Класс споровики (Sporozoa)
Класс споровиков представляет собой обширную группу простейших организмов, ведущих исключительно паразитический образ жизни. Приспособление к паразитизму у них очень глубокое и совершенное. У всех споровиков отсутствуют специальные органоиды захвата пищи, и они питаются осмотическим путем. Хозяевами их являются самые различные беспозвоночные и позвоночные животные. Несколько видов споровиков паразитирует в человеке.
В процессе эволюции они приспособились к паразитированию в самых различных органах и тканях. Многие споровики - паразиты кишечника и различных органов, связанных с пищеварительной системой (в том числе печени). Имеются виды, паразитирующие в органах выделительной системы - почках. Органы кровеносной системы и кровь также служат средой обитания некоторых споровиков.
В этой группе простейших есть немало видов, приспособившихся к внутриклеточному паразитизму. Паразиты проникают внутрь клеток различных тканей хозяина, питаются, растут и развиваются за их счет.
Одной из форм глубокого приспособления споровиков к паразитизму явилась выработка сложных и разнообразных жизненных циклов, обеспечивающих заражение хозяина. По ходу циклов происходит смена разных форм размножения, ведущих, с одной стороны, к увеличению числа паразитов в данной особи хозяина, а с другой - к образованию таких стадий, которые служат для заражения новых особей хозяина.
У некоторых споровиков выработалось не только чередование различных форм размножения, но и смена хозяев, относящихся к разным видам и группам животного мира.
Многие виды споровиков приносят большой вред как возбудители заболеваний человека, домашних и промысловых животных.
В данном разделе мы ограничимся рассмотрением лишь немногих споровиков, представляющих наибольший биологический интерес или особенно большое практическое значение.
Отряд грегарины (Gregarinida)
Грегарины - большая группа споровиков, включающая около 500 видов. Все без исключения грегарины - паразиты беспозвоночных. Наибольшее число видов их обитает в кишечнике членистоногих, в особенности часто у насекомых. Некоторые грегарины паразитируют и в других группах беспозвоночных - в кольчатых червях, иглокожих, оболочниках. Кроме кишечных грегарин, каковых большинство, имеются виды, живущие в полости тела, в органах размножения и др.
Наиболее сложно устроены грегарины, живущие в кишечнике членистоногих. На их примере мы и ознакомимся с организацией представителей этого отряда.
На рисунке 67 показано строение взрослой формы Corycella armata - грегарины из кишечника личинки жука-вертячки Gyrinus natator. Тело ее, достигающее довольно значительной длины, разделено на 3 отдела. В наиболее крупном заднем отделе помещается ядро. Этот отдел называется дейтомеритом. Впереди от него расположен протомерит. Он отделен от деитомерита тонкой прослойкой светлой цитоплазмы, представляющей собой продолжение слоя эктоплазмы. На самом переднем конце грегарины расположен эпимерит. Этот участок тела служит для прикрепления грегарины к стенке кишечника. Часто он бывает снабжен крючьями, иногда нитевидными выростами и может принимать весьма причудливую форму. На рисунке 68 изображены некоторые характерные формы эпимеритов разных видов грегарин. По достижении предельных размеров, когда у грегарины наступают процессы полового размножения, эпимерит обычно отбрасывается. Он целиком состоит из эктоплазмы.
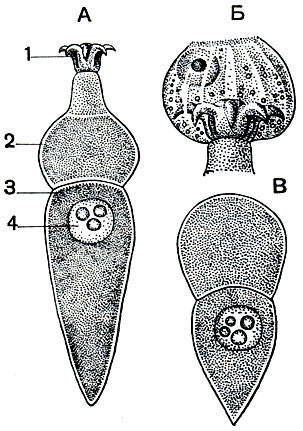
Рис. 67. Грегарина Corycella armata из кишечника насекомых: А - взрослая грегарина; Б - эпимерит грегарины, внедрившийся в эпителиальную клетку кишечника; В - грегарина, сбросившая эпимерит. 1 - эпимерит; 2 - протомерит; 3 - дейтомерит; 4 - ядро
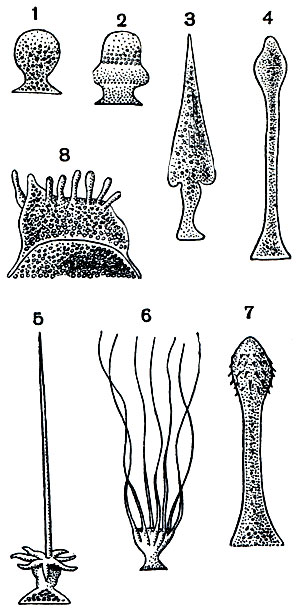
Рис. 68. Эпимериты различных видов грегарин: 1 - Gregarina longa; 2 - Sycia ini-pitata; 3 - Pileocephalus heeri; 4 - Stylocephalus longicollis; 5 - Beloides firnus; 6 - Cometoides crinitus; 7 - Geniorhynchus monnieri; 8 - Echinomera hispida
Разделение цитоплазмы грегарин на два слоя - эктоплазму и эндоплазму - выражено всегда очень отчетливо. Наружный слой эктоплазмы образует прочную пелликулу, определяющую постоянство формы тела грегарин. У некоторых грегарин в эктоплазме найдены тонкие скелетно-опорные волоконца, еще более увеличивающие прочность наружных слоев тела. Немногие виды грегарин способны несколько менять форму своего тела, сокращаться вдоль продольной оси. У таких видов в эктоплазме, на границе с эндоплазмой, найдены сократимые волоконца - миофибриллы.
Эндоплазма грегарины, резко отграниченная от эктоплазмы, при рассматривании под микроскопом живых грегарин выглядит темно-серой. Это обусловлено наличием в эндоплазме большого количества гликогена, нередко сконцентрированного в виде мелких гранул. В этом проявляется одна из характерных особенностей многих внутренностных паразитов, тело которых богато гликогеном. Кишечные паразиты живут в среде, в которой свободный кислород присутствует в ничтожных количествах. Это затрудняет или делает невозможными дыхательные процессы, сопровождающиеся поглощением кислорода. В этих условиях паразитические организмы вынуждены искать взамен дыхания другие источники энергии. Такими источниками оказываются процессы анаэробного (без участия свободного кислорода) расщепления гликогена, процессы гликолиза. Не вдаваясь в химическую характеристику этих процессов, отметим лишь, что при этом сложная молекула гликогена распадается на более простые органические соединения (в частности, некоторые органические кислоты). Этот процесс сопровождается выделением энергии. Однако в отношении количества освобождаемой энергии на единицу веса (на грамм-молекулу) гликогена при окислении выделяется во много раз больше энергии, чем при гликолизе. Окисление - гораздо более "экономичный" процесс, чем гликолиз. Поэтому обычно у организмов, основным источником энергии которых являются анаэробные гликолитические процессы, в теле имеются значительные скопления гликогена. Грегарины - один из ярких примеров, иллюстрирующих это правило.
Ядра, расположенные всегда в дейтомерите, имеют обычно вид пузырька, богатого ядерным соком и снабженного одним или несколькими крупными ядрышками.
Рассмотренное выше строение грегарины свойственно преимущественно кишечным паразитам. Оно характерно для группы видов, объединяемых в семейство Polycystidae. У представителей другого семейства - Monocystidae, которые паразитируют главным образом в полости тела, половых железах и других органах, строение более простое. У Monocystidae нет разделения на эпи-, прото- и дейтомерит, и тело их часто имеет червеобразную форму.
Наблюдая под микроскопом за живыми грегаринами, легко обнаружить, что они способны двигаться, медленно перемещаясь в содержимом кишечника. Это движение носит характер плавного скольжения, не сопровождающегося изменением формы тела.
Механизм этого процесса до настоящего времени не вполне ясен, поскольку у грегарин нет никаких специальных органоидов движения. Исследования Шевякова и других авторов показали, что на заднем конце движущейся грегарины выделяется слизь. По-видимому, она выбрасывается наружу с известной силой, благодаря чему и происходит - по принципу ракетного двигателя - поступательное движение грегарины.
У настоящих грегарин* отсутствует бесполое размножение, они размножаются только половым путем. Рассмотрим типичный цикл развития грегарин (рис. 69).
* (Кроме настоящих грегарин (отряд Eugregarinida), имеется небольшой отряд шизогрегарин (Schizogregarinida), размножающихся бесполым путем.)
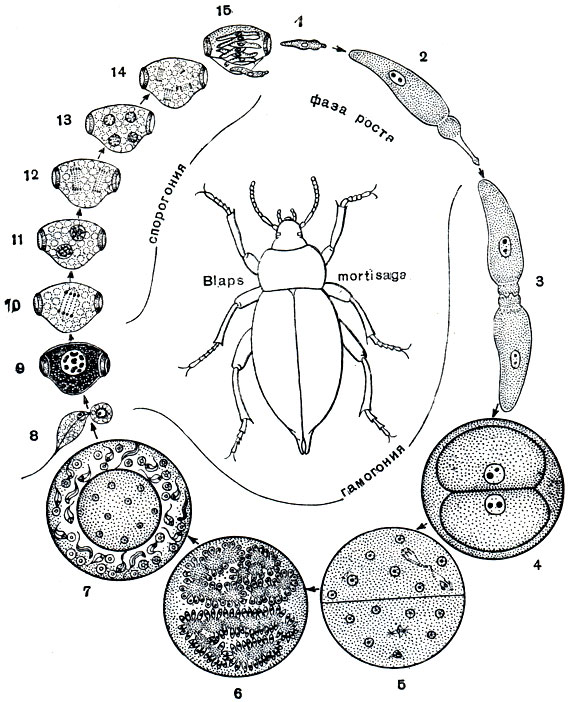
Рис. 69. Цикл развития грегарины Stylocephalus longicollis: 1-4 - в кишечнике жука В laps mortisaga; l - спорозоит; 2 - гамонт; 3 - соединение двух особей, образование сизигия; 4 - циста с двумя округлившимися гамонтами; 5-15 - во внешней среде; 5, 6 - деление ядер гамонтов, ведущее к образованию половых клеток (гамет); 7 - сформированные гаметы; 8 - копуляция гамет; 9 - зигота, образовавшая оболочку (ооциста); 10-14 - развитие спорозоитов в ооцисте (спорогония); 15 - спорозоиты, выходящие из споры
Живущие в кишечнике насекомого грегарины, достигнув предельного роста, теряют эпимерити соединяются попарно. Вокруг такой парочки (сизигия) выделяется общая оболочка (циста). Вскоре после инцистирования в каждой из соединившихся в сизигий особи начинается многократное последовательное деление ядра. В каждой особи образуется множество ядер. К концу этого процесса все ядра располагаются по периферии грегарины и вокруг каждого из них обособляется небольшой участок цитоплазмы. Так формируется множество мелких одноядерных клеток. Большая часть цитоплазмы при этом остается неиспользованной. Это так называемое остаточное тело, которое в дальнейшем процессе размножения участия не принимает. Мелкие одноядерные клетки представляют собой половые клетки - гаметы. Эти клетки сближаются попарно и сливаются.
У некоторых видов грегарин гаметы, образуемые обоими соединившимися в сизигий особями, по размерам и строению неотличимы друг от друга. У других видов они различны. Одна особь дает начало округлым и неподвижным гаметам. Это женские гаметы. Другая особь образует удлиненные гаметы, снабженные жгутиковидным выростом. Это мужские подвижные гаметы.
Образовавшиеся в результате оплодотворения зиготы сразу же покрываются оболочкой, принимая при этом веретенообразную, а нередко бочонковидную форму. Покрывшаяся оболочкой зигота носит название ооцисты. Оболочка цисты в этот период цикла развития еще не разрушается. Таким образом, многочисленные образовавшиеся после копуляции гамет ооцисты лежат внутри оболочки цисты. На этой стадии, а иногда и несколько позже, цисты с ооцистами выводятся из кишечника наружу. Последним этапом цикла развития грегарины является процесс формирования ооцисты, получивший название спорогонии. Он сводится к следующему. Внутри ооцисты ядро (образовавшееся в результате слияния ядер гамет) трижды делится. Вокруг каждого из возникших таким путем ядер обособляется участок цитоплазмы. В каждой ооцисте получается восемь очень маленьких червеобразной формы клеток, называемых спорозоитами. На этой стадии ооцисты способны к заражению новых особей хозяина. Если циста с ооцистами или даже отдельная ооциста будет проглочена насекомым - хозяином данного вида грегарин, то под воздействием кишечного сока спорозоиты выходят из ооцисты. Они подвижны. В дальнейшем из каждого спорозоита развивается грегарина. Достигнув типичного для данного вида размера, грегарины вновь соединяются попарно, инцистируются, и цикл начинается снова.
Таким образом, рассмотренный выше цикл развития грегарин можно отчетливо разделить на три этапа. Первый - это рост и развитие взрослой грегарины. Второй - образование половых клеток (гамет) и оплодотворение. Третьим этапом цикла является спорогония - развитие зиготы (ооцисты) и формирование спорозоитов.
Отряд кокцидии (Coccidia)
Так же как и грегарины, кокцидии представляют собой обширную группу паразитических простейших, включающую около 400 видов. Паразитируют они в самых различных группах беспозвоночных и позвоночных животных: кольчатых червях, моллюсках, членистоногих, позвоночных. Для кокцидии характерен внутриклеточный паразитизм в различных органах и тканях. Их жизненный цикл слагается (за единичными исключениями) из закономерного чередования бесполого размножения, полового процесса и спорогонии. У одних видов спорогония совершается вне организма хозяина, в наружной среде; у других, при наличии смены хозяев, бесполое размножение и спорогония происходят в организме разных хозяев.
Некоторые виды кокцидии являются опасными паразитами домашних животных, домашней птицы и некоторых разводимых в рыбоводческих хозяйствах рыб. Один очень редкий вид кокцидии паразитирует в человеке. Рассмотрим один пример из числа наиболее обычных и вместе с тем патогенных видов.
В кроликах паразитирует несколько видов рода эймерия (Eimeria). Большая часть их - паразиты кишечника, один вид локализуется в печени. Эймерии - кокцидии, развитие которых не сопровождается сменой хозяев. В их жизненном цикле имеется два основных этапа. Первый этап цикла протекает в кишечнике кролика. Это эндогенная часть цикла.
Второй этап проходит вне тела хозяина, во внешней среде - экзогенная часть цикла.
Eimeria magna - один из наиболее широко распространенных видов кокцидии кролика. На рисунке 70 схематически представлены основные этапы жизненного цикла этого паразита.
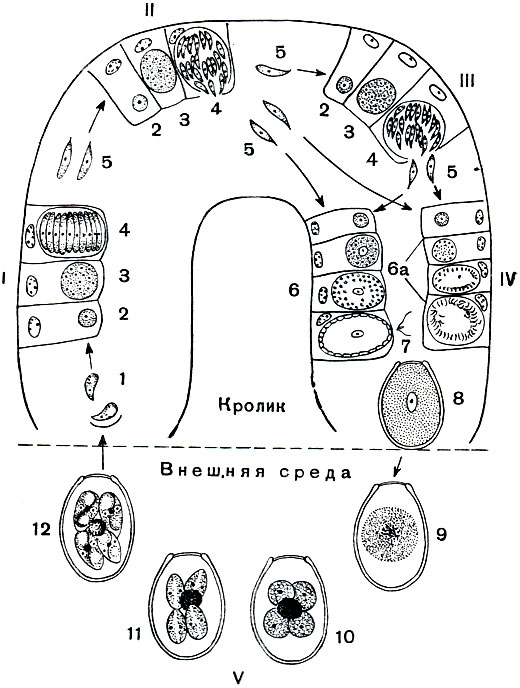
Рис. 70. Цикл развития кокцидий Eimeria magna: I-первое поколение шизонтов; II - 2-е поколение шизонтов; III - 3-е поколение шизонтов; IV - гаметого-ния; V - спорогония. 1 - спорозоиты; 2 - молодой шизонт; 3 - растущий шизонт с многими ядрами; 4 - шизонт, распавшийся на мерозоиты; 5 - меро-зоиты; 6 - развитие макрогаметы; 6а - развитие микрогамет; 7 - микрогамета; 8 - ооциста; 9 - ооциста, вышедшая из кишечника кролика; 10 - ооциста с четырьмя споробластами и остаточным телом; 11 - развитие спор; 12 - ооциста с 4 зрелыми спорами (в каждой споре по 2 спорозоита)
Начнем рассмотрение жизненного цикла Eimeria magna с момента, когда паразит проникает в кишечник хозяина. Первой стадией эндогенной части цикла является спорозоит, выходящий из заглоченной кроликом ооцисты (строение и развитие ооцист будет рассмотрено ниже). Спорозоиты - это очень маленькие клетки (длиной около 8 мк, шириной 2-3 мк), веретеновидной формы с одним ядром. Они активно внедряются в клетки эпителия кишечника, где тотчас же округляются, становятся шаровидными. За внедрением кокцидии в клетку хозяина следует активный рост: паразит увеличивается в размерах. Питание его осуществляется осмотическим путем. На этой стадии развития паразит носит название шизонта в соответствии с тем способом размножения, к которому он приступает.
По мере роста паразита начинается деление ядра, благодаря чему шизонт становится многоядерным. Число ядер варьирует в довольно широких пределах - от 8 до 60. Растущий шизонт разрушает эпителиальную клетку и постепенно перемещается в субэпителиальный слой соединительной ткани. Развитие шизонта завершается бесполым размножением. Вокруг каждого из ядер обособляется участок цитоплазмы, в результате чего весь шизонт распадается на одноядерные мелкие клетки, каждая из которых имеет веретеновидную форму (рис. 71). На этом заканчивается процесс бесполого размножения шизонта. Такое размножение носит название множественного деления или шизогонии. Весь процесс от момента внедрения спорозоита в эпителиальную клетку до завершения шизогонии занимает 80-96 часов. Одноядерные веретеновидные клетки, образовавшиеся в результате шизогонии, называют мерозоитами. Какова их дальнейшая судьба?
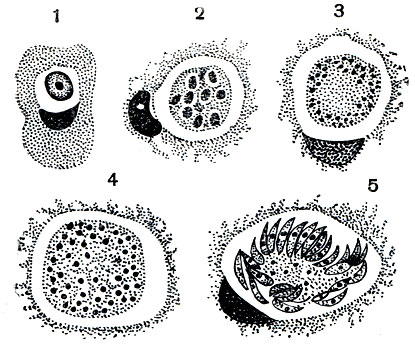
Рис. 71. Рост шизонта и шизогония кокцидий Eimeria magna: 1 - молодой шизонт, ядро еще не приступило к делению; 2-4 - увеличение числа ядер путем деления и рост шизонта; 5 - распад шизонта на мерозоиты
Мерозоиты вновь внедряются в эпителиальные клетки кишечника и дают начало второму поколению шизонтов. Процесс развития шизонтов второго поколения занимает несколько больше времени, чем первого, - 110-132 часа.
Часть мерозоитов от второго поколения дает начало третьему поколению шизонтов. Образующиеся в результате его мерозоиты, так же как и часть мерозоитов второго поколения, не образуют шизонтов. Внедряясь в эпителиальные клетки, они дают начало гаметам (половым клеткам). Гаметы кокцидий резко дифференцированы в половом отношении на мужские микрогаметы и женские макрогаметы. Образование тех и других из мерозоитов протекает различно.
Стадии развития кокцидий, дающие начало микрогаметам, называют микрогаметоцитами. При развитии их, сопровождающемся энергичным ростом, деление ядер начинается на самых ранних стадиях роста, оно совершается гораздо чаще, чем при шизогонии (рис. 72). В результате в цитоплазме располагается несколько сот мелких ядер. Микрогаметы образуются путем вытягивания отдельных ядер. Тело их почти целиком состоит из веретеновидно вытянутого ядра с очень тонким периферическим слоем цитоплазмы. На переднем конце каждой зрелой гаметы имеются два жгута. Длина их очень мала - 4-3 мк. Гаметы подвижны. Большая часть цитоплазмы микрогаметоцита не идет на образование гамет, а остается неиспользованной в виде большого так называемого остаточного тела.
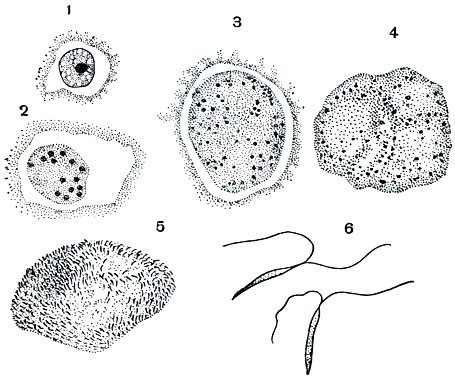
Рис. 72. Развитие микрогамет Eimeria magna: 1 - молодой микрогаметоцит с неразделившимся ядром; 2-4 - стадии роста микрогаметоцита, сопровождающиеся делением ядер; 5 - формирование микрогамет; 6 - микрогаметы, каждая снабженная двумя жгутиками
Совсем иначе происходит развитие женских половых клеток - макрогамет (рис. 73). Происходит рост цитоплазмы, не сопровождающийся делением ядра, размеры которого увеличиваются. Ядро приобретает пузыревидное строение, в центре его расположено крупное ядрышко. В цитоплазме по мере роста происходит накопление большого количества гранул. Макрогаметы сначала имеют сферическую форму, к концу развития они становятся овальными. Часть гранул цитоплазмы приближается к поверхности и располагается периферически в один слой. За счет этих гранул в конце развития макрогаметы образуется оболочка. На одном из полюсов макрогаметы в оболочке остается отверстие (микропиле), через которое проникают микрогаметы.
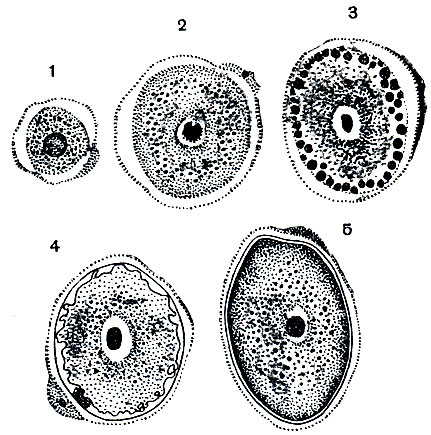
Рис. 73. Развитие макрогамет Eimeria magna: 1-5 - последовательные стадии роста и формирования макрогаметы
Следующий важный этап развития кокцидий - это процесс оплодотворения. Подвижные микрогаметы приближаются вплотную к макрогаметам. Одна микрогамета проникает через микропиле в макрогамету, причем ядра гамет сливаются, образуя характерную веретеновидную фигуру. В момент оплодотворения формируется вторая (внутренняя) оболочка, микропиле закрывается особой слизистой пробкой и зигота попадает в просвет кишечника. Обладающая двумя оболочками зигота получает название ооцисты.
На этом заканчивается эндогенная часть цикла, так как в кишечнике в отсутствие кислорода ооцисты эймерий кролика дальше развиваться не могут.
Экзогенная часть цикла сводится к процессу спорогонии и осуществляется вне тела хозяина. Из кишечника наружу ооцисты выходят вместе с фекальными массами. Только что вышедшие из кишечника ооцисты имеют овальную форму (рис. 70). Все протоплазматическое содержимое отделено от оболочки и занимает центральную часть ооцисты. Споруляция сводится к тому, что после двукратного деления ядра протоплазматическое содержимое ооцисты распадается на четыре одноядерных споробласта. Каждый из них выделяет свою оболочку, превращаясь в спору. Обычно не все протоплазматическое содержимое ооцисты идет на образование споробластов, часть его остается неиспользованной в виде остаточного тела.
В каждой споре ядро делится еще один раз, и образуются два червеобразных одноядерных спорозоита. На этом процесс спорообразования заканчивается. Размеры зрелых ооцист Eimeria magna: длина 25-40 мк, ширина 18-30 мк.
Таким образом, зрелая, способная к заражению ооциста содержит четыре споры, с двумя спорозоитами каждая.
При попадании зрелой ооцисты в кишечник кролика под воздействием кишечного сока пробка ооцисты разрушается, спорозоиты выходят из оболочки спор и проникают в клетки эпителия кишечника, начиная новую, эндогенную часть цикла.
В рассмотренном выше жизненном цикле кокцидий эймерий следует различать три основных этапа, из которых два первых относятся к эндогенной, а третий - к экзогенной части цикла.
Цикл начинается с нескольких поколений бесполого размножения - шизогонии.
Это первый этап. Биологическое значение его очевидно: это увеличение числа особей паразита в данном экземпляре хозяина. Благодаря шизогонии количество паразитов возрастает в огромной степени. У кролика, проглотившего даже небольшое количество ооцист, благодаря шизогонии через короткий срок в кишечнике окажется огромное количество паразитов.
Второй этап - это образование половых клеток и оплодотворение. Он завершается формированием тех стадий жизненного цикла паразита (в данном случае ооцист), при помощи которых происходит распространение паразита на новые особи хозяина, т. е. расселение вида.
Третий этап - спорогония. Он приводит к тому, что "расселительные" стадии жизненного цикла становятся способными к инвазированию (заражению) хозяина. Зрелые ооцисты кокцидий обладают большой стойкостью к действию различных неблагоприятных факторов среды. Наличие двух оболочек является не только механической, но и химической защитой протоплазмы паразита. Это чрезвычайно затрудняет борьбу с кокцидиями, так как применение большинства распространенных дезинфицирующих средств оказывается неэффективным или малоэффективным. Ооцисты очень долго сохраняют жизнеспособность: в течение многих месяцев, а в некоторых случаях до года и более.
Кокцидий - паразиты с очень узкой специфичностью. Почти все виды их строго приурочены к одному виду хозяина и не могут паразитировать даже в близких видах. Например, кокцидий кролика не могут заражать зайцев, а кокцидий зайцев - развиваться в кроликах. В пределах организма хозяина кокцидий локализуются в строго определенных органах и даже частях органов. Например, Е. М. Xейсиным было доказано, что у кролика разные виды кокцидий локализуются в строго определенных отделах кишечника (рис. 74).
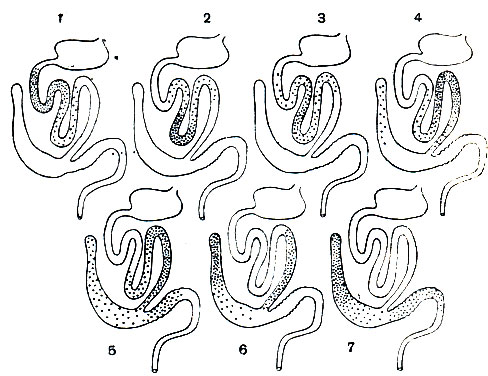
Рис. 74. Локализация разных видов кокцидий рода Eimeria в кишечнике кролика: 1 - Eimeria media; 2 - Е. irresidua; 3 - Е. реrforans; 4 - Е. magna; 5 - Е. intestinalis; 6 - Е. coecicola; 7 - Е. piriformis
Как уже отмечалось, кокцидий приносят большой вред. Следует отметить, что большинство кокцидиозов (заболеваний, вызываемых кокцидиями) поражает молодых птиц и млекопитающих, это главным образом "детские болезни", хотя и взрослые животные могут заражаться кокцидиями.
Ряд видов кокцидий (Eimeria tenella, Е. praecox и др.) поражает кур (преимущественно цыплят), вызывая нередко гибель молодняка.
Крупный рогатый скот также страдает от кокцидий, причем заболевает преимущественно молодняк (Eimeria zurni, Е. smithi, Isospora aksaica).
В рыбоводных хозяйствах (особенно на Украине) кокцидиозы неоднократно поражали карпов (Eimeria cyprini), что также часто приводило к гибели рыб.
Борьба с кокцидиозами - важная задача животноводства и ветеринарии. Она строится главным образом на профилактических мероприятиях, направленных на предохранение животных от заражения.
Отряд кровяные споровики (Haemosporidia)
Кровяные споровики, как показывает и само название отряда, приспособились к паразитированию в крови позвоночных животных, главным образом млекопитающих и птиц. Они являются внутриклеточными паразитами. Место локализации паразитов - кровяные клетки. Число видов кровяных споровиков превосходит сотню.
Жизненные циклы кокцидий и кровяных споровиков очень сходны. Они слагаются из тех же основных этапов: закономерной смены шизогонии, развития гамет, оплодотворения и спорогонии. В отличие от кокцидий, здесь совершенно отсутствуют экзогенные стадии, половая и бесполая части цикла распределяются между двумя хозяевами, из которых один является позвоночным (бесполая часть цикла), а другой - беспозвоночным, обычно насекомым (половая часть цикла и спорогония).
В этих условиях защитные оболочки ооцист и спор, столь характерные для кокцидий, лишены биологического значения и в процессе эволюции исчезают.
Кровяные споровики имеют очень большое практическое значение для медицины, так как к этому отряду относится возбудитель тяжелого заболевания человека - малярии. Существует четыре вида рода Plasmodium, вызывающие малярию (P. vivax, P. malaria, P. falciparum, P. ovale)*. Клиническая картина болезни, вызываемой разными видами паразита, несколько различна. О некоторых из этих различий будет сказано ниже. Жизненный цикл всех четырех видов протекает сходно, что позволяет рассмотреть его для всего рода Plasmodium.
* (Первые три вида распространены широко, Plasmodium ovale встречается редков тропическом поясе Африки и Азии.)
Человек является для Plasmodium хозяином, в котором протекают стадии бесполого размножения (шизогония). Половой процесс и спорогония осуществляются в переносчике, каким для возбудителей человеческой малярии служат виды комаров, относящиеся к роду Anopheles.
Обратимся к рассмотрению жизненного цикла Plasmodium и начнем его с той же стадии спорозоита, как мы это делали выше для кокцидий (рис. 75). Спорозоит проникает в кровь человека при укусе зараженным комаром. Он представляет собой очень маленькую одноядерную веретеновидно вытянутую клетку длиной 10-15 мк при ширине всего в 1 мк. С током крови спорозоиты разносятся по телу человека и попадают в печень. Здесь они активно внедряются в клетки печени и превращаются в шизонтов, достигающих значительной величины. Каждый шизонт распадается в процессе шизогонии на большое количество мерозоитов, которые могут вновь внедряться в клетки печени. После одного или большего числа бесполых поколений в печени шизонты дают мерозоитов, судьба которых оказывается иной. Они внедряются не в печеночные клетки, а в эритроциты крови, где дают начало шизонтам гораздо меньших размеров, чем шизонты, развивающиеся в печени. Начинается период шизогонии в кровяном русле. Число мерозоитов, на которые распадаются шизонты крови, относительно невелико, обычно 8-16.
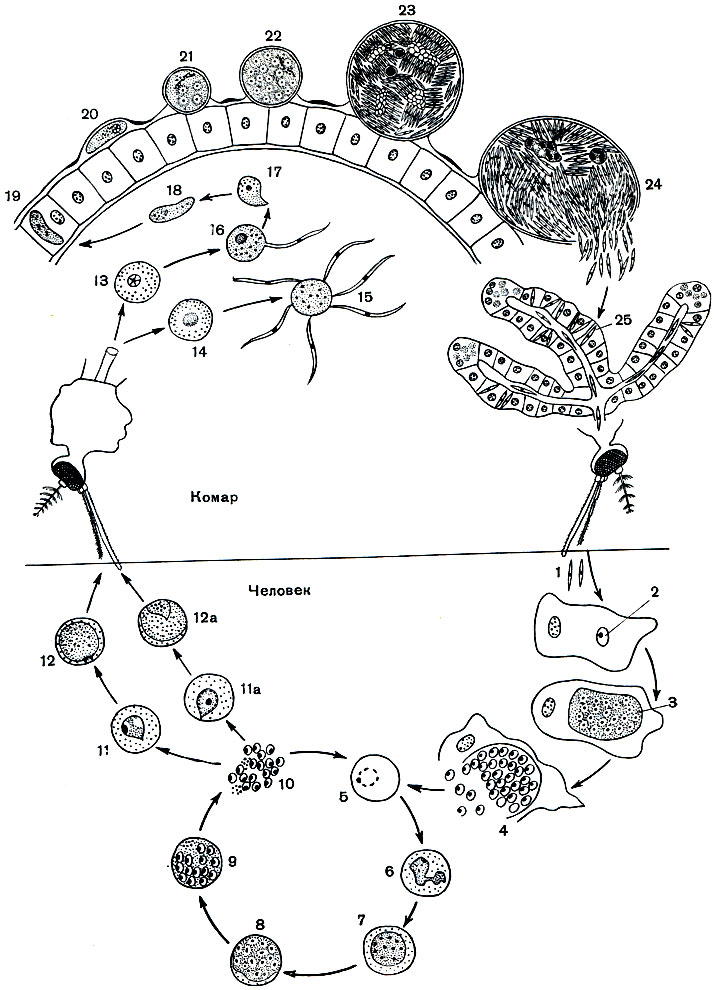
Рис. 75. Цикл развития кровяного споровика Plasmodium vivax (возбудителя малярии): 1 - спорозоиты; 2-4 - бесполое размножение (шизогония) паразита в клетках печени; 2 - спорозоит, внедрившийся в печеночную клетку; 3 - растущий шизонт с многочисленными ядрами; 4 - шизонт, распадающийся на мерозоиты; 5-10 - бесполое размножение (шизогония) в красных кровяных клетках (эритроцитах); 5 - молодой шизонт в форме кольца; в - растущий шизонт с псевдоподиями; 7,8 - дальнейшие стадии роста шизонта, деление ядер; 9 - распад шизонта на мерозоиты; 10 - выход мерозоитов из эритроцита; 11 - молодой макрогаметоцит; 11а - молодой микрогаметоцит; 12 - зрелая макрогамета; 12а - зрелый микрогаметоцит; 13 - макрогамета; 14 - микрогаметоцит; 15 - образование микрогамет; 16 - копуляция микро- и макрогаметы; 17 - зиготы; 18 - подвижная зигота (оокинета); 19 - оокинета, проникающая через стенку кишечника комара; 20 - оокинета, прикрепившаяся к наружной стенке кишечника комара и превращающаяся в ооцисту; 21, 22 - растущая ооциста с делящимися ядрами; 23 - зрелая ооциста со спорозоитами; 24 - спорозоиты, покидающие оболочку ооцисты; 25 - спорозоиты в слюнной железе комара
Шизонты на ранних стадиях развития имеют характерную форму колечка благодаря тому, что центр их тела занят большой вакуолей (табл. 4). По мере роста шизонта вакуоля исчезает и он приобретает форму маленькой амебы. Наблюдения за живыми паразитами показали, что шизонтам свойственно довольно активное амебоидное движение внутри красного кровяного тельца. Мерозоиты, получающиеся в результате размножения шизонта, представляют собой мелкие клетки овальной формы диаметром около 2 мк. По завершении шизогонии оболочка эритроцита лопается и мерозоиты оказываются плавающими в плазме крови. По мере роста шизонт поглощает содержимое эритроцита. Красный пигмент крови (гемоглобин) при этом изменяется химически и превращается в темно-коричневый, почти черный меланин, откладывающийся в цитоплазме паразита в виде мелких зерен. В момент шизогонии пигмент выбрасывается из тела паразита и остается в крови в виде остаточного тела.
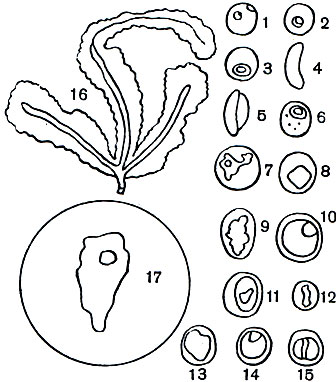
Таблица 4. Разные стадии жизненного цикла возбудителей человеческой малярии в эритроцитах крови человека (1-15) и в слюнных железах комара-переносчика Anopheles (16, 17) с препаратов, окрашенных по Гимза-Романовскому: 1-5 - возбудитель тропической малярии Plasmodium falciparum: 1-начало роста шизонта в эритроците, в одном эритроците два мелких 'кольца', в каждом из них по одному окрашенному в красный цвет ядру; 2, 3-более крупные 'кольца', одно (2) с двумя, другое (3) с одним ядром; 4-бобовидная макрогамета; 5-микрогаметоцит; 6-11 - возбудитель трехдневной малярии P. Vivax; 6-молодой шизонт в начале роста в форме кольца; 7-более крупный одноядерный шизонт амебоидной формы; 8-растущий шизонт с четырьмя ядрами; 9-выросший шизонт, распадающийся на одноядерные мерозоиты; 10-макрогамета; 11-микрогаметоцит; 12 -15 - возбудитель четырехдневной малярии P. malariae: 12-растущий шизонт в форме пояска; 13-выросший шизонт, распадающийся на одноядерные мерозоиты; 14-макрогамета; 15-микрогаметоцит; 16- продольный разрез через слюнную железу комара Anopheles maculipennis; в клетках железы, обладающих крупными ядрами, расположены многочисленные веретеновндные спорозоиты, часть их вышла в проток железы. 17-мазок слюнной железы комара Anopheles superpictus, зараженного малярийным плазмодием; в центре крупная клетка железы с ядром, вокруг нее многочисленные веретеновндные одноядерные спорозоиты
Шизогония в крови осуществляется с большой правильностью в отношении времени роста и размножения шизонта. У видов Plasmodium vivax, P. falciparum, P. ovale шизогония происходит каждые 48 часов, у P. malaria - каждые 72 часа. С моментом завершения шизогонии связаны характерные клинические явления: повышение температуры до 40° С и выше, озноб. Эти явления обусловлены тем, что при разрушении эритроцитов в кровь поступают токсические вещества - результат жизнедеятельности паразита, которые в период роста шизонта были изолированы оболочкой эритроцита.
Мерозоиты вновь внедряются в эритроциты, и весь цикл шизогонии начинается сначала. После каждой шизогонии число паразитов в крови все более и более возрастает.
Шизогония, однако, идет не беспредельно. Через несколько циклов бесполого размножения часть внедрившихся в эритроциты мерозоитов превращается в стадии развития, подготовительные к образованию половых клеток, которые получают название гаметоцитов. Гаметоциты образуются двух категорий: микро- и макрогаметоциты. Первые в дальнейшем дают начало микрогаметам (мужским половым клеткам), вторые - макрогаметам (женским половым клеткам). Между этими двумя категориями гаметоцитов имеются некоторые небольшие различия в строении, которые в основном сводятся к тому, что макрогаметоциты богаче снабжены резервными запасными включениями.
Для дальнейшего развития гаметоциты должны попасть в кишечник комара анофелес, что происходит при сосании им крови больного малярией. Судьба микро-и макрогаметоцитов различна. Каждый микрогаметоцит в желудке комара дает начало 4-8 нитевидным подвижным микрогаметам. Макрогаметоциты без деления преобразуются в макрогаметы. В просвете желудка комара происходит слияние мужских и женских гамет (оплодотворение), и образуется овальной формы зигота. Зигота (которая здесь благодаря своей подвижности получает название оокинеты) прободает желудок и закрепляется на его стенке, обращенной в сторону полости тела, превращаясь в ооцисту. Здесь она одевается оболочкой (эта оболочка несравнима с оболочкой ооцист кокцидий: у плазмодия она образуется за счет тканей комара), после чего начинается очень быстрый рост, в результате которого объем ооцисты увеличивается во много сот раз. По мере роста протоплазмы ооцисты происходит многократное деление ядра. Процесс развития ооцисты на кишечнике завершается образованием длинных (до 14 мк) спорозоитов, очень тонких (1 мк) одноядерных клеток (но спор, как у кокцидий, здесь не образуется). В каждой ооцисте формируется огромное количество (до 10 тыс. в одной ооцисте) спорозоитов. Зрелая ооциста лопается, и спорозоиты попадают в заполненную гемолимфой (кровью) полость тела комара. Интересна их дальнейшая судьба. Спорозоиты активно перемещаются, совершая червеобразные движения. Перемещение их не беспорядочно. Они направляются в слюнные железы комара, в которых собираются в огромных количествах (табл. 4).
Анофелесы, у которых спорозоиты заполняют слюнные железы, являются источником заражения человека. Спорозоиты при укусе проникают в кровяное русло человека, и цикл развития плазмодия начинается вновь.
Сколько времени требуется для завершения цикла развития малярийного плазмодия в комаре и при каких условиях среды он протекает?
Быстрота развития плазмодия в комаре в большой степени зависит от температуры. Цикл развития Plasmodium vivax при 17-18° С завершается в комаре в течение трех недель. При более высоких температурах среды он протекает значительно быстрее. Например, при 25-27° С для развития плазмодия требуется лишь одна неделя. Если температура окружающей среды ниже 15-17° С, то развитие его начинается нормально, происходит оплодотворение и образование оокинеты, но спорозоиты не формируются.
Указанные закономерности зависимости развития человеческих видов Plasmodium в переносчике от температуры объясняют основные факты географического распространения малярии. Ареал комаров рода Anopheles значительно шире, чем ареал возбудителя малярии. Комары могут жить и развиваться при температурах более низких, чем это необходимо для развития Plasmodium. Поэтому наличие Anopheles далеко не всегда означает возможность существования в данной местности малярии.
Малярия - болезнь преимущественно теплых стран, обладающих необходимыми условиями для развития комаров анофелесов. Личинки этих насекомых, развивающихся с полным превращением, живут в воде, в мелких стоячих водоемах (рис. 76), сохраняя, однако, воздушное дыхание. На заднем конце тела личинки расположены дыхальца, через которые воздух проникает в их дыхательную (трахейную) систему. Личинки периодически поднимаются на поверхность воды для дыхания. Куколки комаров тоже живут в воде и дышат атмосферным воздухом. Лишь взрослый комар является воздушным насекомым. Яйца анофелесы откладывают в воду.
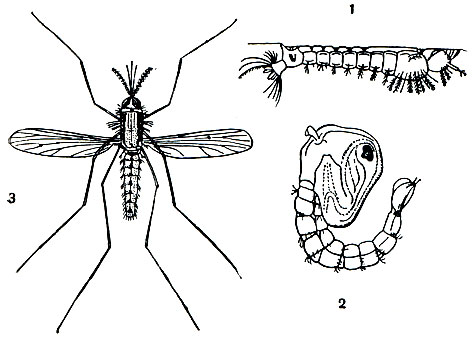
Рис. 76. Комар Anopheles maculipennis, переносчик возбудителя малярии: 1 - личинка в момент захвата атмосферного воздуха через дыхальца у поверхности воды; 2 - куколка; 3 - взрослая самка
Борьба с малярией осуществляется двумя основными путями. Первый - это лечение маляриков специальными лекарственными средствами (хинин, акрихин, плазмоцид и др.). Отсутствие возбудителя в крови людей делает невозможным заражение переносчиков и, следовательно, распространение малярии.
При этом необходимо отметить одну важную особенность, имеющую существенное значение для борьбы с малярией. Дело в том, что большинство специфичных лекарственных средств убивает шизонтов в крови. Гаметоциты же являются гораздо более стойкими и могут сохраняться в эритроцитах продолжительное время в отсутствие шизонтов. Больной малярией после лечения становится вполне здоровым и часто не подозревает, что является носителем гаметоцитов. Он может явиться источником заражения других людей, если комар насосется его крови с гаметоцитами. Поэтому в малярийных местностях необходим систематический контроль за всеми переболевшими малярией лицами. У них периодически берутся мазки крови и исследуются на носительство гаметоцитов.
В случае обнаружения гаметоносительства проводится специальный курс лечения лекарственными препаратами, разрушающими гаметоцитов.
Второй путь борьбы с малярией - это уничтожение переносчиков. Методы этой борьбы разнообразны. В основном это уничтожение личинок путем заливки водоемов керосином или нефтью. Личинки, поднимающиеся на поверхность для дыхания, наталкиваются на пленку нефти, которая, заполняя трахеи, вызывает их гибель.
Существуют и другие методы борьбы с личинками. Особенно интересен биологический метод борьбы с использованием небольших живородящих рыбок - гамбузий (Gambusia affinis, рис. 77). Эти теплолюбивые рыбки (родина их Америка) особенно охотно поедают личинок комаров. Водоемы, являющиеся местами выплода личинок Anopheles, заселяют гамбузиями, которые быстро уничтожают всех личинок. Этот способ применяют в теплых странах, так как гамбузии не могут жить в холодных водоемах.

Рис. 77. Рыбка Gambusia affinis. Слева самец, справа самка. Естественная величина
Борьба со взрослыми летающими комарами представляет большие трудности. Ее проводят в зимнее время на местах зимовок. Анофелесы скопляются в значительных количествах в хлевах, так как, кроме человека, они охотно нападают на рогатый скот. Здесь их и уничтожают, применяя для этого различные инсектициды. В дореволюционной России малярия была широко распространена и для многих областей страны была массовым заболеванием. Десятки тысяч людей болели малярией. Особенно неблагополучными были некоторые районы Кавказа (окрестности Батуми, Сухуми, Сочи), Средней Азии, Поволжья. Большие эпидемические вспышки возникали и в первые годы Советской власти, особенно в период гражданской войны. Органы здравоохранения Советского Союза провели огромную работу по борьбе с малярией. Этим в первую очередь занималась широкая сеть специальных учреждений - малярийных станций, развернутых во всех центральных и южных районах страны. Эта борьба, шедшая по разным направлениям, увенчалась полным успехом. В настоящее время малярия как массовое заболевание на территории нашей Родины не существует.
В других странах Европы малярия как массовое заболевание тоже ликвидирована. Но в азиатских и африканских странах (в особенности в Индии с ее влажным и теплым климатом) малярия - страшный бич населения, от которого страдают десятки миллионов людей. Понадобятся еще большие усилия, чтобы окончательно освободить человечество от этого тяжелого заболевания.
Кровяные споровики, так же как и кокцидии,- паразиты с узкой специфичностью. Виды рода Plasmodium, паразитирующие в человеке, в других млекопитающих развиваться не могут. Благодаря этому не существует природных очагов малярии, не связанных с человеком, какие мы видим, например, у вызываемой трипаносомами сонной болезни.
Среди многочисленных видов гемоспоридий известное практическое значение имеет Plasmodium gallinaceum, паразитирующий в крови домашней курицы. Этот паразит встречается в тропических странах, где он может вызывать тяжелые эпизоотии среди кур. Особенно тяжело протекает заболевание цыплят, обычно приводящее к гибели. Переносчиками P. gallinaceum являются комары рода Aedes.
P. gallinaceum широко используют в лабораторной практике для изучения жизненного цикла, так как исследование человеческих видов (особенно шизогонии, протекающей во внутренних органах) представляет большие трудности.
Приведенный выше обзор основных групп споровиков показывает их глубокую приспособленность к паразитическому образу жизни. Приспособления эти разнообразны. Характерный для большинства споровиков способ бесполого размножения - множественное деление (шизогония) обеспечивает быстрое увеличение числа паразитов в хозяине. При наличии в жизненном цикле экзогенных стадий вырабатываются специальные приспособления защитного характера, допускающие длительное существование паразита в неблагоприятных условиях внешней среды. Примером могут служить ооцисты кокцидий. Наконец, наиболее сложным и биологически совершенным приспособлением к паразитизму является полное выключение экзогенных стадий и появление двух хозяев, в одном из которых осуществляется шизогония, а в другом - половой процесс и спорогония. По этому пути пошла эволюция кровяных споровиков.
Среди какой группы свободноживущих простейших следует искать корни происхождения споровиков? Весьма вероятно, что они взяли начало от жгутиконосцев. Серьезным доказательством в пользу этой точки зрения является наличие в жизненном цикле большинства споровиков жгутиковых стадий. Например, микрогаметы кокцидий по своему строению являются как бы небольшими жгутиконосцами, снабженными двумя жгутиками. Характер полового процесса споровиков - копуляция - весьма близок к тому, что наблюдается у жгутиконосцев. Наконец, как мы видели выше, представители класса жгутиконосцев довольно легко переходят к паразитизму.
Глубокая приспособленность к паразитическому образу жизни обеспечивает споровикам широкое распространение и большое видовое разнообразие.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'