Глава 21. Биотопы и биоценозы в водоёме
Водные обитатели, которым посвящена наша книга, рассматривались нами порознь и лишь с некоторым освещением их отношений, к другим пресноводным животным и к факторам внешней среды. Между тем зависимость их от окружающего мира многосложна; она не ограничивается ближайшим окружением, то есть пределами самого водоёма, но стоит в связи и с более отдаленными косвенно действующими факторами, находящимися вне водоёма.
Совокупность условий, существующих внутри водоёма, внешних по отношению к нему факторов, связанных с ним по месту и времени й истории формирования водоёма, определяет лицо его фауны и флоры. Состав же растительного и животного населения в связи с его экологическими чертами обеспечивается в своей относительной устойчивости существованием биоценотических связей между ними. Так определяется жизнь биоценоза в биотопе, которым является водоём. Небольшие водоёмы представляют единый, однообразный по своему физическому характеру биотоп. Характер и величина водоёмов варьируют от временной лужи и скоплений воды в пазухах листьев, в дуплах деревьев и в углублениях скал до огромных озёр и целого бассейна большой реки. В больших водоёмах присутствует ряд биотопов различного характера, населенных своими биоценозами с их типичными сочленами, свойственными данному биотопу. При расчленении водной среды составляющие её биотопы не отделяются непреодолимыми преградами один от другого; при активной подвижности и случайных обстоятельствах обитатель одного биотопа может очутиться в другом месте водоёма, которому свойственен биоценоз иного состава. Такое обстоятельство не противоречит возможности и необходимости разграничивать биотопы водоёма и рассматривать совокупности его населения как группу биоценозов.
В водоёме различаются: ложе, в котором находится вода, включая сюда дно водоема и примыкающие к нему временно затопляемые или увлажняемые путем заплеска воды берега, и масса воды. В пределах последней на границе её с атмосферой дифференцируется плёнка поверхностного натяжения, по своим физическим свойствам отличная от прочей толщи воды. Зимой поверхность водной массы несет на себе твердый ледяной покров.
Рельеф и геологическое строение окружающей местности, размеры водоёма, конфигурация дна и берегов, характер питания за счет атмосферных осадков, подземных ключей или притока воды о более или менее обширного бассейна, химизм; воды, зависящий от химизма пород, слагающих ложе водоема и других причин, степень его освещенности, условия нагревания и охлаждения - всё это отражается на видовом составе организмов, их количественном развитии и распределении внутри водоёма, и ведет к сосредоточиванию различных видов, в разных участках водоема. Такое распределение является следствием эволюционно сложившихся адаптации организмов к обитанию в различных условиях среды, типич ных для разных частей водоёма. Напомним, что в таком именно смысле различают обитателей текучих и стоячих вод - реофилов и лимнофилов.
В пределах каждого отдельного водоёма принято различать следующие места жизни и населяющие их группы организмов.
А). Дно бассейна или бенталь, населенное донными или бентическими организмами, объединенными под общим термином бентос.
Некоторые из этих организмов свободно ползают по дну, как например, личинки веснянок и ручейников, брюхоногие моллюски, водяные ослики и некоторые водяные клещи; другие - частично или полностью закапываются в дно, как это делают двустворчатые моллюски, личинки некоторых стрекоз, поденок и тендипедид, некоторые ракушковые рачки, многие малощетинковые черви (например, Tubifex) и нематоды; третьи временно или постоянно прикрепляются или даже прирастают к субстрату - к водным, растениям и другим находящимся на дне предметам - таковы губки, мшанки, дрейссена, личинки тендипедид и ручейников, живущие в неподвижных домиках, гидры, куколки многих насекомых и яйцевые кладки разных водных животных; некоторые яйцевые кладки и куколки насекомых свободно лежат на дне.
Обильные скопления нитчаток, или высших водных растений, например роголистника или перистолистника, образуют как бы "второе дно", находящееся иногда на один или два метра над поверхностью истинного дна водоема; "вторым дном" определяется глубина погружения слабо плавающих животных, как например, личинок комаров, для которых большие глубины в некоторых случаях могли бы быть губительными.
Б). Толща воды населена организмами, составляющими несколько крупных экологических групп. Одной из них являются планктические организмы, в совокупности обозначаемые понятием планктон; это очень мелкие, по большей части микроскопические организмы, которые пассивно или слабо активно плавают в воде; у некоторых из них нет органов передвижения (например, у планктических диатомовых водорослей), у других органы плавания настолько слабы, что при малых размерах животных они не дают им возможности противостоять действию ветра и течений - таковы простейшие, коловратки, ветвистоусые и веслоногие рачки планктона. Более или менее крупные организмы, которые могут активно передвигаться в толще воды, составляют экологическую группу, называемую нектон - таковы рыбы.
Микроорганизмы, обитающие в поверхностной плёнке воды, в совокупности составляют группу, обозначаемую термином нейстон. Наконец, в виде особого сугубо сезонного образования выделяется пагон, то есть совокупность организмов, вмерзающих в лед (С. А. Зернов).
В более или менее обширных водоёмах, например, в озёрах, донная область (бенталь) и толща воды далеко неоднородны в разных своих участках, с которыми связаны видовой состав, биологические особенности и адаптивные признаки их населения. В бентали озера это будут:
- омываемое и затопляемое побережье,
- прибрежная зона собственно водоёма или литораль, доступная действию прибоя и зарастающая макрофитами,
- сублитораль, или переходная зона,
- профундаль, недоступная действию прибоя область иловых отложений.
В пределах толщи воды озера различают прибрежную область, подстилаемую литоральной и отчасти сублиторальной зонами дна, и открытую область, или пелагиаль, простирающуюся над профундалью и примыкающей к ней частью сублиторали.
В пределах литорали особенно часто встречаются организмы с различной экологией, которые то держатся на дне или на твердых предметах, находящихся на нем, то свободно плавают в воде, подобно планктическим организмам, то неподвижно сидят, укрепившись своей присоской (сида) или временно зарываются в грунт дна (коретра). Такие организмы составляют переходную группу планктоно-бентос.
Крупные водяные жуки (плавунец, ацилиус) или клопы (гладыш, плавт) то сидят неподвижно, уцепившись за растения, то быстро плавают в воде с помощью своих сильных гребных органов; они составляют группу нектоно-бентоса.
Население континентальных водоемов имеет различную филогенетическую давность связи своих компонентов с водной средой. Одни виды пресноводных животных являются первично водными организмами. Это те существа, предки которых и они сами в течение всей истории своего развития всегда жили в воде и поныне живут в ней; в пресную воду они пришли из солоноватых вод морских побережий; таковы рыбы, ракообразные, пластинчатожаберные и переднежаберные моллюски, черви, губки и простейшие. Эти организмы всю свою жизнь проводят в воде.
Другие виды пресноводных животных являются вторичноводными; ближайшие предки их были наземными обитателями, но в процессе последующего филогенетического развития они приспособились к обитанию в водной среде и частично или полностью порвали с прежней наземной или сухопутной средой.
Степень "привязанности" вторичноводных животных к водной среде различна. Особи многих видов почти всю свою жизнь проводят в воде, как например, лёгочные моллюски и некоторые виды водяных клопов и жуков, лишь по временам в период усыхания водоёма или для расселения покидая его. Другие виды вторичноводных остались настоящими амфибиотическими животными, которые в одной части своей жизни живут в воде, а в другой - её покидают, выходя на сушу или вылетая в воздух (таковы, например, двукрылые, бабочки, вислокрылки).
Как первичноводные, так и вторичноводные животные обладают различными особенностями морфологии, которые позволяют им жить в условиях того или иного биотопа; выявление адаптации, изучение и правильное понимание их значения является одной из интереснейших задач морфологии, физиологии, биологии и экологии.
При изучении степени приуроченности вида к существованию в тех или иных биотопах прежде всего бросается в глаза, что одни организмы могут жить в более или менее ограниченных условиях среды, вторые выдерживают значительные их колебания; первые организмы называются стенотопными и стенопластичными, вторые - политодными и еврипластичными.
Различные группы стенотопных организмов могут жить или только в чистой или в загрязненной воде, могут быть приуроченными к известным узким пределам солености, обитать в условиях постоянной температуры, притом или высокой, или низкой, жить в хорошо освещенных биотопах и в темноте. В пределах каждого из названных биотопов стенотопные организмы могут переносить лишь небольшой, терпимый для данного вида узкий диапазон колебаний факторов внешней среды, то есть они стенопластичны.
Политопные организмы встречаются в различных биотопах; они могут жить в условиях широкого диапазона колебаний факторов среды этих биотопов, они еврипластичны. Так например, еврипластичный прудовик Limnаеа truncatula выдерживает значительные колебания температуры и влажности.
При изучении условий обитания организмов различают оптимум факторов среды, например оптимальную температуру, оптимальный химизм воды, оптимальную освещенность, при которой жизненные функции данного организма протекают наиболее благоприятно для него. Колебание в сторону максимального или минимального действия фактора видоизменяет их то в сторону чрезмерного, вредного для организма ускорения, то, наоборот, понижает их; для стенопластичных организмов характерен узкий диапазон уклонений факторов от их оптимума в сторону переносимых видом максимальных или минимальных их значений, для еврипластичных - широкий; разница показателей максимальных, минимальных и оптимальных значений факторов среды, доступной для жизни этих организмов, велика. Необходимо заметить, что одни и те же виды могут проявлять различное отношение к разным факторам среды: например, быть еврипластичными по отношению к температуре и стенопластичными по отношению к. содержанию в воде органических веществ и кислорода или другому компоненту её химического состава.
Рассмотрим отдельные факторы среды и отношение к ним водных организмов.
Химический состав природных вод зависит от многих причин, в основе которых лежит состав пород, образующих территорию бассейна водоёма, и в частности его берега и ложе. Известно, что дестиллированная вода является ядом для многих живых организмов, которые в ней существовать не могут. В природе самая чистая пресная вода имеет ничтожную примесь различных солей, делающих её пригодной для жизни в ней организмов. Соли вода получает благодаря выщелачиванию пород, слагающих бассейн и чашу водоёма; в зависимости от особенностей пород бассейна вода может принимать иногда весьма своеобразный состав, например, приобретать соленость различной концентрации. Соответственно этому в процессе эволюционной адаптации организмов к столь специфичным условиям водной среды меняется и состав биоценозов. Имеются виды животных, которые могут существовать и в солёной и в пресной воде; таковы эвригалинные организмы, как например угри, лососи, сельди и некоторые инфузории (Condylostoma patens) и планарии (Macrostoma hystrix). Иными качествами обладают стеногалинные организмы, приспособившиеся к жизни в воде определенной степени солёности; галобионтами являются личинки мух Ephydra riparia; их пупарии широкой полосой покрывают прибрежную часть Тамбуканского озера, грязь которого используется для лечения; из других галобионтов следует упомянуть общеизвестного рачка артемию (Artemia salina), а из простейших красно пигментированного биченосца Dunaliella salina. Специфические особенности химического состава водоёмов могут до крайности обеднять их животное население или делать их вовсе безжизненными. Например, соленое озеро Эльтон летом при солености воды 200‰ заселено множеством особей всего лишь двух видов галобионтов - Dunaliella salina и Asteromonas gracilis (Plagellata): фауна других соленых озер с меньшей степенью солёности более обильна (до 72 видов).
С другой стороны, имеются специфически пресноводные животные, не выносящие поднятия солености выше 2‰; таковы моллюски из родов Unio и Anodonta и рачек Diaptomus gracilis.
Возможность обитания животного в воде, по химическому составу для жизни не пригодной, зависит от особенностей, образующихся в конкретных случаях микробиотопов водоёма. Например, сернистые воды непригодны для жизни личинок анофелес. Однако личинки A. maculipennis могут встречаться и в сернистых водоёмах, например, при условии, если в последних поселяются зеленые водоросли, в частности водяная сеточка (Hydrodictyon); в участках воды, заселённых её сетчатыми колониями и скоплениями нитчаток Spirogyra, идет обильное выделение кислорода, и сероводород исчезает; эти-то микробиотопы и заселяются личинками малярийных комаров.
Поучительно рассмотреть частный случай водоема с совершенно специфической химической средой. Дело идет о так называемых дупловых водоемах: (рис. 8). Они образуются то в основании ствола дерева, то на его боку; объём их обычно невелик - с вместимостью 2-3 литра. Вода в дупле скопляется вследствие таяния снега или выпадения дождя. Ложем водоёма является гниющая древесина. В такой водоем попадают упавшие листья и обломки веток, он часто засоряется землей и над придонной гущей в нем остается невысокий слой воды. Самое замечательное в дупловом водоеме - это изменение химического состава и цвета воды. В "старых" дупловых водоемах ясеня вода приобретает цвет темного портвейна или становится почти совсем черной. По химическому составу она отличается исключительно высоким количеством органических веществ, в образце воды дуплового водоёма ясеня близ Пятигорска достигающим 8303 мг на литр (Э. Карстенс, 1927). Количество неорганических веществ также высоко (269 мг на литр). Приведенный пример показывает, насколько резко изменился химический состав воды дуплового водоёма от начального состава дождевой воды, попавшей в дупло, до состояния, рассмотренного выше. Такая перемена зависит от экстрагирования древесины водой и от последующего усыхания его, приводящего дупловой водоём к высокой концентрации химических ингредиентов, а затем к полной ликвидации.

Рис. 8. Водоём в дупле ореха (фот. Н. Г. Брегетовой)
Замечательно, что и такой совершенно исключительный по своим особенностям водоём заселен своеобразным биоценозом, в состав которого входят стенотонные организмы и немногие евритопные виды, выдерживающие столь высокие концентрации органических веществ, в частности гуминовых (В. Рылов). В дупловых водоёмах обитают личинки черноногого малярийного комара Anopheles plumbeus и комара Finlaya geniculata, личинки мокрецов, личинки мухи Myiatropa (близкие к "крыскам" Eristcuis), реже личинки других мух. Все они здесь же окукляются и дают вылет взрослого насекомого, если дупловый водоём раньше того не пересохнет. Личинки Anopheles plumbeus, как и Finlaya geniculata, при высыхании дуплового водоёма уходят в ил на дне и могут жить еще некоторое время в жидкой грязи. Из микроскопических организмов в дупловых водоемах известны 6-7 видов жгутиковых, лишенных хлорофилла, три вида инфузорий и цисты простейших (Е. Павловский и А. Месс). Вследствие специфических особенностей подобных водоёмов, население их качественно бедное.
Другой физико-химической особенностью, не менее важной для населения водоёма, является качественный и количественный со став газов, растворенных в воде.
Вода получает газы из разных источников - внешнего и внутренних. Внешним поставщиком газов является атмосфера, о которой соприкасается поверхность водоема ("дыхание" самого водоёма); внутренними факторами служат процессы питания и дыхания погруженных зеленых растений, жаберного и кожного дыхания животных, гниения белков и разложения растительной клетчатки.
При "дыхании" самого водоёма, его водная поверхность абсорбирует газы, являющиеся составными частями атмосферы в различной степени. Благодаря особенностям коэфициента абсорбции для газов атмосферы пропорция растворения кислорода и азота в воде оказывается вдвое большей для кислорода, нежели в смеси этих газов в атмосфере (в воде отношение кислорода к азоту около 1:2, а в воздухе 1:4).
Внутренним источником получения водою кислорода является выделение сто в процессе фотосинтеза зеленых растений на свету, особенно во время "цветения воды", приводящего иногда к перенасыщению воды кислородом.
Потребителями кислорода в воде являются животные, имеющие водный тип дыхания, то есть дышащие за счет растворенного в воде кислорода (жаберное и кожное дыхание). Кислород поглощают бесхлорофилы - растительные организмы и зеленые растения в темноте. Кроме того, немало кислорода затрачивается на процессы гниения остатков погибших животных и растений. Углекислота поступает в толщу воды при дыхании водных животных и бесхлорофильных растений и при разложении органических веществ; её потребителями являются зеленые растения в процессе фотосинтеза.
Газами, придающими воде специфические особенности, являются метан (болотный газ), образующийся в водоёме при разложении клетчатки, и сероводород, выделяющийся от гниения органических веществ на дне и в толще воды.
Количество растворенного в воде кислорода различно в разных водоемах, и оно варьирует в одном и том же водоёме, особенно на разных глубинах (слоистость или стратификация распределения кислорода). Различные организмы по-разному реагируют на количество кислорода, растворенного в воде. Одни виды, как циклоп (Cyclops strenuus), могут жить при больших колебаниях кислорода в воде (эвриоксибионты); другие же (например, личинки поденки Ecdyonurus и Iron) требуют, гораздо более постоянного количества его в воде (стенооксибионты).
Условия дыхания водных организмов зависят от сочетания таких условий, как: объём воды в водоёме, условия её аэрации, характер и обилие животного и растительного населения, количественные соотношения между ними и объёмом воды, влияние температурного фактора, степень усыхания водоёма и сгущения его населения во времени года. Особо неблагоприятные условия могут создаться суровой зимой при значительной толщине ледяного покрова. Остающиеся в бодрствующем состоянии рыбы могут потребить весь запас растворенного в воде кислорода и погибнуть от удушья (так называемый замор рыбы). Недостаток в воде кислорода в таких случаях можно косвенна наблюдать через проруби, под которыми скопляются рыбы в поисках возможного облегчения дыхания. Замор рыбы бывает и от других причин. "Цветение" воды летом вызывается массовым развитием в ней сине-зеленых водорослей; отмирая, водоросли разлагаются и продуктами своего гниения отравляют воду, что может вызвать массовую гибель рыбы в рыборазводных прудах.
Для некоторых вторично водных животных характерно дыхание за счет атмосферного воздуха через посредство открытых наружу органов дыхания наземного типа, - трахеи паукообразных и насекомых и легкие моллюсков и земноводных. В этих случаях необходим контакт животного с воздушной атмосферой при поднятии животного к самой поверхности воды.
Животному, поднявшемуся на поверхность водоёма или прикоснувшемуся к ней удлиненными дыхательными придатками (сифонами), необходимо преодолеть силу поверхностного натяжения воды; это достигается механически благодаря несмачиваемости хитиновых покровов соответственного места тела насекомого; вода сама сбегает с окружности дыхательного отверстия, благодаря чему устанавливается непосредственное сообщение дыхательной системы с атмосферным воздухом. У других водных обитателей эволюционно выработались приспособления для соприкосновения с атмосферой на известной дистанции; такое значение имеет удлиненный придаток на заднем конце тела у водяного скорпиона, способность "крыски" (личинка, мухи Erystalls tenax) выдвигать дыхательный сифон сантиметров на 15, что позволяет ей, оставаясь на дне, пользоваться для дыхания атмосферным воздухом.
Многие животные адаптивно приобрели способность более или менее долго находиться под водой; однако эта способность их контролируется температурой среды.
Хотя личинки многих водных насекомых, например личинки комара анофелес, являются воздуходышащими, тем не менее, они могут потреблять и кислород, растворенный в воде. Фактором, определяющим такую возможность, служит температура. При более высокой температуре воды (24°-27°) личинки 1-й и 2-й стадии живут в воде, насыщенной кислородом, несколько суток, против всего лишь нескольких часов переживания их в воде с малым количеством растворённого кислорода. Общее понижение температуры до 10°С и ниже смягчает это различие, благодаря понижению у личинок газообмена. В рассматриваемых случаях речь идет о жаберном (анальные жабры) (Надо помнить, что анальные жабры исполняют также осморегуляторную функцию) и частично кожном дыхании личинок анофелес, являющимся дополнением к стигмальному дыханию атмосферным воздухом.
Важным условием, влияющим на характер населения водоёма, является его температурный режим, определяемый временами года. Особое значение имеет зимнее замерзание воды, которое распространяется на некоторую толщину её поверхностного слоя; при малой глубине водоёма он может промерзать до самого дна. Естественно, что и все обитатели такого водоёма вмерзают в сплошную массу образовавшегося льда. С. А. 3ернов с сотрудниками установил большое разнообразие водных организмов, которые могут, таким образом, благополучно пережить зиму.
Для обозначения всей совокупности водного населения, вмерзающего в лед, им был предложен термин "пагон". В состав его компонентов входят под Москвой 7 видов растений, 34 вида простейших, 4 вида червей, 30 видов коловраток, 5 видов моллюсков, 7 ракообразных, 26 насекомых и 4 вида паукообразных. Одни из этих организмов остаются в пузырьках воды в толще льда, другие действительно вмерзают в него. Следует иметь в виду, что лед, вмещающий в себя пагон, не "просто" кусок льда; если обломок льдины с вмерзшими в него организмами взять из водоёма и выставить на мороз, то, в зависимости от силы последнего, весь пагон может погибнуть; но в естественных отношениях при таком же неблагоприятном условии пагон может благополучно пережить и сильные морозы. Это различие объясняется тем, что в естественном своем окружении толща льда с пагоном со стороны атмосферы прикрыта снеговым одеялом; такое же значение имеет и вода, остающаяся подо льдом и сохраняющая более высокую, чем лед, температуру. Амплитуда температуры соответственно понижается от наибольших колебаний воздуха, до гораздо меньших колебаний в толще снега над льдом и еще меньших в толще самого льда. 3ернов в качестве примера приводит следующую шкалу падения амплитуды зимней температуры: -30,2° в воздухе, -6° под снегом на льду и не более -1° в толще льда с пагоном при неизменной температуре +0,6°С воды: подо льдом.

Рис. 9. Биотопы личинок малярийного комара Anopheles superpictus (фот. Е. Н. Павловского)
Диаметрально противоположным фактором является повышение температуры воды в водоёме сверх обычного предела; например, в мелких остаточных водоёмах при летнем спаде уровня горных речек в Средней Азии вода прогревается до 32°-34°, то есть далеко за пределы максимума, обычно наблюдаемого в основном водоёме; это обстоятельство оказывает большое влияние на ускорение жизненного цикла обитателей такого остаточного водоема и в первую очередь на выплаживание комаров (рис. 9). По отношению к Anopheles superpictus и другим малярийным комарам это обстоятельство имеет существенное значение, так как включает в систему противомалярийной борьбы необходимость уничтожения или обезвреживания таких водоёмов, что является кропотливой работой по учету их, определению локального эпидемиологического значения и по самой технике) их уничтожения.
Еще большее значение приобретает температурный фактор в заводях рек (рис. 10) и в рисовых полях, чеки которых сплошь заливаются водой на небольшую высоту (рис. 28). Прогреваемость воды в таких условиях бывает очень высокой; максимальная температура воды рисовых полей может достигать 38°-39° и даже 42° C; наибольший нагрев приходится на время с 11-12 часов дня до 4-5 часов вечера.

Рис. 10. Биотопы личинок Anopheles superpictus в заводях рукава Пянджа (фот. Е. Н. Павловского)
Влияние температуры на водное население является прямым и косвенным; при повышенной температуре идет обильное развитие зоо- и фитопланктона; его биомасса увеличивается, что отражается положительно на непосредственных потребителях этого сорта пищи, в частности на личинках малярийных комаров. Поэтому понятно, что рисовые поля законно считаются рассадниками малярии; для обезвреживания их в этом отношении применяются специальные методы борьбы, которые не должны снижать их сельскохозяйственной продуктивности. Положительное значение имеет проведение прерывистого орошения рисовых полей; не было недостатка и в административных постановлениях о запрещении устройства рисовых полей в ближайшем соседстве с селениями. Впрочем, это мероприятие под влиянием экономических затруднений не всегда проводилось в жизнь.
Под влиянием годовых и широтных изменений температуры может изменяться хабитус (внешность) и самый жизненный цикл организмов.
Некоторым пресноводным организмам, начиная с простейших (Flagellata) и кончая низшими ракообразными, свойственно явление цикломорфоза или сезонной изменчивости. Суть явления заключается в том, что взрослые особи одного и того же вида в разные времена года обладают различной внешностью (например, зимние и летние формы дафний, рис. 11), обитающие в ряду нисходящих поколений в одном и том же водоёме. Так как разные водоёмы в одном и том же сезоне могут иметь значительные температурные различия (например, холодноводные и тепловодные озера), то естественно ожидать в них видоизменения цикломорфоза тех же дафний; цикломорфоз имеет место при температуре, начиная с 12°-16° С. В водоёмах с более низкой летней температурой его не бывает; поэтому рассматриваемое явление не свойственно озерам Арктики и субарктики, равно как и высокогорья.
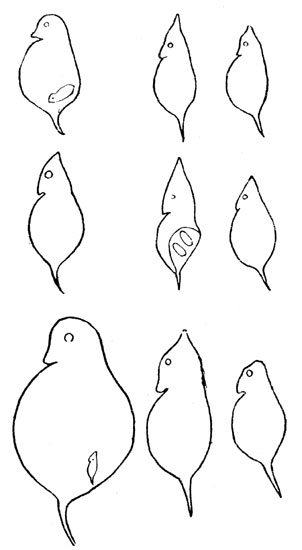
Рис. 11. Цикломорфоз дафний (Daphnia cucullata) в большом глубоком озере (верхний ряд), в мелком озерке (средний ряд) и в маленьком пруду (нижний ряд) (по Везенберг-Лунду)
С влиянием температурного фактора связывается и цикличность размножения, характеризующегося у дафний гетерогонией, то есть чередованием поколений, размножающихся то обоеполо, то партеногенетически; количество таких поколений у одного и того же вида ветвистоусых изменяется в водоемах разных типов, лежащих в разных местах, температурный режим которых неодинаков. Как общую зависимость, можно указать на широтное и высотное расположение рассматриваемого водоема. В связи с этим у некоторых из дафний на севере вовсе выпадает партеногенетическое поколение, тогда как на юге количество их в известных пределах возрастает. Но и в одних и тех же широтах тип размножения одного и того же вида Cladocera может быть различным, в зависимости от особенностей водоемов, в которых они обитают.
Так, например, Simocephalus в хороню прогреваемых мелких водовместилищах проходит дицикличный тип размножения, тогда как в больших озерах размножение идет моноциклически.
Возвращаясь к температуре водоёмов, необходимо отметить существование источников, температура которых не зависит от климата и сезона года, но связана с локальными геологическими условиями. Таковы тёплые и горячие ключи и источники (термы); предельная температура их весьма разнообразна, достигая в некоторых из них почти до 100° C. Важно определить верхнюю температурную границу, при которой возможна жизнь животных организмов в тёплых источниках. Такой температурой оказывается 40°-42° C, в более редких случаях 46°-52°. Понятно, что различные виды организмов по разному стойки к повышению температуры. Так, моллюск Bithynia thermalis в Италии живет в водоёмах с температурой до 53° С, некоторые водолюбы выдерживают температуру до 45° C. Обитание обыкновенных видов в условиях повышенной температуры может привести к изменениям частей их тела; так раковина Limnаеа при таких условиях развивается хуже, чем при обычной температуре. Режим жизни в тёплых водоёмах отличается большим однообразием, хотя бы потому, что нет зимнего замерзания водоёма.
Температурный фактор может привести к высыханию водоема. Это касается водовместилищ малой величины, лишенных естественного питания, и в первую очередь эфемерных водоёмов. Однако и они могут стать местом обитания некоторых специализованных организмов; напомним, что щитни могут быть обнаружены даже во временных дождевых лужах; весенние эфемерные водоёмы могут служить местом выплода ранних комаров. Судьба обитателей пересыхающего водоёма различна: одни из них беспомощны и быстро погибают; другие выработали приспособления к этому крайне неблагоприятному явлению и приобрели известную степень "засухоустойчивости". Особой формой такой адаптация является откладывание самками некоторых видов комаров Aedes яиц осенью на края временных водоёмов, которые неминуемо ежегодно высыхают. Яйца этих комаров зимуют в сухом субстрате. Весной, когда впадины высохших; водовместилищ заполняются талой водой, яйца дают выплод личинок ранневесенних поколений Aedes. Зимовка яиц входит, следовательно, в режим существования Aedes.
Степень освещения водоёма оказывает также влияние на жизнь его населения. Исконное отсутствие света в подземных пещерах наложило мощный отпечаток на эволюцию обитателей постоянных подземных озёр и речек. Так возникла пещерная фауна, водные представители которой лишены пигментации покровов и имеют редуцированные уменьшенные органы зрения или вовсе лишены их.
Открытое положение водоёма по отношению к дневному освещению или большая или меньшая степень его затенения растительностью или горами имеет значение для различных сочленов животного населения водоёма. Так, личинки обыкновенного малярийного комара обычно развиваются в хорошо освещаемых водоемах; на этом основании одной из мер борьбы с ними рекомендуется искусственное затенение хозяйственных водоёмов; практически это достигается обсаживанием берегов водоёма деревьями. Но на юге - в Закавказье, в Средней Азии и в Нижнем Поволжье в разгар лета личинки обыкновенного малярийного комара встречаются в затененных местах; в обычно заселяемых ими открытых водоёмах личинок уже не найти (Казанцев, Беклемишев, Половодова).
В вопросе о влиянии света на личинок анофелес необходимо различать несколько сторон дела: полная темнота делает их почти совсем неподвижными; резкое усиление освещения действует на личинок возбуждающе, что проявляется учащением произвольных перебежек личинок с места на место (явление фотокинеза). Однако реакция на освещение личинок анофелес, характеризующихся положительным фототаксисом, стоит в зависимости от величины водоёма, В обычных крупных водовместилищах реакция личинок на освещение нормальна, тогда как в остаточных микроводоёмах, в галечнике всего в несколько сантиметров величины, личинки держатся часто в тени от мелких галек.
Установившееся распределение личинок анофелес в случаях, которые как будто противоречат общей закономерности их поведения, находит себе удовлетворительное объяснение при более точном анализе действующих причин. Например, если личинок анофелес выставить в наполовину затемнённом плоском сосуде на яркий свет, то через некоторое время все личинки переместятся в тень. Противоречит ли такой результат опыта установившемуся представлению о положительном фототаксисе личинок? Не противоречит, потому что указанный эффект достигается лишь в небольших водовместилищах: при частых перебежках личинок на свету часть их время от времени попадает в тень; затенение же уменьшает подвижность личинок, и они не получают стимула к выходу из затененной части. В конце концов, все личинки могут очутиться в тени; это является не проявлением отрицательного фототаксиса, но "следствием фотокинетического возбуждения" (Беклемишев) в конкретной обстановке места и времени.
Более подробное рассмотрение влияния света на личинок анофелес нами делается в методологических целях; на таком удобном для экспериментирования и наблюдения объекте легко убедиться в том, что организм, подпадающий под влияние различных факторов; среды, может реагировать на основное раздражение различно, смотря по конкретным особенностям наблюдаемого случая.
Источником пищи для самых мелких из рассмотренных выше водных животных служит органический детрит и микроорганизмы планктона, нейстона и бентоса, в том числе бактерии, микроскопические водоросли и простейшие; растительную пищу более крупных животных составляют хлопья нитчаток и высшие водные растения, и животную - более мелкие обитатели водоёма и примыкающей к нему воздушной среды.
Особый интерес представляет возможность питания водных животных за счет самых малых микроскопических частиц. Так, например, личинки анофелес 1-й стадии могут отфильтровывать частицы размерами 22,8-34,2 микрона, а 4-й стадии - 68-165,3 микрона (Шипицына). Личинка 1-й стадии профильтровывает за сутки объём воды в четыре раза меньший, чем личинка 4-й стадии; но объём тела первой в несколько десятков раз менее чем у последней; поэтому личинка 1-й стадии получает относительно больше пищевых веществ (Сениор Уайт). В воде, имеющей лишь мелко дисперсные органические вещества, личинка 1-й стадии может продолжать метаморфоз до 3-й стадии, в которой и погибает. Личинки же 4-й стадии в таких же условиях не растут и гибнут от голода в более короткий срок.
Вследствие сложности и разнообразия источников питания сочлены биоценоза, населяющие биотоп, связаны друг с другом пищевыми (алиментарными) связями, которые в своей последовательности образуют пищевые ряды, или пищевые цепи. Схематически пищевой ряд или цепь можно представить в следующем виде: организм А поедается организмом Б, который в свою очередь . служит пищей организму В; этот последний становится жертвой организма Г и так далее. Если буквенные обозначения заменить названиями организмов, то может обнаружиться зависимость между весьма отдаленными существами, начиная, например, с бактерий и кончая рыбой. Разнообразие и обилие пищевых связей усугубляется еще и тем, что некоторые животные извне вторгаются в биотоп биоценоза и выхватывают для питания тот или иной организм; так например, чайка на лету захватывает плавающую в воде рыбу. Наземные млекопитающие также могут использовать водных животных в качестве пищи. Бурый медведь в некоторых местах истребляет много рыбы; например, на Камчатке во время хода лососевых, которые идут вверх по течению рек сплошной массой к местам нереста, медведи выбирают с берега любых рыб. Лисы пользуются рыбой не только из естественных водоемов, но и из прудовых хозяйств.
У рыб немало врагов и среди водных обитателей; особенно подвергается истреблению выметанная икра на разных стадиях эмбрионального развития, мальки и молодь. Претендентов на такую пищу много, особенно со стороны некоторых насекомых. Особую опасность представляют личинки и имаго плавунцов (Dytiscus), которые могут при подходящих условиях нацело выедать мальков сигов и карпов в выростных прудах. Показателем прожорливости этих насекомых являются экспериментальные наблюдения: взрослый плавунец окаймленный (D. marginalis) съедает в день 4,5 малька, а личинка этого вида - 7,6 мальков; молодая личинка поедает 5,5 мальков. Жукам не уступают и водяные клопы: взрослый гладыш довольствуется пятью мальками, а личинка нуждается в семи. И другие виды водных клопов являются врагами рыб, нападая преимущественно на молодь (водяной скорпион) или на небольших рыб (гладыш). Несоответствие в размерах и в силе гладыша и его жертвы покрывается токсическим действием слюны клопа, которая, впрыскивается в тело пойманной рыбы и парализует её.
Такие крупные хищники, как личинки стрекозы-коромысла справляются с мальками рыб с помощью своего жевательного ротового аппарата.
Из земноводных зеленые лягушки (Rana ridibunda) могут поедать множество мальков промысловых рыб. Водяной уж Natrix tesselata питается в полосе умеренного климата почти только рыбой; поедают её и пресноводные черепахи Emys orbicularis и др.
Рыбы имеют врагов и среди самих рыб. Общеизвестными хищниками являются щуки, а также сомы, излюбленной пищей которых служат, впрочем, лягушки.
Интересны случаи, когда рыбы стремятся использовать для питания животных - обитателей воздушной среды. Охота за подобной добычей достигается двояко - пассивными и активными действиями рыб. Многие сухопутные насекомые падают в воду с берега; их поедают рыбы без труда. В других случаях рыбы стремятся схватить насекомых в период их массового вылета или во время очень близкого нахождения их у поверхности водоёма; так форель прыгает за кладущей яйца веснянкой или подёнкой и ловит вылетающих комаров-дергунов.
Пищевое значение для рыб личинок водных насекомых очень велико; многие из них служат главным кормом для различных видов рыб; при этом используются личинки, держащиеся у поверхности воды (комары) и личинки, живущие на дне на большей или меньшей глубине или у берега водоёма (ручейники, тендипедиды, веснянки, подёнки, вислокрылки).
У рыб наблюдается и каннибализм, то есть пожирание особей своего же вида и даже собственного потомства. Личинкоядная бразильская рыба гамбузия, легко приживающаяся в водоёмах теплой и прилежащей к ней умеренной части палеарктики, живородяща; но если в аквариуме не создать условий для защиты отрожденной молоди, то выметавшая ее особь пожрет все свое потомство. Явление каннибализма у рыб наблюдается и в естественных условиях.
У амфибиотических животных при перемене места обитания меняется и характер питания; как источники пищи, так и строение и физиологические особенности одного и того же вида резко разнятся в фазах его водной и наземной жизни. Так например, личинка поденки в воде - растительноядна, имаго в воздухе вообще ничем не питается. Личинка анофелес в воде питается нейстоном и мельчайшим детритом; взрослый комар в воздухе сосет растительные соки (самец) или кровь животных (самка).
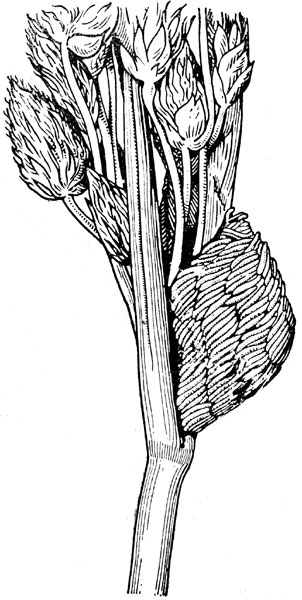
Рис. 12. Кладка яиц слепня (Tabanus autumnalis) на стебле (по Олсуфьеву)
Контакт амфибиотических животных с водоемом в период их наземной (воздушной) жизни имеет особое значение в период их размножения, так как развитие их должно итти в воде. В связи с этим различные виды амфибиотических животных поступают разно.
Самки слепней (Tabanidae) откладывают кучку яиц на поверхность листьев, свисающих над водоемом (рис. 12 и 13). Вылупляющиеся личинки, падая вниз, неминуемо попадают в воду, где и проходят водный период жизни. Другие насекомые выбрасывают яйца во время полёта над водой (подёнка Polymitarcys virgo, ручейник Hydropsyche ornatula). Комары откладывают яйца яа самую поверхность водоёма поодиночке (Anopheles, Aedes) или склеивают их в виде лодочки (Culex); стрекозы Erythromma najas целиком погружаются в воду и откладывают яйца в стебли водных растений.

Рис. 13. Места выплаживания слепней в речке Сиверская (Ленобласть) (по Олсуфьеву)
Контакт насекомых с водоёмом может вызываться потребностью их в воде для питья. Так слепни (Tabanus) в жаркие дни летят к лужам и другим вместилищам воды, с размаху бросаются на поверхность воды и тотчас поднимаются в воздух, унося на нижней стороне тела большую каплю воды, которую и всасывают через хоботок. На этой биологической особенности слепней рода Tabanus основан метод их уничтожения. И. Порчинский заливал некоторые лужи или искусственно сооруженные водовместилища нефтью. Пленка её, покрывающая воду, не отталкивала слепней от таких водоёмов, но при соприкосновении с залитой нефтью водой слепни обволакиваются нефтью и уже не могут оторваться от поверхности занефтеванной воды, остаются на ней и погибают. Н. Олсуфьев (1935) выловил во второй половине лета в Ленинградской области на трех замкнутых водоёмах с водным зеркалом в общей сумме 8 м2, покрытым тонким слоем нефти, за 5 дней 1769 экземпляров слепней, причем улов их колебался от 102 до 581 экземпляров за день. Таким путем вылавливаются и самцы, которые реже попадаются коллектору, очевидно, потому что летают выше, чем обычно самки. Следует принять в расчет что такие "ловчие лужи", или "лужи смерти", действуют в жаркую и более сухую часть лета, когда же в природе достаточно влаги в воздухе, почва также влажна, слепни не нуждаются в посещении водоёмов, и ловчий эффект луж снижается или вовсе выпадает. Рассмотренную потребность прямого контакта с водой проявляют все слепни, кроме дождевок (Chryzosona).
Для понимания всей полноты жизни водоёма необходимо рассмотреть условия существования и возникновения самого водоёма, стабильность его, пределы обмеления или временного высыхания, степень замкнутости или проточности, периодичность возникновения; необходимо также изучение биоценозов в водоёме и истории их возникновения в связи с окружающей водоём наземной природой и в зависимости от влияния деятельности человека. Из этой сжатой характеристики явствует сложность проблемы, в которой ложно разобраться лишь рассматривая вопрос по частям.
Остановимся на ознакомлении с факторами, обусловливающими существование сложившегося в водоёме биоценоза. Ввиду чрезвычайного разнообразия водоёмов и населяющих их биоценозов вопрос ставится здесь в общей, синтетической форме, применительно к водоёму замкнутого характера. Его население состоит из бактерий, водорослей, высших водных растений, простейших, кишечнополостных (гидры), плоских червей (ресничные черви или планарии), кольчатых червей, мшанок, низших и высших ракообразных, водяных пауков и клещей, личинок, нимф и имаго насекомых с неполным и с полным превращением, из низших насекомых (Apterygota), моллюсков, рыб и земноводных хвостатых и бесхвостых. Ближайшее отношение к населению водоёма имеют водоплавающие птицы.
Биоценозу водоёма свойственен определенный видовой состав (структура биоценоза), разнообразие и обилие которого чрезвычайно вариируют, смотря по характеру водоёма и истории его заселения. Биоценоз, или сообщество водных обитателей водоёма или его определенной части, имеет свою историю. Последняя уходит в давнопрошедшее время для постоянных давно существующих водоёмов или протекает на наших глазах в водоёмах временного значения. В процессе эволюционного развития устанавливается более или менее определенный видовой состав биоценоза, который стабилизируется или, как говорят, находится в состоянии подвиж кого равновесия; оно касается как взаимоотношений сочленов биоценоза друг с другом, так и зависимости их и биоценоза в целом от факторов внешней среды.
Необходимо помнить, что наблюдаемый нами состав любого биоценоза является лишь этапом истории его существования; он, конечно, не стабилен в абсолютном смысле этого понятия, но сохраняет более или менее долго постоянство своего видового состава, если не происходит каких-либо особых изменений, например вторжения новых обитателей, нарушающих установившуюся зависимость сочленов биоценоза друг от друга, или колебаний факторов внешней среды, в количественном или в качественном отношениях выходящих за обычные пределы их амплитуды. Следствием таких изменений может явиться полный распад существовавшего ранее биоценоза или его явное видоизменение; последнее выражается в том, что один биоценоз сменяется другим, другой - третьим и т. д. Такое явление называется экологической сукцессией биоценоза. Темпы сукцессии вариируют от нескольких дней для временных водоёмов и до геологических сроков летоисчисления, если рассматривается история населения древних водоёмов. Особо выделяются сезонные сукцессии, то есть изменения состава биоценоза, связанные с переменою времен года. Например, зимою мы не найдем в составе биоценоза куколок комаров сем. Culicidae, различные виды их зимуют по разному - то в фазе имаго (Culex pipiens, Anopheles maculipennis), то в состоянии отложенных яиц (виды комаров рода Aedes). Некоторые же водные обитатели остаются вполне активными зимою подо льдом, например некоторые ветвистоусые рачки; но в их поколениях происходят изменения внешних признаков в связи с зимними изменениями свойств воды как среды их обитания (см выше данные о цикломорфозе дафний).
Биоценотические связи при обилии сочленов биоценоза представляют большую сложность, в которой нелегко разобраться. Поэтому целесообразно выделять ведущий сочлен биоценоза и уже по отношению к нему определять важнейшие взаимосвязи с прочими сочленами того же сообщества, а также и с биоценозами другого порядка, если между ними имеется какая-либо реальная связь (например, зависимость обитателей водоёма от наземных животных, для которых они служат пищей).
Понятие "ведущий сочлен биоценоза" выдвигается применительно к конкретным условиям места и к поставленным целям исследования (Е. Павловский, 1937). Такая установка работы нисколько не снижает её теоретического значения, наоборот, она связывает теоретическое исследование с практикой применения его выводов для хозяйственных или санитарных целей. Подобная задача может быть весьма сложной, вследствие множественности взаимосвязей и многочисленности членов биоценозов, помимо того, взаимосвязи избранного нами сочлена биоценоза бывают различными не только в разных фазах его метаморфоза, но и во время его роста. Например, пищевой режим сельдей в Северном море в различных возрастах различен, причем цепи питания их отличаются своеобразием. Подобные отношения можно наблюдать и в пресноводных бассейнах. Мы подчеркиваем такое положение дела не для расхолаживания исследователя возможными трудностями, но для правильной ориентировки его в работе и в ее последовательности.
Биоценотические связи пищевого характера принято изображать на схематических чертежах стрелками между двумя соответственными членами биоценоза, направленными от поедающего к поедаемому организму; последний, следовательно, всегда обозначается острием стрелки.
Остановимся на нескольких различных по своему характеру примерах.
Прежде всего запомним, что биоценоз водоёма нельзя рассматривать изолированно от воздушно-наземной среды, не только вследствие физико химических зависимостей (их мы здесь не рассматриваем), но и благодаря влиянию на наблюдаемый биоценоз некоторых биотических факторов наземной среды (например, наземные животные, которые питаются рыбой и другими водными животными). На предлагаемых схемах показан и человек; он, конечно, не может считаться сочленом какого-либо водного биоценоза, но он неминуемо сталкивается с его сочленами или сознательно (например, при добывании пищи) или бессознательно, активно или пассивно (когда активность в достижении контакта с человеком проявляется со стороны животных биоценоза). Вопрос о соотношении человека с биоценозом водоёма и о характере влияния и форм использования последнего человеком весьма многосторонен и очень важен практически; его мы рассмотрим в заключительной главе нашей книги.
Из наземных биотических факторов, помимо животных, на биоценоз водоёма влияют растения, что сказывается часто и на окончательной судьбе водоёма. В качестве примера можно привести зарастание водоёма растениями прибрежной зоны, или длительное зарастание болот с превращением их в торфяники. Этим вопросам посвящена большая специальная литература.
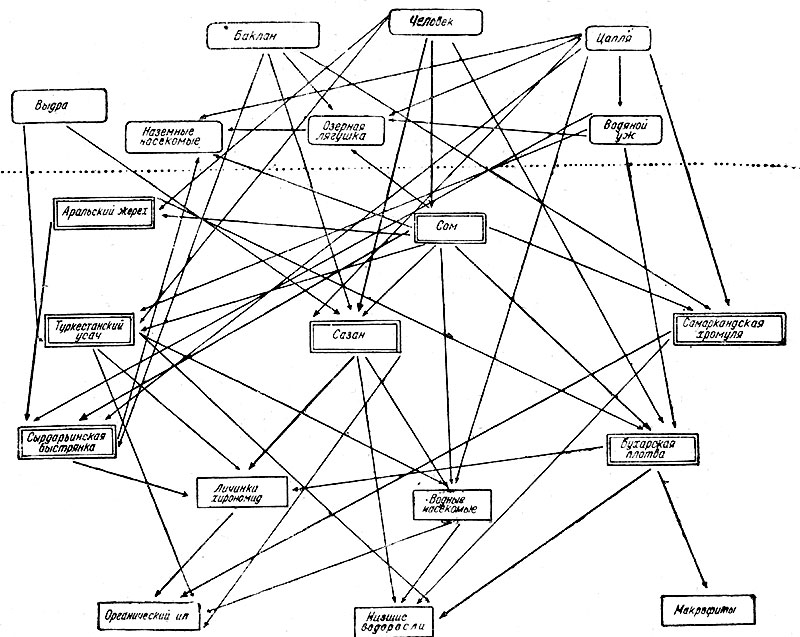
Рис. 14. Схема пищевых связей в озерах равнинного течения Аму-Дарьи (по Никольскому)
В качестве примера конкретных пищевых связей водного биоценоза рассмотрим схему основных связей в озёрах равнинного течения бассейна р. Аму-Дарьи в Таджикистане (по Г. В. Никольскому, 1938, рис. 14).
Первые звенья пищевых цепей представляют органический ил, водоросли и макрофиты (высшие водные растения). Органический ил служит пищей для личинок тендипедид, которые в свою очередь являются преобладающей пищей для карпа (Cyprinus carpio), используемого для питания человеком и выдрой (Lutra lutra) налицо четырехзвенная пищевая цепь. В качестве побочных пищевых веществ карп потребляет водоросли, органический ил и различных водных насекомых, питающихся обоими поименованными, объектами. Органический ил служит главной пищей для хромули (Varicorhinus heratensis), являющейся добычей для сома и цапли (Ardea) и баклана (Phalacrocorax). Бухарская плотва (Rutilus bucharensis) в дополнение к водорослям поедает высшие водные растения и личинок тендипедид; сама же делается добычей жереха (Aspius aspius), водяного ужа (Natrix tesselata) и человека. Водяного ужа пожирает цапля (Ardea). Особенно многообразны пищевые связи сома (Silurus glanis); помимо упомянутых рыб, он пожирает быстрянку (Aiburnoides teniatus), усача (Barbus capIto), жереха (Aspius aspius) и водных насекомых; из земноводных он охотится за лягушками (Rana ridibunda); кроме того, сом потребляет и наземных насекомых. Быстрянка (Alburnoides teniatus), питающаяся личинками тендипедид и наземными насекомыми, поедается сомом (Silurus glanis), жерехом (Aspius aspius), водяным ужом и птицами (Phalacrоcorax, Ardea).
Из наземных животных многообразна пища цапель: в водоёме - водные насекомые, рыбы-хромули, быстрянки, на суше - наземные насекомые, лягушки, ужи.
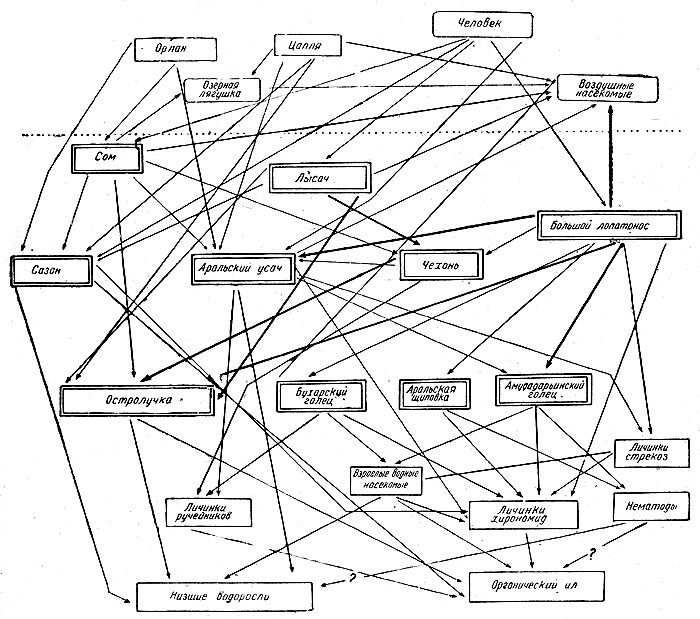
Рис. 15. Схема пищевых связей в русле Аму-Дарьи в равнинном Таджикистане (по Никольскому)
На этих примерах видим, образцы пятизвенных пищевых цепей (ил, личинки тендипедид, бухарская плотва, водяной уж, цапля).
В русле Аму-Дарьи в равнинном Таджикистане в зависимости от большого богатства фауны соответственно умножаются и пищевые связи, о которых дает представление рис. 15; в горной же части бассейна Аму-Дарьи в пределах того же Таджикистана эти отношения оказываются более упрощенными, что зависит от обеднения водной фауны (рис. 16).
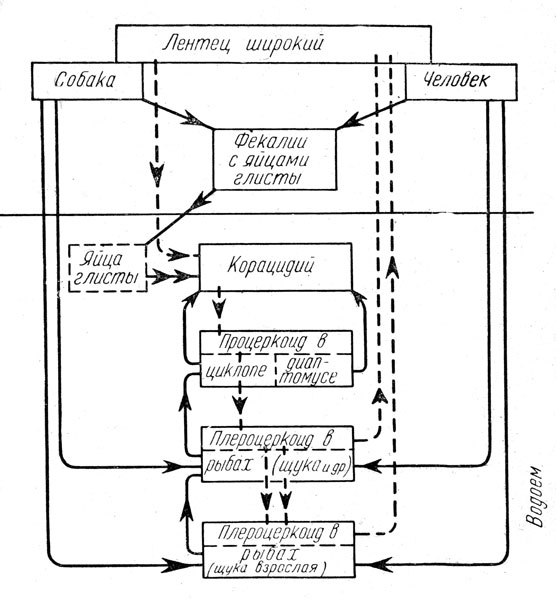
Рис. 16. Схема пищевых связей в горной части р. Аму-Дарьи в Таджикистане (по Никольскому)
Для третьего примера рассмотрим биоценоз замкнутого водоёма средней полосы СССР, избрав в качестве ведущего сочлена биоценоза водные фазы метаморфоза обыкновенного малярийного комара (Anopheles maculipennis), его личинок, куколок и яйца (рис. 17). Одновременно следует рассмотреть и пищевые связи самок того же вида, являющихся воздушно-наземными обитателями. Пищевые связи имеют лишь личинки анофелес во всех стадиях их; яйца й куколки его ничем не питаются, но сами служат пищей другим животным.
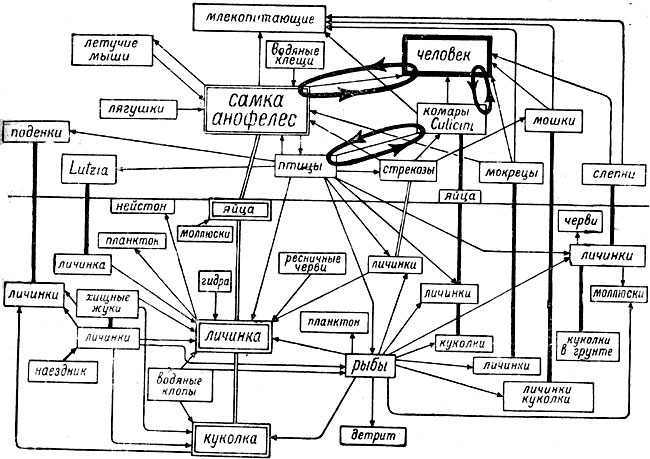
Рис. 17. Пищевые связи малярийного комара в биоценозе мелкого замкнутого водоёма. Замкнутые эллипсы (жирные линии) - пути циркуляции болезнетворного начала (плазмодии и др.), (Оригинальная схема Е. Н. Павловского)
Личинки анофелес питаются неистовом и частично приставшим к пленке воды планктоном и микробентосом; сами же делаются жертвами множества животных: их пожирают гидры, ресничные черви, при особых условиях улитки, личинки стрекоз, ручейников, жуков и различных двукрылых, хищные жуки, водяные клопы (гладыш, водяной скорпион, ранатра), рыбы, а из водных растений - пузырчатка. Из наземных животных их, а также куколок, поедают птицы (утки). Куколки делаются добычей многих из перечисленных обитателей водоёма; яйца анофелес поедаются улитками.
Самки анофелес в воздушной среде пьют кровь человека, многих млекопитающих и птиц, служа в свою очередь пищей для стрекоз, летучих мышей и птиц. Побочные пищевые связи указаны на схеме (рис. 17). Личинки и куколки анофелес держатся у поверхности водоёма, что делает их весьма доступными для нападения многих животных.
Пищевые связи водных фаз метаморфоза, например, слепней, личинки и куколки которых ведут придонный образ жизни, естественно беднеют в своем разнообразии. Примером могут служить слепни (Tabanus). На этом примере мы увидим различие биоценотических связей у личинок Tabanus в разных стадиях их водной жизни. Дело в том, что выходящие из яиц личинки Tabanus, падая в воду, не тонут, так как их широкие трахейные стволы играют роль плавательного аппарата; это свойство их сохраняется до 2-3 стадии метаморфоза (Всего личиночных стадий у слепней семь). Личинки же слепней златоглазиков (Chrysops coecutiens) сразу опускаются на дно; следовательно, биоценотические связи их, так же как и личинок поздних стадий превращения Tabanus, будут существенно иными, нежели у плавающих у поверхности воды личинок 1-3 стадий Tabanus; опасность гибели также соответственно различна у свободно плавающих личинок и у личинок, зарывающихся в ил дна. На рис. 18 дана схема пищевых связей придонных личинок Tabanus. Они являются хищниками или полухищниками и пожирают личинок различных насекомых, связанных в своем пребывании с дном водоёма, мелких моллюсков, червей и других животных, а также органический детрит; главной пищей личинок слепней златоглазиков являются растительные остатки. Сами они могут делаться жертвой личинок стрекоз и водяного скорпиона. Для окукления личинки заползают в толщу берега выше уровня воды.
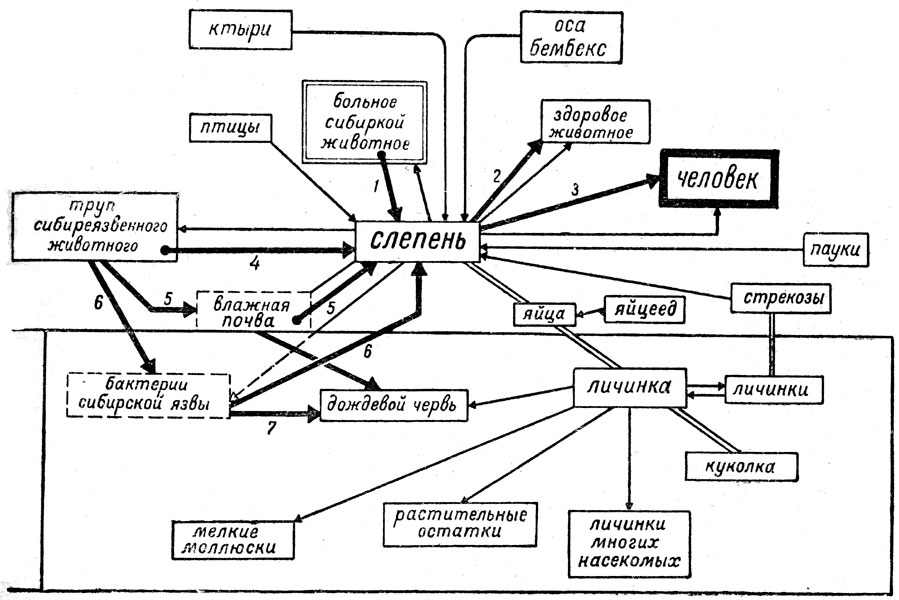
Рис. 18. Пищевые связи слепня и пути циркуляции возбудителя сибирской язвы (жирные линии) в биоценозах водоёма и воздушно-наземной среды (оригин. схема Е. Н. Павловского)
Яйца Tabanus, только что отложенные кучкой над водой, делаются часто добычей яйцееда Telenornus tabani, причем этот паразит откладывает яйца лишь в поверхностно лежащие яйца Tabanus. Всё развитие Telenomus в яйцах слепня длится до 30-35 дней.
Распределение личинок слепней по биотопам зависит от их активной подвижности в поисках пищи и от пассивного перераспределения перезимовавших личинок весенними потоками воды в малых ручьях и половодьем рек в больших потоках. Благодаря этому личинки слепней могут осесть в новых местах: в песчаных наносах берегов, в излучинах ручьев и рек - в отдалении от точек, где они впервые попали в воду (Олсуфьев).
Пищевые связи самок слепней определяются формой их питания - кровососанием; объектами их нападения являются дикие и домашние животные и человек. В сезон своего массового развития слепни являются важным компонентом "гнуса", т. е, всей совокупности летающих кровососущих двукрылых, в своем жизненном цикле связанных с водоёмами (комары, мокрецы, мошки, рис. 19).

Рис. 19. Биотопы симулиид в горном Таджикистане (по Павловскому)
В заключение этой главы подчеркнем, что прежняя фаунистическая работа, ограничивавшаяся установлением видового состава животного населения какого-либо места, переросла в исследование эколого-фаунистического характера, когда видовой состав фауны изучается в современных и по возможности в исторических взаимоотношениях видов друг с другом и с внешней средой. В этом направлении особенно удобно и наглядно исследование пресноводных объектов, исключительно разнообразных по величине и характеру их флоры и фауны и длительности существования; многие из них широко доступны для полевых исследований, в параллель с которыми идут экспериментальные работй в лаборатории. На водных объектах, заслуживающих особого внимания, организованы пресноводные биологические станции. Таковы, Байкальская лимнологическая станция Академии Наук СССР, Биологические станции под Москвой (на озере Глубоком и в Болынове), Лимнологическая станция на озере Севан, Биологическая станция "Залучъе" (под Бологое), являющаяся Отделением Зоологического Института Академии Наук СССР в Ленинграде, Биологическая станция Карело-Финского университета и Бородинская станция Ленинградского о-ва естествоиспытателей на озёрах Карелии. В различных местах СССР энергична работают по исследованию пресных вод отделения Всесоюзного Научно-исследовательского Института Рыбного Хозяйства. Большая работа ведётся в этом направлении многочисленными малярийными станциями, а на юге Союза - Институтами малярии (Тропинституты).
Приходят время от времени вести об организации новых станций в разных местах СССР. Остается пожелать, чтобы сеть их росла и развивалась.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'