые (Neoscopeiidae)
Семейство содержит только три рода; два из них, Neoscopelus и Scopelengys, включают по два или три вида, а один, Solivomer,- монотипический. До недавнего времени эти рыбы в качестве наиболее примитивной группы относились к светящимся анчоусам (сем. Myctophidae), но теперь выделяются в самостоятельное семейство. Оснований к этому вполне достаточно: неоскопеловым присущ иной внешний облик и ряд существенных отличий в строении скелета. Кроме того, у двух родов - Скопеленгисы (Scopelengys) и Соливомеры (Solivomer) - полностью отсутствуют фотофоры. У видов третьего рода - Неоскопелы (Neoscopelus) - фотофоры, правда, имеются, но расположены они непрерывными рядами вдоль брюха, а не обособленными группами, как у настоящих светящихся анчоусов. Считается, что среди миктофовидных (Myctophoidei) неоскопеловые занимают промежуточное положение между миктофовыми (Myctophidae) и зеленоглазками (Chlorophthalmidae). От последних они отличаются большим ртом и наличием плавательного пузыря.
Тело неоскопеловых рыб умеренно вытянутое, с довольно массивной головой. Рот большой и заходит за вертикаль заднего края глаза. Зубы на челюстях очень мелкие. Характерно для неоскопеловых рыб положение анального плавника, который у них начинается позади конца основания спинного. У всех видов хорошо развит жировой плавничок. Длина наиболее крупных экземпляров немного более 20 см.
Фотофоры у неоскопелов устроены иначе, чем у светящихся анчоусов. Они имеют овальную форму и лежат на поверхности брюшных мышц. Светящаяся ткань и окружающий ее черный пигмент находятся в задней части органа, а его передняя часть представляет собой просто блестящий участок гуанина, прикрытого прозрачной тканью. Впереди фотофоры не имеют четкой границы.
Биология неоскопеловых рыб не изучена. Известно только, что они ведут пелагический образ жизни на значительных глубинах, от 500 до 2000 м, встречаясь иногда стаями. Представители этого семейства известны из Тихого, Индийского и Атлантического океанов, примерно от 35° с. ш. до 35° ю. ш.
Семейство Миктофовые, или Светящиеся анчоусы (Myctophidae)
Семейство светящихся анчоусов, или миктофовых, содержит 30 родов и около 200 видов и является, таким образом, наиболее крупным в подотряде миктофовидных (Myctophoidei) и одним из самых больших семейств глубоководных костистых рыб вообще. Светящиеся анчоусы необычайно широко распространены в Мировом океане - разные виды этой группы известны от Шпицбергена и северной части Берингова моря до шельфовых ледников Антарктиды. Наконец, многие виды семейства достигают огромной численности и представляют определенный практический интерес. Все это делает светящихся анчоусов чрезвычайно перспективным и многообещающим объектом исследования, так как на их примере могут быть выяснены общие черты биологии и закономерности распространения, присущие многочисленным обитателям средних слоев пелагиали Мирового океана.
Светящиеся анчоусы - небольшие рыбки, обычной "рыбообразной" формы.

Таблица 24. Кефалеобразные и атерины: 1 - большая барракуда (Sphyraena barracuda); 2 - атерина черноморская (Atherina pontica); 3 - атерина-изо (Iso natalensis); 4 - лобан (Mugil cephalus); 5 - индийская кефаль (Ellochelon vaigiensis); 6 - пальцепер-дара (Polydactylus indicus); 7 - пятипалый пальцепер (Polynemus quinquarius); 8 - обыкновенный пальцепер (P. Plebeius); 9 - многопалый пальцепер (Galeoides polydactylus)

Таблица 24. Кефалеобразные и атерины: 1 - большая барракуда (Sphyraena barracuda); 2 - атерина черноморская (Atherina pontica); 3 - атерина-изо (Iso natalensis); 4 - лобан (Mugil cephalus); 5 - индийская кефаль (Ellochelon vaigiensis); 6 - пальцепер-дара (Polydactylus indicus); 7 - пятипалый пальцепер (Polynemus quinquarius); 8 - обыкновенный пальцепер (P. Plebeius); 9 - многопалый пальцепер (Galeoides polydactylus)
Взрослые особи самых мелких видов - нотолихна (Notolychnus valdiviae), диогенихта (Diogenichthys atlanticus) имеют длину всего 2,5-3 см, а длина наиболее крупных экземпляров некоторых видов рода Лампаникты (Lampanyctus) ирода Нотоскопелы (Notoscopelum) не превышает 20-30 см. Все тело этих рыб покрыто довольно крупной чешуей, как правило, циклоидной, и только у нескольких видов имеется ктеноидная чешуя с более или менее развитыми шипикамипо заднему краю. Боковая линия обычно хорошо развита, но у видов примитивного рода Протомиктофы (Protomyctophum) и видов рода Центробранх (Centrobranchus) она отсутствует. Плавники довольно большие, и только у некоторых видов рода Лампаникт (Lampanyctus) грудные плавники очень малы, а у одного из них (L. achirus) вовсе отсутствуют. Анальный плавник начинается под концом основания спинного или сразу за ним. На верхней стороне хвостового стебля, над концом анального плавника, всегда имеется хорошо развитый жировой плавничок.
Голова у светящихся анчоусов довольно большая, ее длина составляет от одной четверти до одной трети длины тела (без хвостового плавника). Размер глаз у этих рыб значительно варьирует, от средней величины до очень больших, почти до трети длины головы. Несколько видов рода Протомиктоф (Protomyctophum) отличаются так называемым "полутелескопическим" строением глаз (у них зрачок смещен вверх). Вооруженный многочисленными мелкими зубами рот светящихся анчоусов простирается за задний край глаза. Именно большой рот этих рыб, похожий на рот настоящих анчоусов (семейство Engraulidae), и послужил поводом для их названия, хотя в действительности эти две группы довольно далеки друг от друга и принадлежат к разным отрядам.
Наиболее характерной чертой внешнего облика светящихся анчоусов являются различного рода светящиеся органы. По местоположению и строению их можно разделить на четыре типа: фотофоры и хвостовые, окологлазничные и туловищные железы. Наиболее интересно и сложно устроены фотофоры. В отличие от таких же органов неоскопелов (семейство Neoscopelidae) каждый фотофор светящихся анчоусов помещается в чашеобразной впадине мышц, резко отграничен от окружающих тканей, имеет самостоятельное кровоснабжение и обслуживается специальным нервом. Со стороны тела фотофор подстилается слоем черного пигмента, который немного выступает за края органа и на поверхности тела окружает его сплошным черным кольцом. На пигментной выстилке лежит блестящий рефлектирующий слой. Затем следуют студенистая ткань и продуцирующие свет железистые клетки. Снаружи фотофор прикрыт измененной в виде линзы чешуйкой. Под линзой находится полулунная шторка - септа (особенно хорошо заметная у диафов - видов рода Diaphus), с помощью которой рыба, по-видимому, может произвольно изменять силу и направление света. Располагаются фотофоры почти у всех видов семейства только на нижней половине тела, ниже боковой линии, и образуют вполне определенные группы, каждая из которых имеет свое название. Число фотофоров в группах и их взаимоположение специфичны для вида и, вероятно, играют определенную роль в поведении этих рыб. Кроме только что описанных "основных" фотофоров, у нескольких видов имеются и так называемые "вторичные" фотофоры, располагающиеся на каждой чешуйке или только на некоторых из них. Размеры "вторичных" фотофоров, как правило, меньше, чем "основных", и лишь у скопелопсиса (Scopelopsis multipunctatus) величина тех и других органов примерно одинакова.

Таблица 25. Центрарховые, Каталуфовые, Пемферовые: 1, 2, 3 - форелеокунь (Micropterus salmoides); 4 - амблоплитес (Ambloplites rupestris); 5 - помоксис, или краппи (Pomoxis annularis); 6 - солнечная рыба (Lepomis gibbosus); 7 - пристигенис (Pristigenys niphonia); 8 - каталуфа (Priacanthus arenatus); 9 - японский приакант (P. macracanthus); 10 - большеглаз атлантический (Pempheris schomburgki); 11 - лептобрама (Leptobrama milleri)

Таблица 25. Центрарховые, Каталуфовые, Пемферовые: 1, 2, 3 - форелеокунь (Micropterus salmoides); 4 - амблоплитес (Ambloplites rupestris); 5 - помоксис, или краппи (Pomoxis annularis); 6 - солнечная рыба (Lepomis gibbosus); 7 - пристигенис (Pristigenys niphonia); 8 - каталуфа (Priacanthus arenatus); 9 - японский приакант (P. macracanthus); 10 - большеглаз атлантический (Pempheris schomburgki); 11 - лептобрама (Leptobrama milleri)
Хвостовые светящиеся железы встречаются у многих видов семейства и могут иметь различное строение. Иногда это сложные органы, имеющие, как и фотофоры, черную пигментную выстилку, рефлектор и линзоподобные структуры. В таких случаях хвостовые железы играют роль вторичных половых признаков, причем у самцов они обычно устроены сложнее и помещаются на верхней стороне хвостового стебля, а у самок их строение проще и располагаются они на нижней стороне тела, между анальным и хвостовым плавниками. Такие железы отсутствуют у мальков и развиваются только ко времени наступления половой зрелости. У других видов (рода Lampanyctus и близких к нему) хвостовые железы представляют собой простые белые или желтоватые чешуевидные пластинки, лежащие на поверхности тела. В таких случаях они одинаковы у рыб разных полов и появляются во время развития очень рано, одновременно с появлением фотофоров.
Окологлазничные светящиеся железы особенно характерны для диафов (видов рода Diaphus), хотя имеются и у многих других представителей семейства. Строение этих желез у разных видов диафусов различно и часто служит хорошим признаком для их определения. Величина их также значительно варьирует: от маленьких, округлых, не более фотофора, железок до огромных и сложных органов, занимающих всю предглазничную область головы. Интересно, что у диафов, которые не имеют хвостовых светящихся желез, половой диморфизм выражен в строении и величине окологлазничных светящихся органов (у самок многих видов они меньше, чем у самцов).
Туловищные светящиеся железы представляют собой участки светящейся ткани без каких-либо дополнительных устройств вроде рефлектора или линзы. Располагаются такие железы на поверхности тела (и очень легко утрачиваются, не оставляя, в отличие от фотофоров, никаких следов), обычно вдоль оснований плавников и реже между ними, вдоль спины или брюшка. Туловищные железы особенно характерны для высокоорганизованных форм семейства, таких, как лампаникты (Lampanyctus) и близкие к ним.
Продолжительность жизни светящихся анчоусов невелика. Особи сравнительно крупного светлого лампаникта (Stenobrachius leucopsarus), достигающего длины около 10 см, живут всего 3, редко 4 года. Срок жизни более мелких форм (Notolychnus, Diogenichthys, Centrobranchus и т. д.), вероятно, еще меньше.
Светящиеся анчоусы всю свою жизнь проводят в толще вод открытого океана. В тропической области их размножение происходит в течение круглого года. В субтропических и умеренных водах имеет место, по крайней мере для некоторых видов, сезонность размножения. Мелкие личинки светящихся анчоусов держатся у поверхности моря и входят в состав поверхностного планктона. По мере роста, с переходом к мальковой фазе развития, молодь опускается в более глубокие слои воды и постепенно переходит к образу жизни взрослых рыб. Плодовитость светящихся анчоусов довольно велика, и самки длиной около 7 см выметывают 7-8 тыс. икринок.
Пищу светящихся анчоусов составляют преимущественно планктонные ракообразные, но иногда мальки и личинки других рыб. Некоторые виды (например, Ceratoscopelus townsendi) питаются, по-видимому, только ночью, поднимаясь в верхние слои воды. Хотя светящиеся анчоусы приспособлены к питанию мелкими организмами, иногда они заглатывают и довольно крупную добычу. Так, например, однажды в желудке пойманного ночью у поверхности моря диафа (Diaphus dumerili, длиной около 8 см) был обнаружен малек летучей рыбы Exocoetus, длиной около 4 см. Правда, малек этот был несомненно ослаблен, так как на нем сидел крупный паразитический рачок из отряда равноногих раков (Isopoda).
Массовые виды светящихся анчоусов в свою очередь служат важными объектами питания более крупных рыб (лососей, морских окуней, дальневосточной трески, отчасти некоторых тунцов и марлинов), а также морских котиков и китов. Один из видов рода нотоскопелов (Notoscopelus) составляет в пище котиков на местах их зимовки в водах Японии около 60% всего их рациона. Примерно столько же составляют они и в питании дальневосточных лососей в районах их откорма.
Светящиеся анчоусы населяют преимущественно верхний тысячеметровый слой вод океана и на глубинах более 1000 м уступают свое место наиболее многочисленной группе - циклотонам (Cyclothone, семейство Gonostomidae). Среди них есть, однако, виды, которые, насколько сейчас известно, не встречаются выше 700- 800 м (Taaningichthys bathyphilus). По наблюдениям с французского батискафа FNRS-3 в Средиземном море, светящиеся анчоусы встречаются до глубины 2300 м (до максимальной глубины этого погружения).
По-видимому, большинство светящихся анчоусов совершают суточные вертикальные миграции и в темное время суток поднимаются в верхние слои воды, а с рассветом покидают их и уходят на глубину. В этом отношении семейство может быть разделено на две группы. Одна из них включает 30-40 видов так называемых "приповерхностных" светящихся анчоусов (роды Myctophum, Symbolophorus, Gonichthys и др.). Характерной особенностью этих видов является то, что они в темное время суток поднимаются к самой поверхности воды и, таким образом, ведут в это время отнюдь не глубоководный образ жизни. На этом основании их следует рассматривать в качестве временного компонента приповерхностного комплекса рыб (постоянными членами которого являются летучие рыбы, корифены, тунцы и т. д.). Все приповерхностные виды обладают положительной реакцией на свет и привлекаются его искусственными источниками. В свете лампы, опущенной тропической ночью с борта судна, почти всегда можно наблюдать этих стремительных, сверкающих серебром рыбок, преследующих добычу или преследуемых кальмарами или гемпилами (змеиными макрелями).

Рис. 99. Диаф (Diaphus liitkeni)
Вторая группа светящихся анчоусов объединяет остальные виды семейства. Они в ночное время также поднимаются на меньшие глубины, но, как правило, не достигают поверхности моря. Известно, что многие из этих видов входят в состав населения звукорассеивающих слоев и мигрируют вместе с ними.
В связи с различиями в глубине обитания светящихся анчоусов изменяются и некоторые черты их строения. Особенно интересны в этом отношении изменения размеров испускающих и воспринимающих свет органов, т. е. фотофоров и глаз. Оказывается, что самые крупные глаза и фотофоры присущи не наиболее глубоководным, а, напротив, поднимающимся к поверхности видам. С увеличением глубины обитания и те и другие органы закономерно уменьшаются. То же явление известно и для других батипелагических рыб. Исчерпывающего объяснения ему пока не найдено. По-видимому, прямой связи между изменениями величины фотофоров и глаз не существует.
Если размеры фотофоров пропорциональны яркости свечения и если верно, что они создают "противотеневой эффект" (об этом подробнее говорилось в очерке о семействе гоностомовых), тогда можно полагать, что уменьшение их величины у более глубоководных форм является прямым следствием уменьшения яркости верхнего фона в результате поглощения дневного света в толще воды. Понятно, что для создания "противотеневого эффекта" на менее ярком фоне нужны менее яркие источники света.
Как уже говорилось, светящиеся анчоусы представляют собой одну из наиболее широкораспространенных групп глубоководных пелагических рыб. Наибольшего разнообразия они, однако, достигают в субтропических и тропических водах. Огромное большинство видов семейства встречается между 40-45-й параллелями северного и южного полушарий. К югу от 45° ю. ш. постоянно обитает около 20 видов и примерно столько же - к северу от 45° с. ш. В Северной Атлантике, однако, с мощным и теплым потоком Северо-Атлантического течения многие тепловодные виды проникают и дальше на север, почти до 60° с. ш. Вообще же распространение отдельных видов светящихся анчоусов (как и других мелких глубоководных пелагических рыб) связано с водами определенных структур. Так, например, в Атлантическом океане распространение нескольких видов этого семейства очень четко ограничено водами северного центрального круговорота, образуемого Гольфстримом, Канарским, Северным пассатным и Антильским течениями. Та же картина наблюдается и в северной части Тихого океана, где целый ряд видов обитает в круговороте, образуемом Куро-Сиво, его продолжением и Северным пассатным течением. В средних и высоких широтах, где основные течения имеют широтное направление, ареалы светящихся анчоусов также вытянуты в широтном направлении, и их распространение в этих районах носит ярко выраженный зональный характер. Границы распространения некоторых видов светящихся анчоусов оказываются настолько четкими, что по ним могут быть обозначены биогеографические зоны. Так, например, в южной части Тихого океана антарктическую зону населяет один вид рода - антарктическая электрона (Electrona antarctica), а следующую к северу соседнюю нотальную золу - другой вид (Е. subaspera).
К алепизавровидным относятся батипелагические миктофообразные рыбы, существенно отличающиеся от миктофовидных своим строением и биологией. Это хищные рыбы, распространенные в открытом океане от поверхности до глубины 2500 м. Некоторые виды достигают крупной величины, до 150 см и более. Многие виды обладают длинными кинжалообразными зубами. У большинства нет светящихся органов, кожа голая или чешуйный покров слабо развит. Скелет слабообызвествленный. Плавательного пузыря нет.
К этой группе рыб принадлежат 6 семейств, важнейшие из которых веретенниковые и алепизавровые.
Семейство Веретенниковые (Paralepididae)
Веретенниковые - второе по величине после светящихся анчоусов семейство миктофообразных: оно включает 11 родов и около 50 видов. Наиболее древние ископаемые предки веретенниковых очень похожи на современных зеленоглазковых (Chlorophthalmidae). Из числа ныне живущих рыб веретенниковые (и особенно род Stemonosudis) наиболее близки к кинжалозубам (семейство Anotopteridae).
Все представители семейства - настоящие океанические рыбы; некоторые из них достигают длины почти 90 см. Тело этих рыб сильно удлиненное, у некоторых почти угреобразное, спереди сжатое, в задней части овальное или цилиндрическое. Голова большая, уплощенная. Глаза нормального строения, круглые, расположены по бокам головы. Челюсти вытянутые, приостренные. Длина их может достигать половины длины головы. Зубы, подвижные и неподвижные, на челюстях, нёбных костях и жаберных дугах. Самые длинные зубы находятся на нижней челюсти и (подвижные) на передней части нёба.
Спинной плавник маленький, расположен, так же как и брюшной, примерно на середине длины тела. Анальный плавник отодвинут в заднюю часть тела; основание его длинное, примерно равно длине головы. Жировой плавник всегда имеется, развит хорошо и находится над задней частью анального. Примитивные формы семейства, например род Паралепис (Paralepis), имеют чешуйный покров, у более специализированных родов (Lestidium, Lestrolepis и др.) чешуя отсутствует. Плавательного пузыря у всех веретенниковых нет.
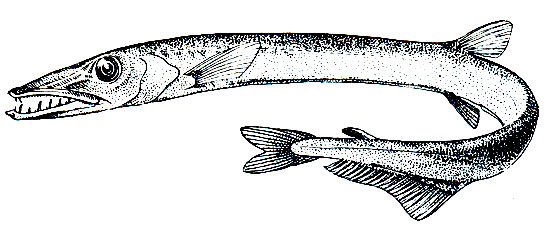
Рис. 100. Веретенник (Lestidiops mirabilis)
Светящиеся органы имеются только у представителей родов Лестидий (Lestidium) и Лестролепис (Lestrolepis). По своему строению они резко отличаются от того, что имеется у светящихся анчоусов или гоностомовых. Светящиеся органы веретенниковых представляют собой внутримышечные каналы, расположенные в брюшной мускулатуре, вдоль средней линии тела между горлом и основанием брюшных плавников. У лестролеписов таких каналов два, у лестидиев - один.
Окраска веретенниковых бывает двух типов. Все чешуйчатые виды имеют довольно толстый слой серебристого гуанина, покрывающего чешую, под которой лежит слой темных пигментных клеток. При жизни тело этих рыб блестит серебром или отливает металлической медью и очень сильно рефлектирует. У бесчешуйных видов тело кажется просвечивающим, зеркально гладкое и переливается всеми цветами радуги. У личинок и молоди некоторых видов сквозь стенки тела просвечивают темноокрашенные участки выстилки брюшной полости, и передняя часть их туловища кажется разделенной на темные сегменты числом до тридцати.
Веретенниковые известны главным образом по молодым, неполовозрелым экземплярам; некоторые роды известны только по молоди. Причина этого состоит, как и для некоторых других глубоководных пелагических рыб, в том, что способные к стремительным броскам крупные рыбы легко избегают медленно движущихся сетей и тралов, которыми работают с научно-исследовательских судов. Достаточно сказать, что только один из известных науке крупных взрослых экземпляров веретенниковых был пойман сетью, а остальные найдены мертвыми или обнаружены в желудках более крупных хищников.
Наиболее удивительная черта биологии веретенниковых - их необычный способ плавания. Во время одного из погружений батискафа FNRS-3 наблюдали на глубине от 600 до 1000 м множество этих рыб, плывших в вертикальном положении. При этом отдельные рыбы могли почти мгновенно переворачиваться, оказываясь то вверх, то вниз головой. Есть основание полагать, что все бесчешуйные виды плавают в вертикальном положении, а все покрытые чешуей - горизонтально. Известно при этом, что у чешуйчатых, примитивных видов спина немного горбата, а голова опущена книзу, т. е. имеются такие особенности в строении тела, которые облегчают переход из горизонтального положения в вертикальное, головой вниз.
Тропические веретенниковые размножаются, по-видимому, круглый год, а обитающие в средних и высоких широтах - весной и в начале лета. Личинки и мальки держатся на меньших глубинах, чем взрослые особи тех же видов.
По характеру питания все веретенниковые - хищники, поедающие мелких рыб, креветок и других беспозвоночных животных. Будучи очень многочисленными, веретенниковые в свою очередь служат пищей более крупным хищникам. Ими питаются дельфины, гренландский тюлень, тунцы, алепизавры, треска и другие крупные рыбы. Однажды в желудке сайды (Pollachius virens) было обнаружено 40 довольно крупных экземпляров веретенника - нотолеписа (Notolepis rissoi). Среди видов этого семейства есть и такие, которые непосредственно используются человеком,- случай для глубоководных пелагических рыб пока еще чрезвычайно редкий. Весной у берегов Юго-Западной Гренландии местные жители собирают довольно много плавающих на поверхности полуживых и уже погибших паралеписов (Paralepis coregonoides) и нотолеписов (Notolepis rissoi), которых они называют "большой песчаный угорь" ("песчаным угрем" они называют песчанку, Ammodytes). Эти рыбы выносятся за пределы своего ареала ответвлениями Северо-Атлантического течения и, попав в холодные воды, погибают.
Распространены веретенниковые чрезвычайно широко и встречаются от Арктики до Антарктики, но наиболее обычны в тропиках. Среди них есть виды с циркумтропическим и даже всесветным распространением (Paralepis atlantica), но есть и такие, распространение которых ограничено определенными районами. Так, например, антарктический нотолепис (Notolepis coatsi) обитает только в Антарктике, а средиземноморский паралепис (Paralepis speciosa) - только в Средиземном море. Некоторые виды очень обычны над шельфом и в закрытых морях, таких, как Средиземное, Мексиканский залив, внутренние индонезийские моря.
Многие виды семейства обычны у поверхности, другие обитают глубже, но ниже 1000-1500 м они не встречаются.
Семейство Кинжалозубые (Anotopteridae)
Семейство кинжалозубых содержит всего один род и вид - кинжалозуб (Аnotopterus pharao). Латинское название рода указывает на исключительную особенность внешнего облика этой рыбы: отсутствие спинного плавника (от греческого, в латинской транскрипции: а - частица отрицания, notos - спина, pteron - крыло или плавник). Этот признак в сочетании с длинным, змеевидным телом, большой головой и громадными клювообразными челюстями настолько характерен, что позволяет безошибочно отличать кинжалозуба от всех других глубоководных пелагических рыб.
Голова и передняя часть тела кинжалозуба сжаты с боков, задняя часть туловища в сечении почти круглая. Чешуи и светящихся органов нет. Кожа гладкая, тонкая. По бокам хвостового стебля у крупных взрослых экземпляров располагаются продольные кожистые кили. Плавники небольшие; анальный - в хвостовой части тела; над ним, на верхней стороне хвостового стебля, имеется небольшой жировой плавничок. Рот огромный - длина челюстей составляет 3/4 длины головы; нижняя челюсть немного длиннее верхней и выступает вперед коническим усиком. Удлинение нижней челюсти не имеет костного основания, и предполагается, что оно служит органом осязания. Самые крупные зубы находятся на нёбных костях; на челюстях, особенно на верхней, зубы мельче и некоторые из них подвижны. Окраска тела медно-серебристая, с радужным отливом, спина темная, у взрослых рыб почти черная. Наиболее крупный кинжалозуб был пойман у Алеутских островов; его длина составляла почти 90 см.
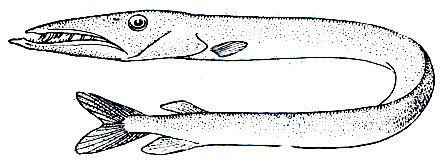
Рис. 101. Кинжалозуб (Anotopterus pharao)
Кинжалозубы населяют средние глубины океана, но встречаются и у поверхности. О их биологии известно очень немногое. И это понятно, так как до сих пор известно всего около 35 экземпляров этого вида. Можно полагать, что кинжалозуб способен к быстрым, резким движениям и именно это его свойство (а не малочисленность) обусловливает его редкость в коллекциях. Возможно также, что он, подобно своим ближайшим родичам из семейства веретенниковых (Paralepididae), плавает в вертикальном положении. О размножении кинжалозуба неизвестно ничего, но, поскольку крупные взрослые рыбы встречаются в более холодных районах, чем молодые, можно полагать, что нерест его происходит в умеренно теплых водах.
По характеру питания кинжалозуб, несомненно, хищник, поедающий других рыб. Известен случай, когда в его желудке были обнаружены два крупных экземпляра из веретенниковых рыб (Notolepis coatsi). Сам кинжалозуб становится жертвой других пелагических хищников: его находили в желудках алепизавров, альбакоров и антарктических китов.
Ареал кинжалозуба является хорошим примером биполярного распространения. Этот вид известен из северных частей Атлантического и Тихого океанов и из антарктических и субантарктических вод, а в пределах тропической области отсутствует.
Семейство Алепизавровые (Alepisauridae)
К семейству алепизавровых принадлежит только один род Алепизавр (Alepisaurus) с двумя или более видами. Его представители наиболее близки к омосудовым (Omosudidae) и кинжалозубам (Anotopteridae), отличаясь от них в основном строением скелета головы, необычайно большим спинным плавником, а от первого из них также удлиненным телом. Алепизавры - крупные, до 2 м, хищные рыбы, обитающие в пелагиали открытого океана. Встречаются они и у поверхности, и на глубине 200 м и более. Географическое распространение их очень широко: эти рыбы отсутствуют, по-видимому, только в полярных бассейнах.
Наиболее характерны во внешнем облике алепизавров слабое, вялое, лишенное чешуи змеевидное тело и громадный спинной плавник. В передней части туловище немного сжато с боков, в хвостовой - почти цилиндрическое, по бокам его с каждой стороны проходит продольный жировой киль. Голова также несколько сжата с боков, при закрытом рте имеет клиновидную форму. Рот огромный, простирается далеко за вертикаль заднего края глаза. Кроме зубов на челюстях, имеются мощные кинжаловидные клыки на нёбных костях. Плавательного пузыря и светящихся органов алепизавры не имеют.
Голова, тело и некоторые плавники алепизавров имеют очень красивый металлический отблеск. Общий тон их окраски серо-сине-черный. Спина почти черная, бока зеленовато-синие, брюхо серебристо-серое. Большой спинной плавник черно-синий. Анальный - черный, с радужным отливом и узкой белой каймой. Все тело рыбы кажется просвечивающим.
Наиболее крупный из известных экземпляров имел длину почти 210 см, а рыбы длиной 1-1,5 м встречаются очень часто в уловах тунцовых ярусов. Интересно, что в различных коллекциях алепизавры представлены именно такими крупными экземплярами; рыбы менее 40 см встречаются в них гораздо реже, и получены они главным образом из желудков других, более крупных особей своего же вида. Алепизавры - весьма прожорливые хищники, и желудки пойманных рыб обычно бывают наполнены самой разнообразной пищей: здесь можно встретить рыб, осьминогов, кальмаров, сальп, различных ракообразных. Благодаря тому что у алепизавров (так же как у омосудиса) переваривание пищи происходит только в кишечнике, содержимое их желудков представляет собой интереснейший материал для ихтиологов-систематиков. И немало новых для науки видов рыб было получено именно из этого источника. Сами алепизавры служат пищей тунцам, некоторым пелагическим акулам и своим более крупным собратьям.
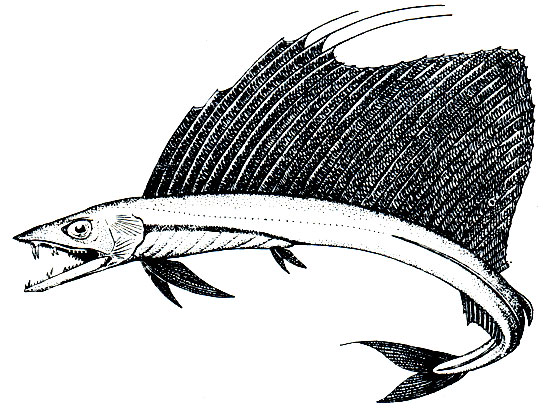
Рис. 102. Алепизавр (Alepisaurus ferox)
О размножении алепизавров известно немногое. По-видимому, они гермафродиты, и в теле каждой рыбы развиваются и женские и мужские половые продукты.
Алепизавры постоянно попадаются в качестве прилова при ярусном лове тунцов. Вряд ли часто употребляются они в пищу, но мясо их, несмотря на некоторую дряблость, отличается превосходным вкусом.
Семейство Омосудовые (Omosudidae)
Это семейство содержит всего один род - Омосудис (Omosudis) с единственным видом (О. lowei), который обитает преимущественно в тропических водах Атлантического, Индийского и Тихого океанов. Омосудис - настоящий океанический вид, редко встречающийся над континентальным шельфом. Он населяет толщу воды до глубины примерно 2000 м и совершает в течение суток вертикальные миграции, поднимаясь ночью на относительно меньшие глубины. Кроме того, для омосудиса характерно изменение глубины обитания с возрастом: чем он старше и крупнее, тем глубже живет.
Личинки этого вида ночью встречаются у поверхности моря, взрослые же рыбы, по-видимому, никогда к ней не поднимаются.
Омосудис имеет очень характерный и запоминающийся облик, и его трудно спутать с другими близкими к нему рыбами. Тело его сравнительно короткое и высокое, очень сильно сжато с боков и лишено чешуи. Голова также сильно сжата, очень большая.
Рот огромный, с необыкновенно массивной нижней челюстью. Громадные ланцетовидные зубы располагаются на челюстях и нёбных костях.
Глаза омосудиса имеют нормальное строение и лежат по бокам головы. Спинной плавник маленький, находится немного позади середины тела, над брюшными. Жировой плавник есть. Анальный плавник отодвинут далеко назад. С каждой стороны на хвостовом стебле имеется довольно высокий и длинный хвостовой киль.
Плавательного пузыря и светящихся органов у омосудиса нет.
У живых взрослых омосудисов тело имеет очень яркую иридирующую медно-серебристую окраску, которая к спине постепенно переходит в черную или темно-коричневую; сквозь радужную кожу просвечивает агатово-черная брюшина. Над кожистым килем на хвостовом стебле проходит интенсивно-черная полоса; также черные конец рыла, конец нижней челюсти и края обеих челюстей. Вертикальные плавники темные или черные, в грудных слегка окрашены только верхние лучи, брюшные не пигментированы.

Рис. 103. Омосудис (Omosudis lowei)
Некоторые исследователи полагают, что сверкающая радужная окраска омосудиса служит, поскольку у него нет светящихся органов, двум целям. Во-первых, блестящая поверхность тела, отражая свет, излучаемый светящимися организмами, улучшает условия зрения и тем самым облегчает добычу пищи. Во-вторых, постоянно меняющийся при движении рыбы отраженный свет может сбить с толку хищника и сохранить омосудису жизнь.
По характеру питания омосудис - хищник, причем в раннем возрасте он поедает главным образом молодь кальмаров, а становясь старше, постепенно переходит на рыбу. Чаще всего в желудках омосудисов встречаются рыбы-топорики (Sternoptychidae), на втором месте стоят светящиеся анчоусы (Myctophidae). Нередко размер добычи превышает размер омосудиса. Это объясняется тем, что он имеет мощные зубы, очень большой растягивающийся желудок и довольно длинный и сильный хвост, позволяющий ему быстро двигаться и настигать крупную добычу. Печень и сердце омосудиса помещаются в самой передней части тела, кости плечевого пояса - в наружном слое мышц, половые железы очень узкие и длинные - все это хорошо приспособлено к заглатыванию и перевариванию крупной добычи. Интересно, что переваривание пищи у омосудиса (так же как и у его ближайшего родственника - алепизавра) происходит только в кишечнике и проглоченные им рыбы или кальмары хорошо сохраняются в желудке до поступления в кишечник. С чем связана эта особенность, не ясно, и сейчас лишь остается предполагать, что желудок омосудиса изменил свою основную функцию и служит только своего рода складом. Основными врагами омосудиса, насколько известно, являются алепизавры и тунцы; был он также обнаружен и в желудке хиазмода (семейство Chiasmodontidae).
Семейство Эверманелловые (Evermanellidae)
Семейство эверманелловых наиболее близко к омосудовым (Omosudidae), жемчужноглазым (Scopelarchidae) и веретенниковым (Paralepididae). С первым из них его объединяют многие черты строения скелета и боковой линии, а также редуцированный язык, со вторым - общий внешний вид и телескопические глаза, с третьим - зубное вооружение. От этих и других близких форм эверманелловые хорошо отличаются тем, что имеют очень короткий язык и необыкновенно длинные зубы на нёбе.
Семейство содержит 3 рода и 6 видов. Для этих рыб характерны довольно высокое, сжатое с боков и лишенное чешуи тело, большая голова с тупым рылом, довольно массивная нижняя челюсть, усаженная длинными саблевидными зубами. У родов Коккорелла (Coccorella) и Эверманелла (Evermanella) глаза имеют телескопическое строение, а у одонтостомов (род Odontostomus) глаза обычного типа. Кости скелета, как и у многих близких групп, тонкие и слабые. Плавательного пузыря нет. Светящиеся органы у эверманелловых рыб, по-видимому, также отсутствуют. Светлое блестящее пятно, лежащее под глазом, скорее всего, служит лишь рефлектором, отражающим свет других организмов. Окраска живых рыб очень красива. У молодых черных эверманелл (Evermanella atrata) тело агатово-черное; щеки, жаберные крышки, челюсти, брюхо переливаются золотым, синим и зеленым; спина перед спинным плавником сверкает изумрудной зеленью. Окраска взрослых серебристых эверманелл (Е. balbo) другого вида - серебристая, с бледно-розовым оттенком; все тело этой рыбы покрыто черными точками и звездчатыми пятнышками.
Биология эверманелловых рыб изучена еще очень мало. По характеру питания они хищники, и в их желудках неоднократно находили полупереваренные остатки мелких рыб, и в частности светящихся анчоусов (Myctophidae).
Распространены эверманелловые рыбы довольно широко и известны из Средиземного моря и тропических и субтропических областей всех трех океанов. Вертикальные пределы их обитания слабо изучены. Наибольшее количество экземпляров было поймано при ловах от глубины 700-1000 м до поверхности, а в ловах менее чем от 200-метровой глубины попадались только мелкие мальки. По-видимому, взрослые рыбы не поднимаются выше 200 м. Что касается нижней границы распространения эверманелловых рыб, то, судя по их строению и окраске, можно полагать, что она проходит на глубине окола 2000 м.

Рис. 104. Одонтостом (Odontostomus atratus)
Семейство Жемчужноглазые (Scopelarchidae)
Семейство жемчужноглазых рыб сравнительно невелико и содержит 5 родов (Scopelarchus, Scopelarchoides, Phanops, Benthalbella, Neoscopelarchoides) с примерно 20 видами. Эти рыбы наиболее близки к семейству эверманелловых рыб (Evermanellidae), с которыми их сближает общий внешний облик и телескопическое строение глаз, у основания которых имеется своеобразное жемчужного оттенка пятно, вероятно светящееся у живых рыб. Характерно также не слишком удлиненное и относительно высокое, покрытое чешуей тело, большой рот с мощным зубным вооружением, довольно длинный язык, также несущий крупные зубы, отсутствие светящихся органов на теле и беловато-серый тон окраски с легким медным отблеском.
Голова довольно большая; глаза большие, телескопические и направлены у разных видов вперед, или вверх, или даже вверх и немного назад. Рот большой, достигает заднего края глаза. Нижняя челюсть, на которой находятся самые крупные зубы, не такая массивная, как у омосудиса или у эверманелловых рыб. Все плавники, кроме анального, небольшие; длина анального плавника превосходит длину головы. Над его задней частью, на спинной стороне тела, всегда есть хорошо развитый жировой плавничок. Плавательного пузыря у жемчужноглазых рыб нет.

Рис. 105. Жемчужноглаз (Scopelarchus candelops)
Наиболее крупный из известных представителей семейства имел длину всего около 15 см. Есть, однако, основания полагать, что эти рыбы могут достигать по крайней мере вдвое большей длины. По- видимому, жемчужноглазые рыбы - сильные и быстрые пловцы. Так можно думать на том основании, что науке известно очень немного более или менее крупных экземпляров, пойманных исследовательскими орудиями лова или найденных в желудках более крупных хищников.
Сами жемчужноглазые - активные хищники и питаются, судя по содержимому их желудков, главным образом рыбой (светящимися анчоусами, семейство (Мусtophidae) и, в меньшей мере, креветками.
Распространены жемчужноглазые очень широко и встречаются во всех трех океанах, преимущественно в тропических и умеренно теплых водах; один из видов обитает в Антарктике. Отсутствуют эти рыбы в Средиземном и Красном морях, в заливах и прибрежных водах; очень редко встречаются они над прибрежным мелководьем - шельфом. Наибольшая глубина, с которой известен представитель семейства, составляет около 2500 м. В общем для жемчужноглазых рыб существует тенденция к увеличению глубины обитания с возрастом. На наименьших глубинах, близ поверхности, ловятся личинки и мелкие мальки, подросшие неполовозрелые рыбы известны с глубин 750-1650 м, а взрослые - с глубин 1650-2500 м.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'