Глава десятая
- Так и так одинаково, - сказал он. - Джордж, почему одинаково"? - Не знаю, - сказал Джордж. - Так их рисуют.
В седьмой главе, которая осталась так далеко в прошлом, я упомянул о том, что нам хотелось предоставить еще одной перенаселенной колонии больший простор и посмотреть, как это подействует на ее "общественное устройство" и на размножение. У нас как раз сложилась колония, удивительно подходившая для такого эксперимента. Вначале она росла быстрее остальных, главным образом потому, что первые самки обе остались в живых и сохраняли плодовитость. Кроме того, несколько их дочерей успели вырасти и принести потомство до того, как перенаселенность дала о себе знать.
Еще одной причиной быстрого роста колонии была низкая смертность мышат. Например, согласно переписи от 27 июля 1955 года, в этой выгородке находилось 143 мыши, включая 23 новорожденных. Хотя в выгородке имелось восемь укрытий для гнезд, одно из них служило тесным обиталищем для 61 мыши. Эта масса мышей включала 28 самцов (весом от 10,0 до 17,2 грамма), 29 самок (весом от 10,0 до 21,1 грамма) и четыре помета разного возраста - всего 14 мышат. Поскольку укрытие имело площадь только 30 на 30 сантиметров, а высоту 8 сантиметров, этим мышам там было негде повернуться в буквальном смысле слова. Однако, хотя кормящие самки после переписи немедленно переселились под другое укрытие, что было чревато опасностью для молодых мышат, при следующей переписи 11 из них были живы и сильно выросли.
Большинство самок, рожденных в этой выгородке, впадало в длительное состояние анэструса, что приводило к снижению прироста. Из общего числа учтенных молодых мышат (162) на долю первых двух самок приходилось 68- 82. Более точную цифру дать невозможно, так как при подобных условиях материнство иногда неустановимо. Прочие 80-94 рождения были прослежены только у 8 из подросших дочерей. Остальные же 19 самок первого поколения и все 45 самок второго поколения потомства не дали вовсе. Они не беременели - оставались в состоянии анэструса.
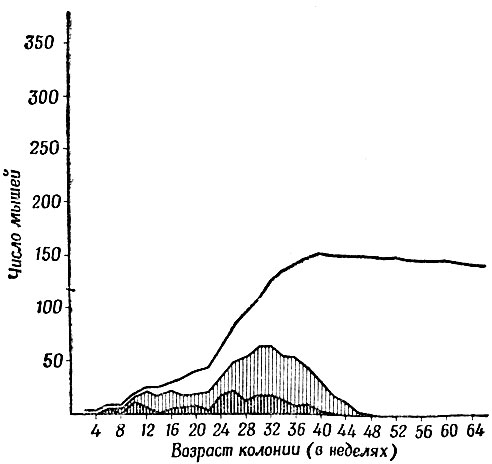
Рис. 21. Рост замкнутой колонии домовых мышей, получавших обильный корм. Нижняя кривая показывает число новорожденных, средняя новорожденных и молодых мышат, а верхняя - общее число всех мышей. Через 48 недель популяция по возрасту стала совершенно "взрослой", но большинство самок ни разу не дало потомства
По мере того как мышата вырастали, но не вносили своей доли в прирост популяции, возрастная структура колонии начала отражать ее старение. Популяция со здоровым ростом обладает широким фундаментом из новорожденных и молодых мышат, крепким первым этажом из взрослых особей, дающих потомство и более узким вторым этажом из стариков-пенсионеров. Проходили недели, и популяция стала почти статичной, поскольку все молодые особи достигли зрелого возраста, а рождения прекратились и смертей почти не было. Возможность того, что мышата рождались без нашего ведома и их съедали, полностью исключается, так как половые щели самок оставались плотно закрытыми. Только у одной-двух самок старшего поколения они открывались на короткие периоды.
Это постепенное изменение возрастной структуры колонии показано на рис. 21. Верхняя линия, соответствующая общей численности, образует довольно точную "сигмоидную" кривую, похожую на те, которые можно видеть во всех учебниках экологии. Другими словами, популяция стабилизировалась, когда полностью использовала какой-то ограниченный ресурс - в данном случае жизненное пространство. Недостатка в корме не было, так как даже наиболее подчиненные мыши могли наедаться досыта днем, пока остальные обитатели колонии оставались в гнездах. Слабое понижение кривой объясняется несколькими смертями, причина которых осталась невыясненной, но, вероятно, не была связана с перенаселенностью. А в остальном мыши были здоровы, и способность самцов к продолжению рода отнюдь не угасала.
Мы сохраняли эту чисто взрослую колонию в течение 26 недель и с большим удовольствием сохранили бы ее еще и дольше. Мы полагали, что размножение начнется снова, когда смерть от старости приведет к достаточному сокращению численности. Но нам необходимо было завершить весь эксперимент не позже чем через два месяца. Я уже перешел на административную работу в министерство и даже на этот срок мог сохранить колонию только благодаря любезности моего начальства. Мы с Фредом решили отвести напоследок душу и выпустить нашу колонию в более обширное помещение.
16 марта 1956 года мы осмотрели и взвесили мышей в выгородке, а вечером открыли подъемную дверцу в новую выгородку площадью около 36 квадратных метров (рис. 22). Перепись выявила наличие 140 мышей. Шестьдесят семь из них были самками, но половая щель была открыта только у одной - у еще не скончавшейся бабушки. На этот раз, как и тогда, когда мы расширили выгородку № 3, первые исследователи нового пространства уже через несколько минут принялись "засталбливать" участки и агрессивно бросаться на всех остальных. Покидая в эту ночь Ругэмский аэродром, мы знали, что, вернувшись на следующей неделе, обнаружим очень интересное положение вещей.

Рис. 22. Общий вид одной из экспериментальных колоний. В нижнем левом углу расположена маленькая подъемная дверца, соединяющая перенаселенную выгородку с большой центральной выгородкой. В верхней части снимка - стремянка, с которой велись наблюдения
Через восемь дней мы бесшумно опустили металлические рукава на каждое укрытие для гнезда и снова осмотрели мышей. Изолировав таким образом каждое гнездо, мы могли не только установить, в каком состоянии находятся самки, но и получить сведения об "общественной организации", сложившейся в новой выгородке. Там обитало 26 самцов и 29 самок. Ни в одном укрытии не жило больше одного самца. Многие самцы были "на холостяцком положении", как выразился Фред, но у семи из них было по одной или больше подруг. Половая щель у большинства этих самок (19 из 29) была открыта или вот-вот должна была закрыться.
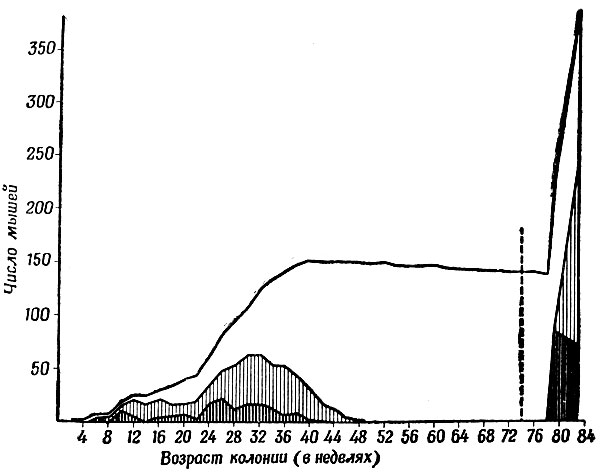
Рис. 23. Та же колония, что и на рис. 21, давшая взрыв роста численности мышей вслед за их расселением в выгородке большей площади. При отсутствии дальнейшего расселения количество мышей снова стабилизировалось бы, но на более высоком уровне
В старой выгородке распределение следовало прежней схеме. В одном укрытии жил одинокий самец, но остальные были заняты смешанными группами самок и самцов поплоше. Только у 15 из 38 самок половая щель была открыта или закрывалась, из чего следовало, что между колонизаторскими способностями самок тоже существуют различия. Чтобы завершить эксперимент, нам оставалось сделать еще два дела: во-первых, определить конкретные границы участков с помощью как прямого наблюдения, так и установления личности самцов, принимающих участие в пограничных спорах, и, во-вторых, убедиться, действительно ли изменившееся состояние самок приводит к размножению. Благодаря неизменной любезности доктора Юона Томаса мы смогли выполнить и ту и другую задачу.
Когда через 23 дня мы провели новую перепись, то обнаружили 82 молодых мышонка, а многие из некормящих самок были беременны. Еще через 35 дней мы устроили последнюю перепись, которая была и моей заключительной работой в мышином доме. В промежутках между переписями мы вели кое-какие наблюдения и достаточно точно установили границы индивидуальных участков. Последняя перепись показала появление 160 новых младенцев!
Таким образом, 140 мышей за два месяца после расселения успешно произвели на свет 242 отпрыска! Большинство мышат, помеченных во время первой переписи, были обнаружены и при второй, что указывало на крайне низкую смертность. Но скорость размножения уже уменьшалась. На рис. 23 показан взрыв, вызванный тем, что мы расширили жизненное пространство колонии. Я не сомневаюсь, что, будь у нас возможность сохранить эту колонию подольше, кривая снова выровнялась бы. А тогда, если бы мы открыли дверцу в еще более просторную выгородку... Но самым интересным было бы наблюдение за тем, как новая перенаселенность привела бы к преобразованию общества с отдельными участками в иерархию доминирования - подчинения.
Как мы видели и в других выгородках, площадь индивидуальных участков была очень различной (рис. 24). Некоторые самцы властвовали над относительно большими территориями и успешно не допускали на них никаких других мышей, кроме своих подруг (участки А, Д и Ж). Но, как и в людских браках, ответить на вопрос "Сам он выбрал или его выбрали?" было бы нелегко. Другие самцы защищали одно только укрытие для гнезда внутри узкого кольца территории (участки Г, Л и Н). Две мыши обзавелись участками, не включавшими укрытия. Первым был самец, который защищал открытое гнездо, пристроенное к одной из стен (участок Г), но не сумел подыскать себе подругу. А второй была беременная самка, которая устроила гнездо в одной из кормушек (участок М) и там успешно вырастила своих мышат.
Заштрихованные участки оставались во владении одного и того же хозяина все два месяца. Незаштрихованные, возникнув примерно в то же время, позже были брошены, и хозяева их по большей части вернулись в старую выгородку. Эти мыши, так сказать, были иммигрантами, у которых не хватило упорства и решимости преодолеть первоначальные трудности освоения нового края и они предпочли вернуться в свои родные, перенаселенные, но обжитые места.
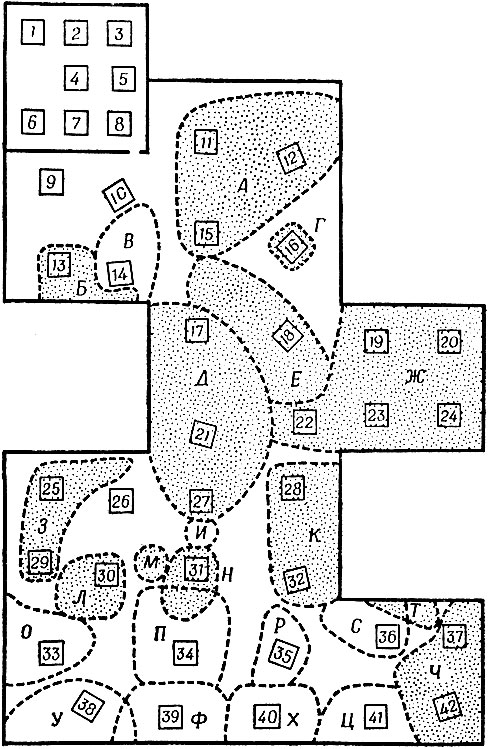
Рис. 24 Примерные границы индивидуальных участков (А - Ч) в выгородке № 8 через восемь дней после начала расселения. Заштрихованные площади защищались одними и теми же самцами все последующие два месяца. Укрытия для гнезд № 1-8 находятся внутри исходной выродки
Некоторые мыши переменили участки. Несколько участков было захвачено теми, кто явился в эту выгородку позднее. Но девять из первоначальных колонистов сохранили сразу выбранные ими участки, показав, что основа "общественного порядка" была заложена на первых же стадиях переселения. Хотя нам не удалось собрать достаточно неопровержимые доказательства этого факта, я тем не менее твердо убежден, что "раздел" новой территории был завершен еще до истечения вторых суток после начала переселения. Если вы намерены стать в Новом Свете главой старинного рода, отправляйтесь туда на первом же корабле.
Самцы, которые владели участками до самого конца, были значительно тяжелее и старше тех, которые остались в старой выгородке. Как мы обнаружили еще в выгородке № 3, они вовсе не обязательно были теми, кто первым попытался захватить участок. Нет, это были мыши, достаточно крупные и сильные, чтобы свести на нет то небольшое преимущество, которое приобретали мыши, первыми вступившие на новую территорию. Все самцы - владельцы участков, кроме одного, имели по одной или по нескольку подруг. При первой переписи с одним самцом жило семь самок, но при второй переписи он оказался в старой выгородке. Единственный самец, сохранивший трех самок (участок Ж), был, кроме того, и владельцем самого большого участка - как и единственный многоженец в круглой выгородке, описанной в главе пятой.
Изучение биографий иммигранток дало не менее интересные результаты. Из 29 самок, живших на новой территории при первой переписи, 27 там и остались, и к ним присоединилось еще 11 самок. При первой переписи эти самки были весьма неравномерно распределены между самцами, но затем они в большинстве оказались партнершами в моногамных союзах, и это положение сохранялось, пока они растили мышат. Преобладание моногамии объяснялось не столько склонностями самцов, сколько агрессивностью их первых подруг. Эти 38 колонисток были также значительно старше и тяжелее тех самок, которые оставались в старой выгородке. Такое возрастное разделение было даже еще более четким, чем у самцов, и протоколы показали, что в новой выгородке поселились только уже рожавшие самки. Это также дает пищу для не относящихся к делу размышлений, но мы не будем им предаваться.
В те годы, когда Фред Роу и я работали с мышами, а также и впоследствии многие экологи и физиологи занимались мышиными популяциями, и хотя я не намерен давать тут общий обзор, который не может не быть весьма сухим, все-таки стоит посмотреть, как наша работа укладывалась в общие рамки. Нас не слишком интересовали возможные теоретические следствия наших открытий. Из экспериментов с популяциями мы не извлекли никаких сведений, которые могли бы оказаться полезными в борьбе с домовой мышью как с вредителем, и узнали только, что популяционные уровни, на которых вступает в действие механизм саморегулирования, слишком высоки, чтобы рассчитывать на этот механизм в общей программе очищения штабелей и скирд от мышей. Эти эксперименты были полезны для установления, какими при определенных условиях могут быть распределение мышей и их "общественное устройство".
Эксперименты Стрекера и Эмлена с замкнутыми колониями, получавшими ограниченный корм, уже упоминались. В этих колониях, как и в наших, падение плодовитости играло в ограничении численности гораздо большую роль, чем смертность. Одна из их популяций стала чисто взрослой, как и наша популяция в выгородке № 8. Как ни странно, мыши обеих групп - и те, кто получал мало корма, и те, кто получал его в изобилии, - имели в теле жировые запасы. Без сомнения, этот жир должен был бы помочь им начать жить заново, если бы им удалось найти избавление от неблагоприятной среды.
Саутвик (1955) опубликовал результаты дальнейших экспериментов в Висконсине как раз тогда, когда наша работа подошла к концу. Чак задумал чрезвычайно изящный эксперимент с помещением мышей в две экологически резко отличные среды. К несчастью, то, что представляется резко отличным человеческому взгляду, оказалось совершенно одинаковым с точки зрения мышей, но это не помешало ему опубликовать несколько работ о мышах, которые являются наиболее полными из всех, появившихся до сих пор. И Саутвик и Кристиен (1956) давали корм без ограничений, но оба они, как и Браун, обнаружили, что основным фактором, ограничивающим численность популяции, была высокая смертность среди новорожденных и молодых мышат. Наблюдалось и снижение плодовитости, особенно среди молодых самок, но этого оказалось недостаточно, чтобы уравновесить рост популяции.
Таким образом, некоторые исследователи обнаружили, что замкнутые колонии перестают расти из-за большого сокращения рождаемости и небольшого увеличения смертности, тогда как другие наблюдали небольшое сокращение рождаемости и значительное увеличение смертности. Мне кажется странным, что наблюдения над одним и тем же животным дали столь принципиально различающиеся результаты, но Кристиен (1959) считает и те и другие результаты в принципе одинаковыми, так как, по его мнению, прекращение рождаемости и смерть новорожденных представляют собой два вида действия одного и того же механизма - "зависящей от плотности социо-психологической обратно-действующей постоянно оперирующей системы, которая функционирует в качестве основного, а часто и единственного регулятора роста популяции". Кристиен, по-видимому, полагал, что, изучая колонии диких мышей в клетках (оптимистически именуемые "свободно растущими" колониями) и сравнивая некоторые их особенности с особенностями изолированных мышиных пар, он исследует единственную переменную величину - плотность. На самом же деле он, помимо многого другого, изучал конструкцию ящичка-гнезда, вмешательство наблюдателя и "убежища" - изучал на материале мышиного концентрационного лагеря, комендант которого не счел нужным вытатуировать номера на заключенных.
В одной из статей мы с Фредом писали:
"Если подобные "механизмы управления численностью популяций" настолько зависят от специфически экспериментальных условий, мы по-прежнему не можем не ставить под вопрос их значение для популяций, живущих на свободе. Ни у одной возникшей естественно мышиной популяции не было обнаружено снижения плодовитости как реакции на большую численность. Даже в английских скирдах, которые представляют собой среду обитания, наиболее благоприятную для быстрого роста численности, не было обнаружено плотности популяций, близкой к наблюдавшимся в наших колониях. Таким образом, данный саморегулирующий механизм, возможно, начинает действовать при плотностях, которых живущие на свободе мыши достигают редко (если вообще когда-нибудь достигают), поскольку их численность сокращается по другим причинам". Позже Фред обнаружил частичное подавление размножения в скирдах хлебных злаков. Что касается реального механизма подавления (Роу и др., 1964), то скирды двух разных типов как будто соответствуют двум разным типам условий в выгородках. В овсяных скирдах, где снопы, сильно засоренные сорняками, складываются свободно, наблюдались значительно более высокие плотности мышиных популяций, гнезда были лучше, имелось больше молодых мышат и способных к размножению самок. В пшеничных скирдах с их тесной укладкой плотности были ниже, гнезда хуже, мышат меньше, а смертность новорожденных выше. Реальное жизненное пространство в пшеничных скирдах гораздо меньше, и ходы были обнаружены только между снопами.
Следует помнить, что хлебная скирда - это, в сущности, такое сооружение, с помощью которого фермер обеспечивает домовым мышам возможность благополучно перезимовать в количествах, достаточных для того, чтобы на следующую весну как следует заразить поля. Когда речь идет о естественно выработавшемся механизме контроля над численностью популяций, популяцию, обитающую в скирде, можно назвать "естественной" только с очень большой натяжкой. Практика хранения зерна, бесспорно, создает такие защищенные от непогоды места, где корм обилен, а хищников нет и лишь перенаселенность вызывает падение плодовитости. Однако пока никто еще не показал, что подобные физиологические моменты воздействуют на сокращение численности каких-либо видов животных в "полевых условиях".
Работая над этой главой, я увидел в местной газете сообщение о нашествии мышей, поразившем округ Саут-Хаммок вблизи Порт-Уэйкфилда, на юге Австралии. Так как я ни разу не видел подобных нашествий и скептически относился к сообщениям о младенцах, изгрызенных, в колыбелях, и об объевшихся котах, которые не желают больше обращать на мышей никакого внимания, я поехал туда, чтобы своими глазами увидеть, как обстоят дела. Да, это было настоящее нашествие. Межи и поля были пронизаны норами и ходами, туда отовсюду слетелись орлы, ястребы и соколы, а по ночам, если мы выходили без фонарика, мыши шмыгали прямо под ногами.
У нас там стоял автоприцеп, и вот, вооружившись священными текстами по Экологии Животных, я удалился в оный автоприцеп, дабы написать заключение к этой главе. Здесь, где никто не будет мне мешать, никто не будет отрывать меня, я сделаю обзор теорий регуляции численности популяций и закончу эту легкомысленную книжку на глубоко интеллектуальной ноте.
Но мыши уже успели забраться в автоприцеп. Я пишу, а они бегают по столу и дерутся на полу возле моих ног - эти сытые полевые мыши, которым очень понравились мои кукурузные хлопья. Прошлой ночью мне не давал уснуть шум, который они поднимали, и их вторжения в мой спальный мешок. А теперь меня томит из-за них тоска по мышиному дому, по хлебным полям у заброшенного аэродрома, по неуемному энтузиазму давно ушедших лет. И пожалуй, лучше кончить на этом - просто закрыть пишущую машинку и выбросить все остальное. Как сказал Ленин, когда он бросил мертвого щенка: "И без того плохо..."
Литература
Christian J. J. (1956), Adrenal and reproductive responses to population size in mice from freely growing populations, Ecology, 37, 258-273.
Christian J. J. (1959), The roles of endocrine and behavioural factors in the growth of mammalian populations, Symp. on Comparative Endocrinology, 71-97, John Wiley and Sons, N. Y.
Crowcroft P., Rowe F. P. (1957), The growth of confined colonies of the wild house mouse (Mus musculus L.), Proc. zool. Soc. Load., 129, 359-370.
Crowcroft P., Rowe F. P. (1958), The growth of confined colonies of the wild house mouse: the effect of dispersal on female fecundity, Proc. tool. Soc. Lond., 131, 357-365.
Negus N. C, Gould E., Сhipman R. K. (1961), Ecology of the rice rat, Oryzomys palustris (Harlan), on Breton Island, Gulf of Mexico, with a critique of the social stress theory, Tulane Stud. Zool., 8(4), 93-123.
Rоwe F. P., Taylor E. J., Сhud1eу А. Н. J. (1964), The effect of crowding on the reproduction of the house mouse (Mus musculus L.), living in corn-ricks, J. Anim. Ecol., 33, 477-483.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'