7. Одноклеточные организмы. Мопега и Protista
Ключевые вопросы
Какие признаки характерны для большинства подразделений царства Монера?
Какие факторы внешней среды, существовавшей ранее на Земле, благоприятствовали возникновению разнообразия среди прокариот?
Существуют ли другие факторы, помимо условий окружающей среды, которые могли повлиять на эволюцию протист? Каковы основные подразделения протист?
Схема классификации, состоящая из пяти царств, основана на точке зрения, что все многоклеточные организмы - растения, грибы и животные - возникли много лет назад из эукариотических одноклеточны х организмов, существовавших в ту пору, и которые, возможно, были бы теперь классифицированы в царство Protista.
Эти протисты, в свою очередь, произошли от ранних одноклеточных прокариотических организмов, вероятно сходных с современными представителями царства Мопега (рис. 7-1).
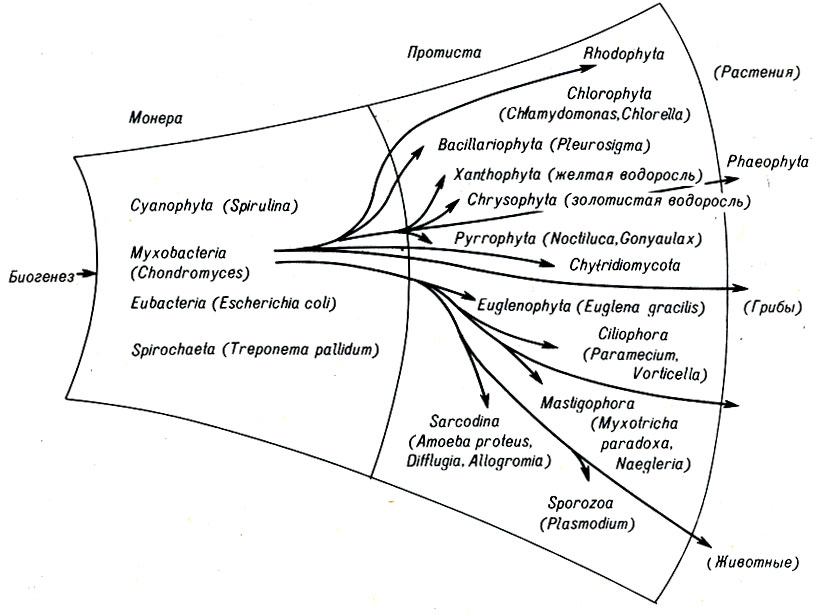
Рис. 7-1. Подробный обзор двух царств, демонстрирующий положение основных типов и представителей, обсуждаемых в тексте
Чтобы понять, как могли осуществиться эти сложные эволюционные преобразования в далеком прошлом, необходимо рассмотреть некоторые из основных свойств прокариот, прежде чем обсуждать признаки, способствовавшие возникновению протист.
7.1. Прокариоты царства Мопега подразделяются на четыре типа: Eubacteria ["истинные" бактерии], Myxobacteria, Cyanophyta [сине-зеленые водоросли] и Spirochaeta [спирохеты]
Тип Eubacteria содержит огромное количество видов. Они еуществуют в форме палочек, сфер (кокки) или спиралей и имеют плотные клеточные стенки. Некоторые из них неподвижны, другие способны передвигаться с помощью одного или нескольких бактериальных жгутиков. Бактериальные жгутики представляют собой закрученные нити, состоящие из белка флагеллина, связанные с бактериальной мембраной. В противоположность этому жгутики эукариот состоят из осевой нити, образованной комплексом микротрубочек, выступающей из клетки и окруженной натянутой плазматической мембраной.
У эубактерий в процессе эволюции возникло широкое разнообразие способов получения энергии из химических соединений окружающей среды. Едва ли существует органическое соединение, которое не могло бы расщеплять и использовать бактерии какого-либо вида. Среди эубактерий существуют аэробные (использующие кислород) формы. Эти бактерии имеют связанные с мембранами дыхательные пигменты, подобные дыхательным ферментам в митохондриях эукариотических клеток. Некоторые эубактерий являются фотосинтезирующими микроорганизмами.
Тип Myxobacteria представляет собой другую большую группу организмов, родственную сине-зеленым водорослям. Они имеют относительно тонкие клеточные стенки, и некоторые виды обнаруживают интересный способ "скользящего " движения, сопровождающегося часто изменениями формы клеток. В настоящее время неизвестно, связан ли этот вид движения с движением цитоплазмы или с амебоидным движением, наблюдаемым у протист.
Представители типа Cyanophyta (сине-зеленые водоросли) сходны с миксо-бактериями во многих отношениях, но они содержат хлорофилл и другие пигменты, придающие им характерную окраску. У многих сине-зеленых водорослей фотосинтетические пигменты локализованы в слоистых мембранах. Некоторые сине-зеленые водоросли могут фиксировать атмосферный азот, т. е. синтезировать азотсодержащие вещества из газообразного азота воздуха.
Многие нитевидные сине-зеленые водоросли передвигаются путем скольжения. Одна из наиболее интересных форм движения наблюдается у Spirulina, которая представляет собой нить, свернутую в спираль. В зависимости от направления скручивания спирали цепь клеток движется вперед или назад. Нити растут за счет деления составляющих их клеток. По мере добавления новых клеток нити сплетаются и в конце концов распадаются на более короткие фрагменты. Постоянный рост, разрыв нитей и случайное передвижение - все это способствует их распространению.
Представители типа Spirochaeta (спирохеты) - организмы спиралеобразной формы, скрученные вокруг осевой нити.
Они часто имеют нитевидную или фибриллярную структуру.
Большинство таких микроорганизмов - безвредные симбионты. Их можно обнаружить в пищеварительном тракте насекомых и млекопитающих. Однако одна спирохета далеко не безвредная. Treponema pallidum вызывает сифилис, венерическое заболевание, которое принесло большие страдания человечеству.
7.2. Полагают, что эволюция прокариот осуществлялась путем естественного отбора в популяциях, характеризующихся большим генетическим разнообразием
Генетические исследования на бактериях показали, что мутации встречаются довольно часто в больших популяциях (одна на 106-109 делений). Принимая во внимание короткое время генерации многих бактерий (от 20 мин до нескольких часов), мутации могут возникать один раз в течение нескольких часов или суток. Врачи, лечившие бактериальные инфекции с помощью пенициллина, не знали, насколько быстро могут развиваться пенициллиноустойчивые линии мутантных микроорганизмов. На заре применения пенициллиновой терапии инфекционные заболевания часто лечились слишком короткое время и лейкоциты крови не успевали разрушать все инфекционные бактерии. (Пенициллин не убивает бактерии, он почти полностью останавливает их размножение, позволяя лейкоцитам хозяина фагоцитировать их.) Когда терапия завершается слишком быстро, инфекция вспыхивает вновь и некоторые выжившие бактерии оказываются пени-циллиноустойчивыми.
По-видимому, условия, существовавшие на Земле в далеком прошлом, были более благоприятными для возникновения мутаций. В тот период в атмосфере содержалось небольшое количество кислорода и Земля не имела защитного озонового экрана, поглощающего ультрафиолетовое излучение и сформировавшегося позднее из кислорода. Известно, что ультрафиолетовое излучение является мощным фактором возникновения мутаций. Ранняя эволюция бактерий, по-видимому, была в значительной степени ускорена за счет распространения индуцированных ультрафиолетом мутаций, и некоторые из этих ранних бактерий далекого прошлого, возможно, дали начало клеткам, которые в настоящее время мы рассматриваем как эукариотические.
7.3. Эволюция эукариотических Protista из прокариотических Моnеrа, вероятно, осуществлялась за счет двух процессов, естественного отбора и наследственного симбиоза
Существуют большие различия в структуре и в организации между наиболее сложными прокариотами и простейшими эукариотами.
В прошедшее десятилетие были выдвинуты теории, позволившие в общих чертах понять, как могли эволюционировать протисты.
Многие исследователи считают, что некоторые характерные признаки эукариотических клеток развились в результате наследственного симбиоза. Многие формы симбиоза не являются наследственными; в этом случае оба организма, которые существуют вместе для их взаимной выгоды, размножаются независимо. Их потомки вновь объединяются друг с другом на более поздней фазе жизненного цикла. При наследственном симбиозе организмы двух видов проявляют более полную ассоциацию друг с другом и существуют вместе из поколения в поколение. Paramecium bursaria (рис. 7-2), например, является простейшим, в цитоплазме которого обитают клетки зеленой водоросли Chlorella, Экспериментальным путем P. bursaria может быть лишена своих Chlorella-симбионтов и последние также могут быть изолированы и выращены отдельно. Если парамеции поместить, со свободно-живущей Chlorella, которая была взята из водоема, то они заглатывают водоросли и затем переваривают их. Если, однако, их накормить симбиотической Chlorella, которая была удалена из P. bursaria, водоросли не будут перевариваться. Более того, однажды проглоченные, симбионты будут воспроизводиться до определенного числа особей, и такое ограничение их количества генетически детерминировано хозяином. Эти результаты дают возможность предполагать, что либо протисты "узнают" своих симбионтов, либо симбионты обладают каким-то свойством, позволяющим им избежать переваривания.
Рис. 7-2. Paramecium bursaria - протиста, содержащая много симбиотических водорослей Chlorella (А)
В обоих случаях, когда Paramecium bursaria делится, ее дочерние клетки наследуют симбиотические водоросли Chlorella и вместе с ними механизм, регулирующий их численность.
Многие протисты имеют бактерий, живущих симбиотически на их поверхности или в цитоплазме. Примером является Myxotricha paradoxa-жгутиковое, живущее в кишечнике австралийского термита. Когда это простейшее исследовали под электронным микроскопом, было установлено, что его жгутики, считавшиеся эукариотическими, являются бактериальными и принадлежат спирохетам, пристроившимся в углублениях клеточной поверхности. Любопытно, что эти жгутики служат для движения хозяина. Однако синхронность их биения обусловлена не каким-либо видом нервной координации, а лишь гидродинамическим взаимодействием между соседними жгутиками. Фактически Myxotricha имеет два вида спирохет-симбионтов и по крайней мере еще один вид бактериальных симбионтов.
Чтобы объяснить эти наблюдения, а также эволюцию древних протист, предположили, что самые первые эукариоти-ческие клетки и, следовательно, первые протисты были способны заглатывать различные виды прокариот; некоторые из них были устойчивы к перевариванию и стали эндосимбионтами. Поскольку, как известно, простейшими клетками, способными заглатывать другие клетки, являются амебоидные, было выдвинуто предположение, что этими первыми эукариотическими протестами моглц быть очень простые анаэробные амебы. Такие организмы существуют в настоящее время, и многие из них содержат бактерии и спирохеты, живущие в них, как симбионты. Примером является один вид гигантской амебы Pelomyxa palustris, которая обитает в иле на дне мелких прудов в анаэробных условиях. В цитоплазме этой амебы содержатся симбиотические бактерии.
7.4. Митохондрии у протист могли возникнуть из аэробных бактерий, существовавших в виде эндосимбионтов
Фактически во всех клетках цитоплазма содержит все ферменты, необходимые для катализа реакции анаэробного гликолиза. Эта одна из простейших форм энергетического метаболизма, возможно, и была именно той, которая использовалась ранее амебами и другими протистами, жившими в анаэробных условиях окружающей среды (как некоторые существуют до сих пор). В настоящее время широко распространено мнение, что у определенных видов амеб возникла способность извлекать пользу из условий, богатых кислородом, и более эффективно использовать пищу за счет приобретения аэробных бактерий в качестве эндосимбионтов. Эти бактерии, сходные во многих отношениях с бактериями, существующими сегодня, могли превратиться в митохондрии - структуры , имеющие признаки, общие с бактериями. Митохондрии содержат одну нить ДНК (подобно бактериальной "хромосоме"), в которой локализованы гены, контролирующие выработку некоторых ферментов, обеспечивающих клеточное дыхание (аэробный гликолиз), и большинства структурных белков, необходимых для формирования новых дочерних митохондрий.
7.5. Центриоли, жгутики и реснички могли возникнуть из спи-рохет-эндосимбионтов
Центриоль представляет собой структуру, обнаруживаемую у многих протист и почти во всех клетках животных. Она существует в нескольких формах и служит либо в качестве базального тельца, из которого берут начало ресничка или жгутик, либо как организующий центр, состоящий из микротрубочек и принимающий участие в формировании митотического веретена и лучистых сфер. В отличие от протист, которые обычно имеют центриоли или их производные, у большинства клеток высших растений они отсутствуют.
В процессе эволюции примитивные амебоидные эукариоты, возможно, могли приобрести, как и в случае с митохондриями, наследуемого симбионта, превратившегося в центриоль. В этом случае симбионт, по-видимому, является организмом, напоминающим некоторые сегодняшние спирохеты. В дальнейшем такие симбионты должны были бы развиваться несколькими путями в клетках своих хозяев, прежде чем стать центриолеподобной органеллой древних форм эукариот. Современные жгутиковые имеют самые разнообразные формы центриолей и их производных. У многих видов протист центриоли и базальные тельца являются равноценными и могут воспроизводиться в клетке.
Одним из аргументов, ставящих под сомнение гипотезу об эндосимбиотиче-ском происхождении центриолей и их производных, является тот факт, что в центриолях не содержится ДНК. Это может означать, что она отсутствует, что ее количество слишком мало, чтобы быть обнаруженной с помощью современных методов, или что ДНК каким-то образом стала частью генома хозяина.
Каково бы ни было происхождение центриолей, обладание одним или несколькими эукариотическими жгутиками, образовавшимися из центриолей, дало жгутиковым преимущества движения. Внешне симбиотические взаимоотношения могут напоминать взаимоотношения между человеком и лошадью: транспорт в обмен на пищу и приют. Исходя из разнообразия жгутиковых и сохранения жгутиковых стадий в развитии высших организмов, мы можем предположить, что жгутик был важным селективным преимуществом в процессе эволюции, т. е. организмы, имеющие жгутики, могли легко распространяться в благоприятных условиях среды и в борьбе за существование выживали более часто, чем их безжгутиковые родственники.
7.6. Процесс митоза установился рано в истории протист и явился основой для их дальнейшей эволюции
Митоз наблюдается при размножении всех эукариотических клеток, как у протист, так и у многоклеточных организмов. Однако среди протист существует значительно большее разнообразие в деталях митоза, чем в любом из трех царств многоклеточных организмов. Поэтому считают, что митоз возник очень давно и его эволюция заняла длительный период времени. Вероятно, для развития достаточно сильно отличающихся механизмов митоза животных и растительных организмов потребовался период времени в миллиард лет. Дивергенция митоза и цитокинеза дает ключ к пониманию того, как могли возникнуть растения на Земле.
Ядро эукариот отличается от нуклеоида прокариот несколькими важными признаками. Прокариоты имеют единственную свободную нить ДНК. В противоположность им эукариоты могут содержать до нескольких сотен хромосом, в которых ДНК связана с гистонами и другими белками. Кроме того, все хромосомы эукариот находятся в пределах ядерной оболочки, которая отсутствует у нуклеоида прокариот. Неизвестно, каким образом появились эукариотическое ядро и процесс митоза, но результат их присутствия очевиден. Протисты смогли претерпеть далеко идущую эволюцию, коль скоро они приобрели механизм для точного и равномерного распределения реплицированных хромосом между дочерними клетками. Возможность такого точного равномерного распределения обеспечивало наличие митотического веретена.
Мейоз и половое размножение в значительной степени обогатили разнообразие популяций благодаря появлению рекомбинации генетического материала. Мейоз существует у представителей всех основных групп протист и, по-видимому, вносит значительный вклад в эволюцию форм этого царства организмов.
7.7. В настоящее время царство Protista содержит более 35 000 видов, чрезвычайно разнообразных по структуре и функции
Протисты обычно подразделяются на девять или более типов (табл. 2). Одним из самых крупных типов является тип Mastigophora, зоофлагелляты. Другой большой и важной группой является тип Sarcodina, который включает амеб, планктонные организмы, такие, как фо-раминиферы и радиолярии, солнечники и многие другие. Также имеются тип Spo-rozoa, тип Ciliophora и тип Pyrrophyta.
Другие типы являются; по-видимому, переходными между протистами и грибами (например, тип Myxomycoeta, слизевики) или между протистами и растениями (тип Chrysophyta включает желтые водоросли; тип Bacillariophyta - диатомовые водоросли и тип Chlorophyta - зеленые водоросли).
| Царство Моnеrа (одноклеточные прокариотические организмы) | Тип Eubacteria (истинные бактерии) Тип Myxobacteria (миксобактерии) Тип Cyanophyta (сине-зеленые водоросли) Тип Spirochaeta (спирохеты) |
| Царство Protista (одноклеточные эукариотические организмы) | Тип Sarcodina (организмы с псевдоподиями) Класс Rhizopoda (голые амебы) Класс Testacea (раковинные амебы Класс Foraminifera (раковинные ретикулоидальные организмы) Класс Radiolaria (ретикулоидальные организмы, имеющие скелет) Класс Actinopoda (солнечники, или "маленькие солнечные животные") Тип Ciliophora (организмы, двигающиеся при помощи ресничек) |
| Царство Protista (одноклеточные эукариотические организмы) | Класс Ciliata (реснитчатые) Класс Suctoria (сосущие - имеют реснички на ювенильной стадии) Тип Mastigophora (зоофлагелляты) Тип Sporozoa (споровики - паразитические, неподвижные или пассивно перемещающиеся) Тип Euglenophyta (эвгленовые, фотогетеротрофные флагелляты) Тип Pyrrophyta (динофлагелляты) *Тип Chrysophyta (золотистые водоросли) *Тип Xanthophyta (желтые водоросли) *Тип Bacillariophyta (диатомовые) *Тип Chlorophyta (зеленые водоросли) |
| *Группы, содержащие многоклеточные формы | |
7.8. Amoeba proteus является современным представителем типа Sarcodina
Протисты, которые двигаются исключительно при помощи псевдоподий, образуют тип Sarcodina. В дополнение к амебам, не имеющим защитного слоя, существует широкое разнообразие раковинных амеф.
Хотя амебоидное движение, вероятно, возникло рано в истории протает, некоторые современные саркодовые, по-видимому, представляют собой реликтовые формы.
Многие современные амебы, такие, как Amoeba proteus (рис. 7-3), имеют сложную функциональную организацию и жизненный цикл.
Рис. 7-3. Вид Amoeba proteus при дифференциальной интерференционной контрастной микроскопии
Amoeba jproteus обнаруживается в незагрязненных озерах и прудах. Она и многие другие виды свободноживущих амеб могут быть собраны при осторожной очистке водных растений в сосуде с водой, взятой из водоема. A. proteus можно видеть невооруженным глазом в виде белого пятнышка на дне сосуда. Ее поверхность покрыта удивительно плотной мембраной, плазмалеммой, на внешней стороне которой находится очень тонкая пушистая мукопротеиновая оболочка, гликокаликс. Цитоплазма содержит тонкие нити F-актина и толстые нити, представляющие собой скопления миозина. Данные структуры используются при сокращении цитоплазмы, образовании псевдоподий и, возможно, также при фагоцитозе (питание клетки) и пиноцитозе (поглощение клеткой воды). Цитоплазма A. proteus содержит митохондрии, комплекс Гольджи, везикулы, преломляющие тельца и кристаллы продукта выделения азота, триурета. Обычно также присутствуют бактериальные эндосимбионты. Ядро бобовидной формы и окружено необычной "сотовидной" ядерной оболочкой, внутри которой находится большое количество ядрышек. A. proteus питается преимущественно инфузориями и жгутиковыми, которых она захватывает псевдоподиями, образующими пищевую чашу. В одних случаях эти пищевые чаши формируются заранее и затем закрываются, если туда попадает добыча. В других случаях амеба вытягивает псевдоподию в направлении добычи и образует пищевую чашу, по размеру и форме соответствующую жертве.
Захваченные организмы окружаются участком плазмалеммы, который превращается в мембрану пищеварительной вакуоли.
Лизосомы сливаются с мембраной пищеварительной вакуоли и доставляют ферменты, переваривающие добычу. Любой непереваренный материал выбрасывается при слиянии мембраны пищеварительной вакуоли с плазмалеммой.
Амебы так малы, что газообмен у них эффективно осуществляется путем диффузии. Азотистые продукты распада, образовавшиеся в результате разрушения белков и аминокислот, могут или диффундировать из клетки (как аммиак или моча), или удаляться через сократительную вакуоль. Эта структура, обнаруживаемая наиболее часто в хвостовой части амебы, постепенно, в течение примерно полминуты, наполняется водой и растворенными в ней веществами цитоплазмы, а затем сокращается и выбрасывает содержимое через временное отверстие в плазма лемме.
Продукты распада нуклеиновой кислоты накапливаются в цитоплазме амебы в виде кристаллов триурета (образованного из разрушенных азотистых оснований) и в виде сферических преломляющих телец (содержащих полифосфаты). По-видимому, амебы редко избавляются от этих необычных метаболических твердых отходов.
A. proteus двигается при помощи псевдоподий, временных выпячиваний, которые могут вытягиваться или втягиваться. Вытягивание псевдоподий происходит вследствие сокращения цитоплазмы, которая перетекает в них.
Поведенческие реакции амеб не ограничиваются вытягиванием и втягиванием псевдоподий. Они определенным образом отвечают на целый ряд стимулов окружающей среды. Например, интенсивный белый или голубой свет, попавший на хвостовую ее часть, заставляет амебу быстро двигаться от света. Облучение концов псевдоподий вызывает их втягивание. На механические стимулы (прикосновение) концы псевдоподий отвечают остановкой, однако задняя, или хвостовая, область клетки довольно нечувствительна к прикосновению. Химические стимулы могут заставить амебу остановиться, быстро двигаться или поглощать какую-то жидкость из окружающей среды путем пиноцитоза. На добычу и вещества, диффундирующие из нее, амебы отвечают формированием пищевой чаши.
Механизмы поведенческих реакций амебы выяснены не до конца. Раздражители могут либо действовать на мембрану, либо включать в цитоплазме механизм сокращения. В настоящее время считается вполне установленным, что в основе цитоплазматического сокращения лежит регуляция концентрации кальция в цитоплазме амебы.
Амебы размножаются путем митотического деления ядра, за которым следует деление всей клетки. Перед митозом амеба прекращает движение и принимает округлый и бугристый вид, но после деления ядра приобретает снова обычную форму и восстанавливает амебоидное движение. Пока клетка временно является двухъядерной, псевдоподии вытягиваются в противоположных направлениях и она буквально разрывает себя на две одноядерные амебы.
7.9. Другие саркодовые отличаются от амеб структурой псевдоподий, жизненным циклом и способом защиты
Ближайшими родственниками амеб, не имеющих защитного слоя, являются виды, образующие раковину или же формирующие раковину или панцирь из твердых частиц, собранных из окружающей среды. Эти раковинные амебы (рис. 7-4, А) часто встречаются в пресной воде и легко определяются в ее пробах по характерным раковинам. Амебы, которые живут в раковинах, очень похожи на некоторые виды амеб, не имеющих защитного слоя. Более отдаленными родственниками этих групп являются фораминиферы, радиолярии и солнечники (рис. 7-4, Б, В, Г). Эти три класса саркодовых имеют тонкие нитевидные псевдоподии, но способ их движения и поведенческие реакции отличаются от амеб.
Рис. 7-4. Некоторые представители саркодовых. A. Difflugia - раковинная амеба. Б. Allogromia - фораминифера, не имеющая твердой раковины. 5. Radio phrys - колониальный солнечник. Г. Acanthometron - радиолярия. А и В - пресноводные; Б и Г - морские. (А, Б и Г - микрофото автора; B - по С. W. Wattets.)
7.10. Фотосинтезирующие и гетеротрофные жгутиковые типа Mastigophora обладают большим разнообразием способов питания, которые позволяют им существовать в различных условиях
Очевидно, жгутиковые являются преуспевающей группой одноклеточных. Большое разнообразие обнаруживают как Фотосинтезирующие ("растениеподоб-ные") фитофлагелляты, так и гетеротрофные ("животноподобные") зоофлагелляты. Некоторые фитофлагелляты в условиях отсутствия света могут осуществлять обмен веществ подобно животным клеткам и имеют мутантные формы без хлоропластов. Таким образом, отличия между фито- и зоофлагеллятами должны быть подчеркнуты особо. Действительно, оба типа, возможно, являются близкородственными.
Некоторые зоофлагелляты питаются бактериями, тогда как другие поглощают и расщепляют различные органические молекулы. Многие зоофлагелляты обитают в кишечнике высших животных в качестве симбионтов или паразитов. Разнообразные по структуре зоофлагелляты обнаружены в кишечнике различных видов термитов. Здесь они живут как симбионты, переваривая древесину для собственных нужд и обеспечивая питанием термита, в то время как термит, в свою очередь, обеспечивает приют, транспортировку и постоянную поставку древесины. В одном термите можно обнаружить более сотни различных видов зоофлагеллят.
7.11. Euglena является хорошим примером фитофлагеллят типа Euglenophyta
Хлоропласты фитофлагеллят, по-видимому, первоначально возникли из проглоченных сине-зеленых водорослей. Характерным представителем типа флагеллят является Euglena gracilis (рис. 7-5). Эвгленовые обычно обитают в пресной воде, богатой органическими питательными веществами и обладают способностью двигаться в направлении света. Эвг леновые представляют собой удлиненную клетку с хорошо сформированным ядром, содержащим ядрышко, двумя жгутиками (из которых выдвигается только один), митохондриями и сократительной вакуолью. Каждая клетка также содержит несколько хлоропластов и может осуществлять процесс фотосинтеза на солнечном свету. В темноте эвгленовые живут за счет органических веществ мертвых организмов.
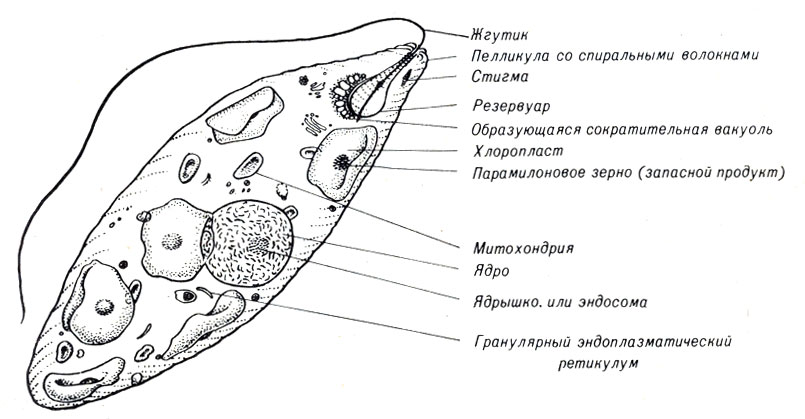
Рис. 7-5. Схема строения Euglena gracilis
Поскольку эвгленовые малы, то газообмен с окружающей средой происходит путем диффузии. В дневное время они утилизируют как углекислый газ, так и кислород. В теплое время суток у них осуществляется кислородное дыхание.
Подобно большинству пресноводных протист эвгленовые имеют сократительные вакуоли, которые удаляют из цитоплазмы избыток воды, входящей в клетку в результате диффузии.
Более длинный жгутик эвгленовых используется для движения. Кроме того, кортикальная область клетки под пелликулой (внешней оболочкой) содержит спирально извитой стержень сократительного материала, производящий перистальтические, так называемые эвгленовые, движения. Эти движения происходят, когда клетка подвергается освещению высокой интенсивности. Рядом с вакуолью Е. gracilis расположено красное пятно, называемое стигмой. Оно служит затеняющим устройством для фоторецептора и главным образом определяет поведение эвглены.
7.12. Организмы типа Ciliophora, по-видимому, произошли от примитивных жгутиковых предков и содержат различные специализированные структуры
Полагают, что представители типа Ciliophora произошли от примитивных жгутиковых предков. Обе группы обладают структурами, возникшими из центриолей, которые видоизменились в различных направлениях.
Латинские названия cilia (реснички) и flagella (жгутики) введены задолго до того, как было обнаружено, что их сходство по структуре и химическому составу, вероятно, значительно большее, чем некоторые различия в обеспечении способов движения. Однако, несмотря на сходство этих двух структур, реснитчатые и жгутиковые организмы определенно различаются во многих отношениях, что свидетельствует об их дивергенции на очень ранних этапах эволюции.
Представители класса Ciliata, или реснитчатые, составляют успешно развивающуюся группу одноклеточных, которые адаптировались к таким различным местам обитания, как открытые океаны, дренажные канавы и даже кишечник животных. Реснитчатые классифицируются по расположению ресничек на поверхности тела. Реснички содержат два вида ядер - микро- и макронуклеусы. Отдельные представители основных групп показаны на рисунке 7-6.
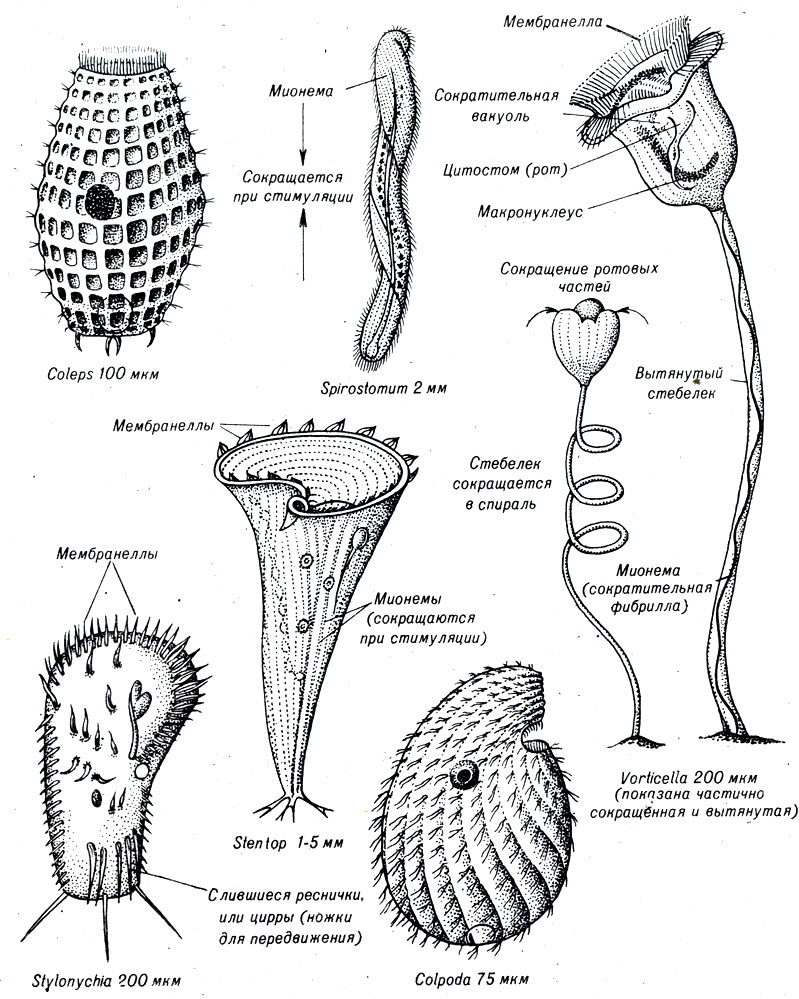
Рис. 7-6. Некоторые разнообразные формы свободноживущих реснитчатых
Многие реснитчатые размножаются бесполым путем - делением. У некоторых же наблюдается половой процесс в виде коньюгации, который включает обмен микронуклеусами между двумя организмами. В результате конъюгации генетический полиморфизм и вследствие этого увеличивается генетическое разнообразие реснитчатых. Многие реснитчатые обладают постоянными специализированными структурами, характерными для парамеций, которые будут рассмотрены в качестве примера в следующем разделе.
7.13. Paramecium является одним из наиболее изученных представителей реснитчатых, и в настоящее время многое известно о ее структуре, химическом составе, генетике и поведенческих реакциях
Частое использование Paramecium в качестве объекта исследований связано с тем, что по крайней мере один из нескольких ее видов обнаруживается почти повсеместно в пресной воде.
Paramecium multimicronucleatum (рис. 7-7) представляет собой животное в форме туфельки с постоянным околоротовым углублением. Ряды ресничек покрывают все тело, включая околоротовое углубление. Наиболее заметными внутренними структурами являются две сократительные вакуоли, несколько микронуклеусов и единственный макронуклеус. Кортикальный слой цитоплазмы содержит базальные тельца ресничек, заряженные трихоцисты и различные фибриллярные структуры. Трихоцисты - это органеллы, расположенные под клеточной поверхностью, которые выстреливают нитеподобные структуры, охраняющие животное от хищников или захватывающие жертву. Внутренняя цитоплазма содержит митохондрии, комплекс Гольджи и другие органеллы, обычные для эукариотических клеток.
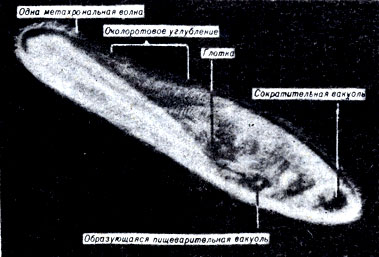
Рис. 7-7. Paramecium multimicronucleatum, сфотографированная при световой вспышке, чтобы остановить метахрональный характер биения ресничек. (Фото Н. Machemer.)
С помощью ресничек парамеция создает постоянный ток воды, направляющий пищевые частицы в околоротовое углубление, где они поступают в цитостом (клеточный рот) и проходят в пищеварительную вакуоль, окруженную мембраной. В то время как пища переваривается, пищеварительные вакуоли подхватываются циркулирующим потоком цитоплазмы, или циклозом, и перемещаются внутри клетки. Непереваренная часть пищи" остающаяся в вакуоли, выбрасывается. Азотистые продукты распада могут удаляться из клетки либо путем диффузии, либо вместе с водой через сократительную вакуоль. Сократительная вакуоль необходима пресноводным протистам для удаления избытка воды, диффундирующей в клетку. Газообмен у парамеции осуществляется путем диффузии.
Перемещаясь в воде, парамеции вращаются вокруг своей продольной оси. Индивидуальные реснички бьют по воде подобно тоненьким веслам, чередуя эффективный взмах с восстановительным. Реснички бьются метахрональными волнами, это значит, что в каждый данный момент времени реснички, расположенные вдоль тела клетки, находятся в нескольких различных фазах своего движения (рис. 7-7). Такая картина несколько напоминает вид пшеничного поля, охваченного порывами ветра.
Парамеции имеют тенденцию двигаться почти по прямой линии до тех пор, пока они не натолкнутся на препятствие. В этом случае они изменяют направление своего эффективного взмаха и гребут обратно, затем производят поворот на 25-30° и начинают снова двигаться вперед. Этот процесс называют реакцией уклонения. Недавно Эккерт (R. Eckert) и Найтох (Y. Naitoh) показали, что парамеции осуществляют реакцию уклонения путем изменения проницаемости плазматической мембраны, регулирующей поступление в клетку кальция. Если ионы кальция присутствуют в кортикальном слое, животное плывет назад; если же они удаляются, то животное возобновляет нормальный курс.
Парамеции обычно размножаются путем клеточного деления (см. рис. 1-1), но могут также обмениваться генетическим материалом при конъюгации. Для осуществления этого примитивного полового процесса два организма временно сливаются в области околоротового углубления и обмениваются микронуклеусами через временный мостик.
7.14. Тип Sporozoa состоит из паразитирующих форм со сложным жизненным циклом. Некоторые формы вызывают серьезные заболевания человека и животных
Тип Sporozoa получил свое название потому, что на одной из стадий своего сложного жизненного цикла эти паразиты существуют в виде спор, содержащих инфекционные спорозоиты. Представители этого типа одноклеточных привлекают большое внимание, поскольку являются возбудителями заболеваний у человека и животных. Жизненный цикл споровиков может включать бесполое и половое размножение и происходить со сменой двух хозяев.
Plasmodium является паразитом, вызывающим заболевание малярией. Хотя распространение малярии в настоящее время ограничено тропической зоной, полстолетия назад это заболевание было широко распространено в умеренных зонах Европы и Соединенных Штатов Америки. Заболевание переносится комаром Anopheles, в теле которого проходит половина жизненного цикла паразита. Другая половина его жизненного цикла протекает в теле человека. Схема жизненного цикла плазмодия представлена на рисунке 7-8.
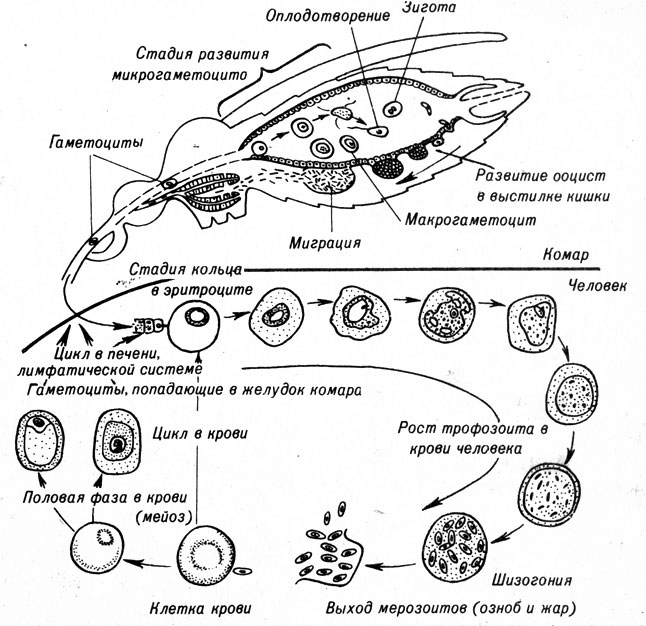
Рис. 7-8. Цикл развития малярийного паразита Plasmodium. Спорозоиты проникают в тело человека со слюной комара и попадают в печень и лимфатическую систему. Здесь они развиваются в мерозоиты, которые проникают в новые клетки печени или переходят в клетки крови. В крови мерозоиты размножаются и дают начало большему числу мерозоитов, которые вместе с токсическим веществом вызывают озноб и жар, характерные для малярии. Этот цикл может характеризоваться 24-часовой периодичностью. Некоторые мерозоиты претерпевают мейоз с образованием двух типов гаметоцитов. Гаметоцихы попадают в желудок комара и там сливаются. Зигота внедряется в стенку кишечника комара и превращается в ооцисту. В ооцисте путем митотического деления образуется большое количество спорозоитов, которые попадают в слюнную железу при разрушении ооцисты
Для установления контроля над заболеванием необходимо разработать меры борьбы с комаром Anopheles. В прошлом это осуществлялось с помощью инсектицида ДДТ, но этот препарат сейчас используется значительно реже из-за его токсичности и продолжительности действия на окружающую среду. Предпринимаются усилия для ограничения численности комара Anopheles и других опасных насекомых с помощью биологических мер борьбы, которые не наносят вреда окружающей среде, но эффективно тормозят размножение насекомых и тем самым снижают численность их популяции.
7.15. Protophyta и Protozoa ставят перед таксономистами крупную проблему: являются ли они протистами, растениями или грибами
При введении классификации из пяти царств было принято произвольное решение: отнести все одноклеточные эукариотические организмы к одному царству - Protista. Такое отделение одноклеточных Protista от многоклеточных Planta, Fungi и Animalia не всегда четко проявляется в природе, и это, соответственно, ставит проблемы перед систематиками. Не имеет значения, какие границы проведены между царствами, типами, классами и другими подразделениями. Они искусственны, созданы для удобства человека, и не все организмы будут точно подходить под установленные категории. Замечательным примером в этом отношении являются растениеподобные протесты. Эти организмы могут быть классифицированы по-разному среди ;царств Planta, Protista и Fungi. В данной книге типы Chrysophyta, Bacillariophyta, Xanthophyta, Pyrrophyta и одноклеточные Chlorophyta (зеленые водоросли) будут рассмотрены среди протает.
Обсуждение многоклеточных представителей Chlorophyta вместе с бурыми и красными водорослями отнесено к разделу растений, а слизевики (тип Мухо-mycota) будут рассмотрены вместе с грибами в главе 8.
7.16. Тип Pyrrophyta включает динофлагеллят, которые в больших количествах встречаются в фитопланктоне
Динофлагелляты вместе с диатомовыми и другими растительными протестами составляют фитопланктон, или траву моря. Большинство динофлагеллят имеет два жгутика одинаковой длины, один из которых расположен в неглубоком желобке, огибающем тело. Они секретируют целлюлозную оболочку часто неправильной, но видоспецифичной формы и обычно имеют панцирные щитки. Хлоропласты динофлагеллят содержат хлорофилл и другие пигменты, которые придают им красно-коричневый оттенок. Они содержат красное пятно, подобное стигме эвгленовых флагеллят.
Наиболее известные динофлагелляты Noctiluca и Gonyaulax являются причиной люминесценции моря. В местах интенсивного размножения Gonyaulax число этих организмов увеличивается иногда в 5-6 млн. раз, и тогда море приобретает красный цвет. В это время у берегов наблюдаются красные приливы и отливы. Токсины, секретируемые динофлагеллятами, убивают рыбу, которая разлагается и поглощает из воды так много кислорода, что гибнет еще большее количество рыбы. Благодаря гниению образуются питательные вещества, которые, в свою очередь, способствуют увеличению численности представителей рода Gonyaulax.
7.17.Типы Chrysophyta [золотистые водоросли], X anthophy ta [желтые водоросли] и Bacillariophyta [диатомовые] являются важными первичными продуцентами в водной среде
Типы Chrysophyta, Xanthophyta и Bacillariophyta включают десятки тысяч видов, большинство из которых являются одноклеточными. Известны также колониальные, нитевидные и многоклеточные формы. У этих организмов хло-ропласты (желтые, коричневые, золотисто-коричневые) содержат три различных типа хлорофилла и несколько вспомогательных пигментов. Они имеют мощные клеточные стенки и запасают питательные вещества в виде масла или лейкозина. Уникальность клеточных стенок диатомовых заключается в том, что они являются углеродными и пропитаны кремнеземом. При митотическом делении каждая дочерняя клетка получает только часть клеточной стенки и затем синтезирует остальную. Панцири диатомовых симметричны, богаты орнаментами и особенно красивы, когда содержат живую клетку (рис. 7-9). Морфология панциря является основой для определения видов. Диатомовые чрезвычайно многочисленны и отвечают за большую часть фотосинтеза, который осуществляется в океанах. Погибшие диатомовые опускаются на дно, где в течение тысячелетий их кремневидные скелеты накапливаются на больших глубинах на дне озер и океанов. В результате сжатия земной коры иногда обнажаются огромные отложения диатомового грунта. В наши дни диатомовые отложения добываются и используются во многих производственных процессах.
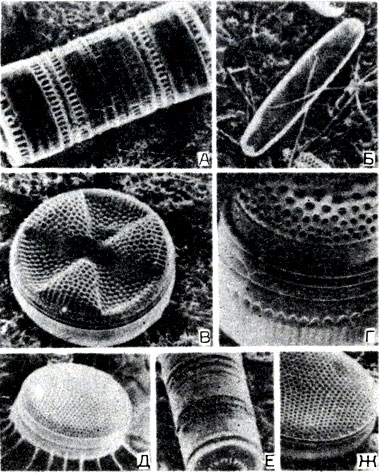
Рис. 7-9. Коллекция морских диатомовых, демонстрирующая значительное разнообразие формы и морфологии раковины. A. Paralia sulcata. Б. Navicula sp. В. Actinoptychus siharilus. Г. Thalassiosira. Д. Planktoniella. Е. Paralia sp. Ж. Coscinodiscus radiatus. Сфотографированы при помощи сканирующего электронного микроскопа. (Фото S. Honjo.)
7.18. Одноклеточная зеленая водоросль Chiamydomonas является представителем фитофлагеллят. Некоторые родственные ей многоклеточные формы занимали ключевые позиции в эволюции растений
Тип Chlorophyta (зеленые водоросли) будет рассмотрен также и в следующей главе, так как его представители относятся к двум царствам, если считать многоклеточность (или многоядерность) основной разграничения на протесты и более высокоорганизованные формы организмов.
Chiamydomonas является одним из наиболее простых и подробно изученных одноклеточных представителей типа Chlorophyta. Как и все представители этого типа, они имеют три характерные особенности: клеточные оболочки, со^ держащие целлюлозу, хлоропласт, имеющий хлорофилл, и крахмал в виде запасного питательного вещества. Chiamydomonas (рис. 7-10) широко распространена в пресных водоемах. При длине около 25 мкм она имеет плотную клеточную оболочку и два жгутика одинаковой величины. Хламидомонада содержит единственный хлоропласт в форме чаши, который частично охватывает ядро. Будучи обычно гаплоидной, она размножается бесполым путем - митотическим делением. Иногда дочерние клетки функционируют как изогаметы и сливаются (в начальной стадии путем жгутикового контакта), формируя диплоидную зиготу с четырьмя жгутиками. Затем зигота переходит в стадию покоя и перезимовывает. При -благоприятных условиях среды происходит мейоз, который восстанавливает нормальное гаплоидное состояние. Полагают, что этот простой половой цикл представляет собой самую раннюю форму полового размножения растений.
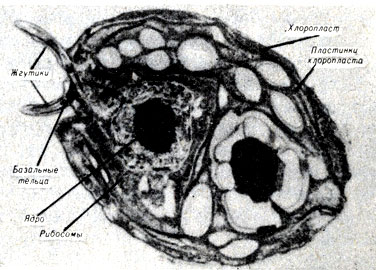
Рис. 7-10. Chiamydomonas reinhardtii, одноклеточная зеленая водоросль, сфотографирована при помощи трансмиссионного микроскопа, включает два жгутика, ядро и хлоропласты. (Фото D. Ringo.)
7.19. При изучении водорослей возникают проблемы их систематизации. Как должны быть классифицированы колониальные и нитчатые водоросли? И куда отнести слизевиков?
Среди зеленых водорослей существует особенно много форм, тесно связанных с одноклеточными колониальными и нитчатыми водорослями. У одних водорослей все клетки сходны по структуре и функции, у других одна или несколько клеток дифференцируются и образуют половые клетки или прикрепляющуюся часть таллома. В этом случае совокупность клеток в действительности не является больше колонией, а представляет собой очень простой многоклеточный организм. Вопрос о разделении на проти-сты и растения до сих пор остается спорным, однако границы между ними провести необходимо, хотя они будут искусственными.
Для установления наиболее естественных категорий исследователи пытаются реконструировать возможный путь эволюции. Таксономисты растений длительное время были заняты вопросом происхождения наземных растений и спорили о возможном эволюционном пути, который привел к их появлению. Эволюция, по-видимому, сопровождалась появлением огромного числа небольших наследственных изменений, только некоторые из них были подхвачены естественным отбором. Очень немногие формы живых ископаемых организмов еще сохранились, иллюстрируя путь эволюции. Перед таксономистами стоит важная проблема, является ли все, с чем они работают, листьями эволюционного дерева, а не стволом или ветвями, по которым можно было бы проследить взаимосвязь между листьями. Тем не менее в настоящее время считают, что наземные растения произошли от предков зеленых водорослей типа Chlorophyta. Более того, некоторые группы водорослей этого типа, по-видимому, находятся где-то близко к эволюционному пути, который привел к мохообразным и высшим наземным растениям. Таким образом, одноклеточных представителей типа Chlorophyta следует рассматривать как протист - предков высших растений.
Другой серьезной проблемой при классификации протист является разграничение между красными и зелеными водорослями, с одной стороны, и грибами - с другой. Теперь кажется вероятным, что слизевики являются промежуточной группой. Типы Myxomycota (истинные, неклеточные слизевики) и Acrasiomycota (клеточные слизевики) часть своего жизненного цикла существуют как почвенные амебы или как жгутиковые. Это позволяет отнести их к протистам. Однако слизевики в амебоидной стадии могут (но не обязательно) сливаться в многоядерную цитоплазматическую массу, или плазмодиум, который растет и мигрирует в поисках бактерий. В этой плазмодиальной стадии слизевики напоминают некоторые виды грибов. Они развиваются во влажном воздухе после дождя, разлагая поваленные деревья. Когда появляется солнце и лес высыхает, плазмодиум также начинает подсыхать и формирует тонкие плодоносящие тела, содержащие споры, что является характерным признаком грибов. В дальнейшем споры высыпаются и разносятся на значительные расстояния. При благоприятных условиях среды, наличии влажности и питательных веществ они переходят в следующую стадию жизненного цикла. Следовательно, слизевики являются промежуточными формами и могут рассматриваться и как протисты, и как грибы. Происхождение высших грибов в настоящее время неизвестно. Существует предположение, что различные группы грибов произошли от зеленых водорослей, красных водорослей или зоофлагеллят.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'