Подкласс Открыточелюстные, или настоящие, насекомые (Insecta-Ectognatha)
Основные сведения о насекомых
Из общего числа видов животных, населяющих Землю, на долю насекомых приходится около 70%. Число уже описанных видов приближается к миллиону, но ежегодно специалисты открывают и описывают все новые и новые тысячи видов, и, по принятым в настоящее время подсчетам, на земном шаре обитает более 2 миллионов различных видов насекомых.
Основная масса насекомых - обитатели cyши, и здесь насекомые распространены необычайно широко. Исключительно богат и разнообразен мир насекомых в тропиках. В тропических лесах богатство видами многих групп насекомых, в том числе наиболее бросающихся в глаза, например бабочек, так велико, что легче встретить новый вид, чем другой экземпляр уже встреченного! В более умеренных широтах число видов насекомых не так велико, но зато общая их масса огромна. Академик В. И. Вернадский, создатель науки биогеохимии, образно показал, что, например, вес одной стаи саранчи, однажды перелетавшей через Красное море в начале нашего века, превышал вес всех цветных металлов, включая свинец и медь, выплавленных человечеством за всю историю до этого дня!
Насекомые заходят высоко в горы. Их можно встретить на огромных высотах. Например, мелкая жужелица Bembidion bracculatum в Гималаях населяет только пояс от 4300 до 5000 м, приспособившись к жизни в этих суровых условиях, а всего из зоны вечного снега в Гималаях описано 25 видов только этого рода жужелиц!
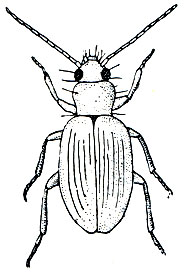
Рис. 104. Гималайская жужелица Bembidion bracculatum
Насекомые населяют и абсолютно безводные пустыни, такие, как, например, пустыня Намиб в Южной Африке, где никогда не бывает ни капли дождя, где нет никаких растений и где живут жуки-чернотелки (Gonopus, Syntyphlus subterraneus), питающиеся только приносимыми за сотни километров знойным ветром остатками растений, отлагающимися у оснований барханов. Необходимую для жизни влагу чернотелки получают биохимическим путем, используя воду, которая образуется при окислении углеводов сухой, недоступной другим организмам пищи!
Много различных насекомых обитает в пещерах. Там, где нет, кажется, никакого поступления органических веществ, в обстановке вечной тьмы, встречаются слепые представители этой группы, чаще всего жуки, использующие те ничтожные запасы пищи, которые удается отыскать.
Первое пещерное насекомое было открыто в пещере Постойна на территории Югославии - жук Leptodirus hohenwarti - в 1831 году, а с тех пор из разных стран описана богатая и удивительная фауна насекомых-троглобионтов.
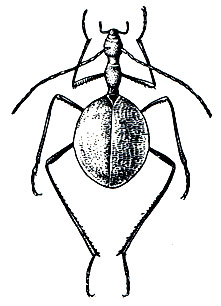
Рис. 105. Пещерный жук Leptodirus hohenwarti
Очень далеко заходят насекомые и в высокие широты: за полярным кругом, на почти голых скалах, покрытых большую часть года снегами и льдом и лишь на очень короткий период освобождающихся от зимних оков, среди лишайников и мхов можно встретить мелких представителей этого класса.
Даже на островах Антарктики недавно найдены бескрылые комары-дергуны, как антарктический комар (Belgica antarctica) - родич нашего мотыля, красные личинки которого хорошо известны рыболовам и любителям аквариумных рыб.
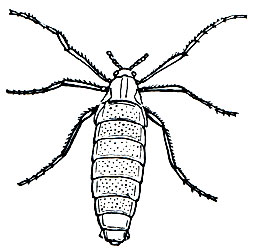
Рис. 106. Антарктический комар (Belgica antarctica)
Приспособившиеся к полету крылатые насекомые завоевали воздушную среду - лучшие летуны среди них активно поднимаются на высоту нескольких сотен метров (двукрылые, бабочки), но можно встретить насекомых и выше, на высоте нескольких километров. Сюда они заносятся восходящими токами нагревающегося у поверхности земли воздуха и затем ветром переносятся на огромные расстояния. В высоких слоях атмосферы чаще встречаются не хорошо летающие, а легко парящие в воздухе насекомые - мелкие мушки, в основном злаковые мушки (Agromyzidae), тли, трипсы. Но не только крылатых насекомых можно встретить на высоте нескольких десятков и сотен метров. Легкие плоские маленькие личинки щитовок (Coccidae, рис. 217, 3), сдуваемые порывами ветра с ветвей деревьев, тоже поднимаются с пылью высоко вверх и так расселяются. Высокой способностью к расселению насекомых объясняется то, что на всяком океаническом островке, удаленном за тысячи километров от остальной суши, можно найти насекомых. А на поверхности суши они заселяют все растения, поднимаясь к вершинам самых высоких деревьев. В тропических лесах Африки, например, есть такие насекомые, которые еще недавно считались исключительно редкими, а оказались обычными, но постоянно обитающими высоко в кронах гигантских деревьев; они недоступны для сборов без пожарных лестниц или вертолетов. Все участки крон деревьев, весь ковер травянистых растений, мхов и лишайников, вся почва на глубину того слоя, куда проникают корни растений, и даже глубже осваиваются насекомыми. В Туркмении, например, закаспийский термит (Anacanthotermes ahngerianus) прокладывает свои ходы на глубину до 12 метров!
Некоторые насекомые приспособились к жизни в тканях растений, к развитию в теле различных животных - от губок до млекопитающих и человека, к использованию всевозможных очень специфических местообитаний, самых разных источников пищи.
По способности жить на суше в самых различных условиях класс насекомых не имеет себе равных и именно благодаря тому, что отдельные виды приспособились к жизни в самых разных, иногда невероятных условиях (например, личинки некоторых мух в горячих источниках или в насыщенных солевых растворах), насекомые в процессе своей бурной эволюции, продолжающейся уже во всяком случае более 200 миллионов лет, дали то исключительное богатство видов, которое обеспечивает современное процветание этой группы членистоногих.
От жизни на суше многие насекомые перешли к жизни в воде. Уже в каменноугольном периоде палеозойской эры в воде развивалось большое количество видов насекомых, и в частности вымерший в конце этого периода отряд палеодиктиоптер (Palaeodictyoptera, рис. 185), некоторые представители которого были много крупнее, чем самые крупные из ныне живущих насекомых (до 60 см). Завоевали пресные водоемы и стали водными в полном смысле слова, т. е. дышащими за счет того кислорода, который растворен в воде, только личинки насекомых, взрослая окрыленная стадия которых способна оставлять воду. Если же насекомое и во взрослом состоянии живет в воде, то оно дышит кислородом воздуха, поднимаясь к поверхности. Только некоторые, как например мелкие (0,2 мм) наездники - карафрактусы (Caraphractus) и прествичии (Prestwichia aquatica, рис. 371, 3, 4), развивающиеся в яйцах жуков-плавунцов, ведут во взрослом состоянии совершенно подводный образ жизни.
Некоторые насекомые, как например клопы-водомерки (Hydrometridae и Gerridae), живут на поверхности воды, бегая по ней, отталкиваясь длинными, как у пауков, ногами от пленки поверхностного натяжения.
Близкие к нашим водомеркам представители семейства Halobatidae - морские водомерки - бегают по поверхности морей, а Halobates micans в тропическом поясе обитает в океанах за сотни километров от берега. Правда, нельзя морских водомерок назвать настоящими водными насекомыми- они живут на поверхности воды, питаются всплывающими трупами животных, а яйца откладывают на плавающие водоросли, перья птиц и даже на себе подобных. Настоящих морских насекомых, развивающихся в морской воде, очень мало - немногим более десятка, все из отряда двукрылых. Только один вид комаров-долгоножек (Limonia monostromia), обитающий у берегов Японских островов, в течение всего цикла развития не оставляет моря.
Насекомые как класс сформировались на суше и происходят от уже наземных многоножкообразных животных, этим и объясняется малое количество среди них морских форм. При этом все морские насекомые - представители наиболее высокоорганизованных отрядов.
Размеры насекомых варьируют в очень широких пределах: самые крупные превосходят по длине тела самых мелких примерно в 1000 раз. Самые крупные представители современных насекомых - обитатели тропиков. Среди жуков жук-геркулес (Dynastes hercules, рис. 262) и жук-слон (Megasoma elephas) достигают длины 12 см; палочник Phybalostoma cervicornis имеет длину 26 см; хищный водный клоп Belostoma grande - 11,5 см; шелкопряд Attacus atlas в размахе крыльев достигает 24 см, а бабочка Thysania agrippina (рис. 328) - 26 см. Даже в фауне Европейской части СССР есть крупные насекомые. Наш большой дубовый усач (Cerambyx cerdo, рис. 284, 2) достигает длины 5 см, большой ночной павлиний глаз (Saturnia pyri, табл. 47, 4) -15 см в размахе крыльев. Самые же мелкие из насекомых имеют длину меньше 0,25 мм (жуки-перистокрылки - сем. Ptiliidae, наездники-яйцееды- сем. Mymaridae). Таким образом, самые мелкие насекомые мельче, чем самые крупные простейшие, а самые крупные - больше, чем самые мелкие млекопитающие. Несмотря на огромное разнообразие условий и образа жизни, на исключительные различия размеров и обилие видов, чаще всего, увидев насекомое, можно с уверенностью сказать, что это насекомое.
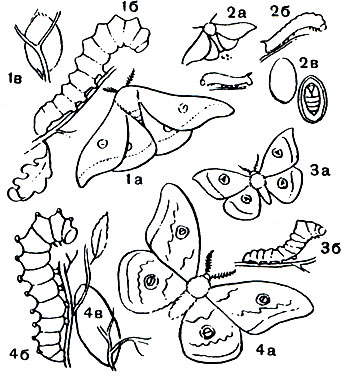
Таблица 47. Тутовый шелкопряд и павлиноглазки: 1 - китайская дубовая павлиноглазка (Antheraea pernyi); 2 - тутовый шелкопряд (Bomdyx mori); 3 - рыжий павлиний глаз (Aglia tau); 4 - большой ночной павлиний глаз (Saturnia pyri); а - бабочка, б - гусеница, в - кокон

Таблица 47. Тутовый шелкопряд и павлиноглазки: 1 - китайская дубовая павлиноглазка (Antheraea pernyi); 2 - тутовый шелкопряд (Bomdyx mori); 3 - рыжий павлиний глаз (Aglia tau); 4 - большой ночной павлиний глаз (Saturnia pyri); а - бабочка, б - гусеница, в - кокон
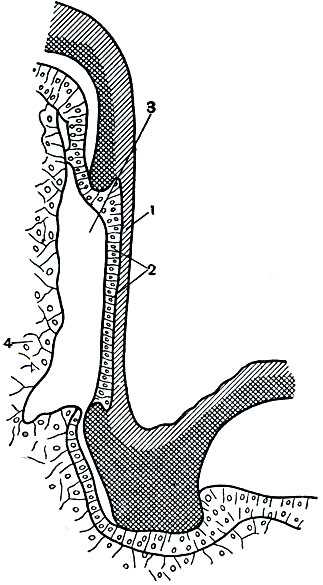
Для насекомых характерно то, что тело их разделено на 3 отдела - голову, грудь и брюшко, каждый из которых выполняет определенную функцию. На голове находятся основные органы чувств, позволяющие ориентироваться в пространстве, и ротовые органы - органы захватывания пищи. В грудном отделе у насекомых сосредоточены основные органы передвижения - двигательные конечности (ноги), а у взрослых высших насекомых - крылья. Брюшко - отдел, в котором сосредоточена основная масса внутренних органов, включая среднюю и заднюю кишку, жировое тело, выделительные и половые органы, совокупительный аппарат или яйцеклад и т. д. Такое деление тела на отделы (рис. 107) хорошо прослеживается у большинства взрослых насекомых и бывает не выражено либо у личинок, либо у таких взрослых насекомых, строение которых вторично изменилось под влиянием неподвижного или паразитического образа жизни.
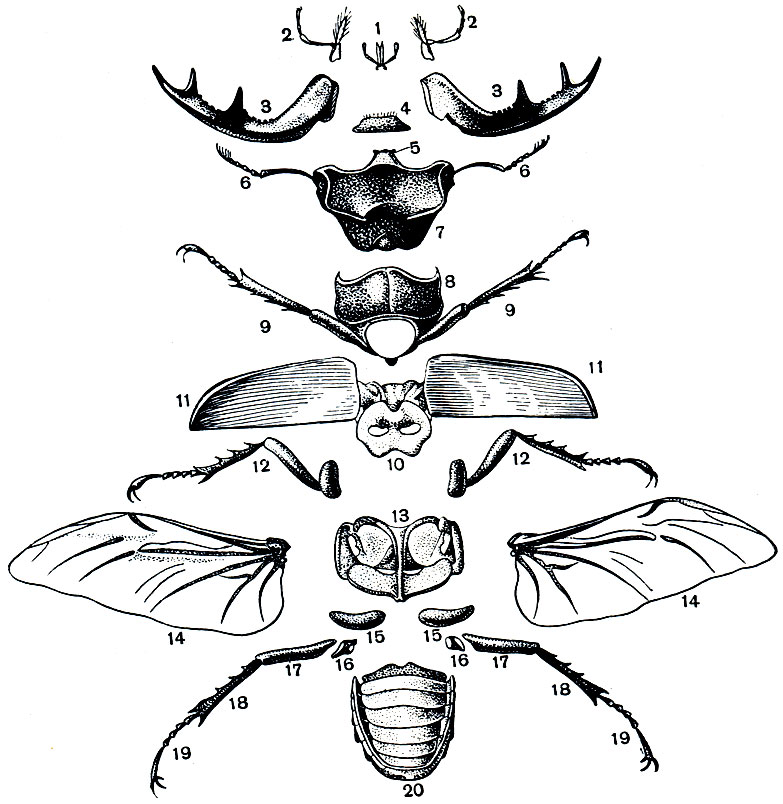
Рис. 107. Расчлененный самец жука-оленя (Lucanus cervus): 1 - нижняя губа с щупиками; 2 - нижние челюсти с щупиками; 3 - жвалы (верхние челюсти); 4 - верхняя губа; 5 - наличник; 6 - усики; 7 - голова; 8 - переднегрудь; 9 - передние ноги; 10 - среднегрудь; 11 - надкрылья; 12 - средние ноги; 13 - заднегрудь; 14 - крылья; 15 - тазики задних ног; 16 - вертлуги; 17 - бедра; 18 - голени; 19 - членистая лапка с коготками; 20 - брюшко
Голова образуется из 6 сегментов, закладывающихся у зародыша, но у сформировавшегося насекомого границы сегментов головы не видны - сегменты полностью сливаются.
Грудных сегментов у насекомых всегда 3. Каждый грудной сегмент несет по паре ног, а у многих насекомых ко второму и третьему сегментам причленяются органы полета - крылья, представляющие не конечности, а специализированные складки. В грудном отделе сосредоточена основная двигательная мускулатура насекомых.
Брюшко у насекомых образуется за счет закладки не более 12 зародышевых сегментов, у большинства высших десятый сегмент является последним. В некоторых случаях передние брюшные сегменты сливаются с грудным отделом, явственно отделяясь от остального брюшка перехватом. Такой случай наблюдается у жалоносных перепончатокрылых, например у пчел. Еще реже встречается слияние грудных сегментов с головой, как это имеет место у самок веерокрылых (Strepsiptera, рис. 307, 2; 309, 2), но в этом случае слияние связано с общим регрессом в развитии.
Наружный покров у насекомых представлен сложной кутикулой, выделяемой слоем расположенных под ней клеток - гиподермой. Кутикула покрывает не только открытые участки тела, но выстилает и передний отдел кишечника (рот, глотку, пищевод), заднюю кишку, ближайшие к дыхальцам участки трахейных трубок. У наземных насекомых в кутикуле различаются прилегающий к живым клеткам внутренний слой - эндокутикула, очень эластичная, составляющая основную часть толщи панциря, следующая за ней механически прочная экзокутикула и самая наружная, содержащая много жироподобных и воскоподобных веществ эпикутикула, имеющая ничтожную толщину - обычно менее 1 мк. Эпикутикула не пропускает воду. Она не развивается у большинства личинок насекомых, обитающих во влажной почве, в воде или в тканях растений. Чем суше та среда, где живет насекомое, тем яснее у него выражена эпикутикула. Естественно, что в кутикуле, выстилающей кишечник, нет непроницаемого слоя - эпикутикулы.
На теле насекомых кутикула развита неравномерно. Особенно сильно она выражена на сегментах; между сегментами находится нежная перепонка, состоящая из гораздо более тонких эндокутикулы и экзокутикулы. Так как складки перепонки обычно бывают скрыты под более уплотненными щитками, на ней эпикутикулы иногда нет.
Уплотненные толстые щитки на сегментах называются склеритами. Они развиты сильнее там, где больше требуется защита от повреждении, и там, где прикрепляются мышцы. Склериты туловища у насекомых расположены симметрично с правой и левой сторон, но на спинной стороне правый и левый склериты часто сливаются, образуя единый щиток - тергит. Склериты по бокам тела называются плейритами, а на брюшной стороне - стернитами.
Голова у насекомых (табл. 14, 15) почти всегда сильно склеротизована, часто сильнее, чем другие отделы. Это объясняется тем, что прочная сплошная головная капсула у насекомых служит местом прикрепления мощной челюстной мускулатуры. Вспомним, что возникновение черепной коробки и у позвоночных было связано с этой же функцией. Слияние сегментов, из которых состоит голова, настолько велико, что у сформировавшегося насекомого никаких следов границ сегментов нет. Швы, которые наблюдаются на голове (рис. 108),- это не границы сегментов, а либо места прикрепления мышц, либо те линии, по которым идет растрескивание черепной капсулы при линьке. Но при изучении эмбрионального развития насекомых установлено, что голова у них образована за счет 6 зародышевых сегментов. Первому сегменту соответствуют глаза, второй сегмент несет усики, третий сегмент ("вставочный") не несет придатков и сливается со вторым, на четвертом, пятом и шестом сегментах развиваются парные части ротового аппарата - верхние челюсти (жвалы, или мандибулы), нижние челюсти (максиллы) и нижняя губа. Эти ротовые части у зародыша закладываются точно так же, как закладываются ноги на следующих сегментах тела. Это дает основание считать ротовые части насекомых конечностями, вошедшими в состав ротового аппарата. Что конечности могут входить в состав ротового аппарата, хорошо видно на примере губоногих многоножек (см. выше), у которых первая пара туловищных ног, сохраняя еще типичное для ног расчленение, стала действовать в сущности как мандибулы. То, что головная капсула имеет значение как место прикрепления мышц, доказывается тем, что, например, у личинок мух, питающихся уже полностью переваренной пищей, головы нет.
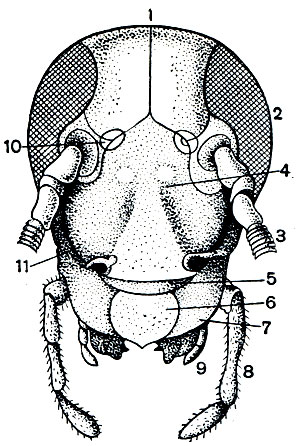
Рис. 108. Голова черного таракана (Blatta orientalis) спереди: 1 - темя; 2 - сложный глаз; 3 - усик; 4 - лоб; 5 - наличник; 6 - верхняя губа; 7 - жвала (верхняя челюсть); 8 - щупик нижней челюсти; 9 - щупик нижней губы; 10 - простой глазок; 11 - щека

Таблица 14. Разнообразие формы головы насекомых. Голова бабочки-желтушки (Colias myrmidone)

Таблица 14а. Разнообразие формы головы насекомых. Голова долгоносика Curculio pellitus
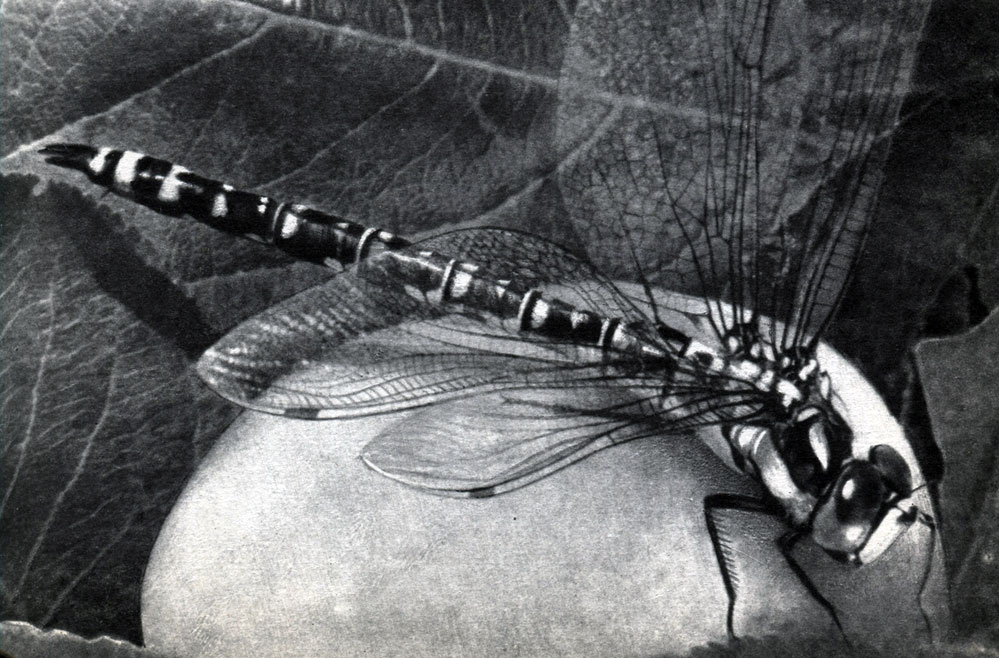
Таблица 14б. Разнообразие формы головы насекомых. Стрекоза-коромысло (Aeschna) с крупными фасеточными глазами

Таблица 15. Шмель Bombus hypnorum, вид спереди
Глаза у насекомых бывают двух типов - фасеточные, как и у высших ракообразных, и простые. Фасеточные глаза всегда располагаются по бокам головы и свойственны большинству взрослых насекомых (табл. 14). Иногда они занимают большую часть поверхности головы и у многих хорошо летающих насекомых число фасеток в них огромно. У стрекоз, например, в сложном глазу бывает до 28 000 фасеток, у бабочек - до 17 000, у комнатной мухи - 4 000.
У видов насекомых, разные особи которых ведут неодинаковый образ жизни, можно наблюдать зависимость сложности фасеточного глаза от биологических особенностей. Например, у муравья-вора (Solenopsis fugax) бескрылые рабочие особи, живущие в ходах почвы, имеют глаза, состоящие из 6-9 фасеток. В глазах самок, летающих во время роения, фасеток около 200, а у самцов, которые во время полета должны найти и опознать самок своего вида, в глазу свыше 400 фасеток. У некоторых рабочих муравьев (Ponera punctissima) в каждом глазу по одной фасетке.
У личинок насекомых с полным превращением и у таких паразитических форм, как вши, а также у личинок щитовок (Coccidae) на месте сложных глаз развиваются боковые простые глазки. Иногда простых глазков бывает много (более 30 у личинок скорпионниц) и они выглядят как фасеточный глаз. Между фасеточным глазом и скоплением глазков основная разница заключается в том, что каждый простой глазок имеет свою кутикулярную роговицу, а в фасеточном глазу все омматидии, т. е. отдельные глазки сложного глаза, имеют общую роговицу.
Наряду со сложными глазами у многих хорошо летающих насекомых имеется по 3 или 2 простых глазка, расположенных между сложными глазами на лбу.
Усики (иначе называемые антеннами) у насекомых располагаются в передней части головы. Вариации строения усиков у насекомых невероятно широки, но, как правило, целые семейства, подотряды или отряды насекомых характеризуются той или иной определенной формой усиков. Обычно усики состоят из большого числа члеников, но иногда они короткие и члеников в них немного - у мух и стрекоз, например, в усиках всего по 3 членика, а у многих личинок, например у личинок долгоносиков и ручейников, вместо усиков имеются только едва заметные рудиментарные сосочки, у личинок мух усиков нет. У жуков усики обычно 11-члениковые, у а перепончатокрылых они состоят из 12-13 члеников. Иногда число члеников в усиках бывает очень велико, так как часто исходные членики распадаются на множество колец. У таких насекомых, как, например, кузнечики и некоторые жуки-дровосеки, усики бывают длиннее тела (рис. 154, 283, 284; табл. 29, 4; 40, 6, 43, 3).
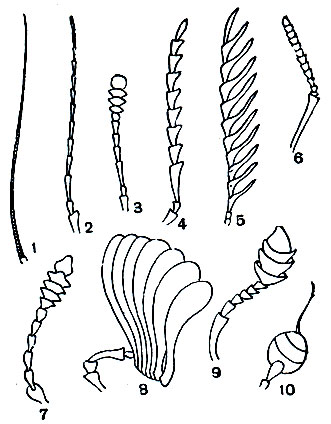
Рис. 109. Типы усиков насекомых: 1 - щетинковидный; 2 - нитевидный; 3, 7 - булавовидный; 4 - пильчатый; 5 - гребенчатый; 6 - коленчатый; 8 - пластинчатый; 9 - коленчато-булавовидный; 10 - булавовидно-щетинковидный
Различают много форм усиков. Сужающиеся равномерно к вершине называются щетинковидными (как у таракана). Усики нитевидные, если все членики одинаковой толщины и каждый членик цилиндрический. Примером могут служить усики жужелиц (рис. 251). Если у члеников усиков середина усика толще, чем основание и вершина, усики четковидные, как у термитов. У таких жуков, как щелкуны, усики гребенчатые (рис. 268, 5). Если от каждого членика усиков в обе стороны отходит по тонкому выросту, усики перистые, как у ночных бабочек-шелкопрядов. Бывает, что усик на большей части своей длины более или менее одинаковой толщины, а у вершины расширяется - это булавовидный усик (у дневных бабочек). Если первый членик усика длинный, а последующие располагаются под углом к нему, усики коленчатые. Иногда у коленчатых усиков бывает булава, состоящая из веерообразно сложенных пластинок (пластинчатоусые жуки) или из гребешков (жук-олень). Бывают различной формы неправильные усики (рис. 247), иногда с причудливыми выростами, как у жука-нарывника из рода Cerocoma.
Ротовой аппарат включает 3 пары придатков, развивающихся как конечности. Наиболее хорошо выражены все ротовые части у представителей более древних групп насекомых, имеющих грызущий ротовой аппарат, при питании отгрызающих твердые кусочки пищи Таковы тараканы, саранчовые и многие другие (рис. 110).
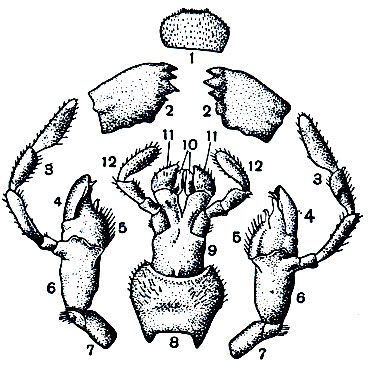
Рис. 110. Ротовой аппарат черного таракана: 1 - верхняя губа; 2 - жвалы (верхние челюсти); 3-7 - нижние челюсти: 3 - щупик нижней челюсти, 4 - наружная жевательная лопасть, 5 - внутренняя жевательная лопасть, 6 - стволик, 7 - основной членик; 8-12 - нижняя губа: 8 - подподбородок, 9 - подбородок, 10 - язычки, 11 - добавочные язычки, 12 - щупики нижней губы
Первая пара челюстей - верхние челюсти - у всех насекомых состоит из цельных, нерасчлененных пластинок (жвал). По форме жвал бывает довольно легко установить, какой пищей питаются насекомые. У потребителей растительной пищи поверхность вершин жвал широкая, зазубренная, приспособленная к отрыванию твердых тканей, а у основания жвал широкая бугорчатая перетирающая поверхность - полная аналогия резцам и коренным зубам травоядных млекопитающих. Такие жвалы - у саранчовых, гусениц, многих жуков. А у хищников, таких, как жуки-скакуны, личинки златоглазки и др., жвалы серповидно изогнутые с острыми концами. Такие жвалы аналогичны серповидно изогнутым клыкам хищных млекопитающих - клыкам, приспособленным к пронзанию тела добычи и разрыванию его. Жуки-скакуны по-английски недаром даже называются tigerbeetles - "жуки-тигры" - и за хищный образ жизни, и за сходство их мандибул с клыками саблезубого тигра.
Так характер пищи накладывает отпечаток на строение верхних челюстей, играющих наиболее ответственную роль в грызущем ротовом аппарате. Мандибулы бывают очень прочными. Достаточно сказать, что известны случаи, когда личинки земляных усачей повреждали своими мандибулами свинцовую оболочку подземного электрического кабеля, а жуки-короеды и точильщики легко прокладывают ходы в твердой древесине дуба.
У хищных личинок жуков-плавунцов серповидные мандибулы служат не только для прокалывания добычи. В каждой мандибуле у них есть желобок, через который личинка сперва вводит пищеварительные соки в тело жертвы, а затем, когда внутри тела схваченного животного белки его мышц разжижаются и перевариваются впрыснутыми ферментами, личинка плавунца всасывает получающуюся переваренную кашицу через эти же каналы.
Следующая пара челюстей - нижние челюсти - у насекомых с грызущими ротовыми частями представляет членистые образования. Каждая челюсть состоит из основного членика, стволика и на вершине несет две жевательные лопастинки - внутреннюю и наружную. Часто они сливаются вместе или одна из них недоразвивается. От стволика отходит и членистый чувствующий придаток - щупик. Щупики иногда похожи по строению и функции на усики (например, у личинок жужелиц), иногда целиком редуцируются (у стрекоз, личинок сетчатокрылых).
Принципиально так же устроена и нижняя губа у насекомых, которая развивается из двух зачатков конечностей, срастающихся затем под ротовым отверстием так, что из частей, соответствующих основным членикам, образуется непарный подподбородок, а из стволиков - подбородок. Жевательные пластинки нижней губы не срастаются, а образуют язычки и придаточные язычки. Иногда нижняя губа становится основным хватательным аппаратом, выполняя такую роль, как направленные вперед мандибулы у многих хищных личинок. Это характерно для личинок стрекоз (рис. 182).
В ряде случаев ротовой аппарат сохраняет типичное для грызущего типа строение, а функционирует как сосущий, причем канал для введения в тело добычи фермента образуется не внутри мандибулы, а трубкообразным просветом между мандибулой и соответствующей максиллой. Такие каналы имеются у личинок муравьиного льва, личинок светляков.
В большинстве случаев в состав грызущего ротового аппарата входят не только 3 пары конечностей, но и прикрывающая ротовое отверстие сверху непарная складка покровов - верхняя губа, подвижно сочлененная с передним отделом головной капсулы. У личинок жуков, прокладывающих ходы направленной вперед головой, верхняя губа часто редуцирована (личинки щелкунов, жужелиц). В ротовой аппарат входят также непарные утолщенные части стенок ротовой полости - надглоточник, часто сливающийся с верхней губой, и подглоточник, подходящий к середине нижней губы.
У тех насекомых, которые добывают пищу высасывая соки из тканей растений или животных, ротовые части преобразуются в колюще-сосущий аппарат - в хоботок.
У полужесткокрылых, например, вытянутая нижняя губа превращается в трубчатый футляр со спрятанными внутри способными выдвигаться колющими щетинками, в которые превратились верхние и нижние челюсти (рис. 221). У комара колющими щетинками становятся и другие части ротового аппарата - подглоточник и верхняя губа с надглоточником (рис. 407, В). Жидкая пища (соки растений, кровь) поступает из ранки в рот насекомого по капиллярам, образованным соприкасающимися колющими щетинками, или по желобкам в них.
У бабочек, сосущих нектар (что не требует прокалывания твердых тканей), развивается хоботок, состоящий из сложенных, очень сильно вытянутых нижних челюстей. Верхние челюсти полностью исчезают, нижняя губа представлена маленькой пластинкой, от которой отходят хорошо развитые нижнегубные щупики, выполняющие функцию органов химического чувства (рис. 320). Описание всех тех преобразований, которые характерны для ротового аппарата разных насекомых, требует нескольких специальных томов. Отметим только, что иногда ротовые части с утратой потребности в них сильно или целиком редуцируются. Последнее наблюдается у личинок многих мух, живущих в том веществе, которое служит им пищей (например, личинки мясных мух Callihora в трупах животных); они разжижают пищу, отрыгивая пищеварительные соки наружу. Пища переваривается личинкой вне организма и всасывается в жидком легко усваиваемом виде. У таких личинок вместо 3 пар челюстей остается только пара ротовых крючков, помогающих при ползании. Не развиваются ротовые органы и у многих взрослых непитающихся крылатых насекомых (поденки, самцы щитовок, самцы бабочек-чехлоносок).
У всех насекомых за головой следует грудь. Иногда голова незаметно переходит в грудь, широко с ней соприкасаясь, иногда бывает отделена тонким перехватом. Большинство насекомых может двигать головой только сверху вниз или наклоняя ее в стороны, и только богомолы (Mantodea) могут поворачивать голову, как это делаем мы. Грудь состоит из передне-, средне- и заднегруди. У первичнобескрылых сегменты груди подвижны друг относительно друга, а в большинстве случаев происходит слияние средне- и заднегруди в единое целое. Развитие отдельных сегментов груди зависит от степени участия разных ее придатков в движении.
У взрослых насекомых каждый из сегментов груди несет по паре конечностей, состоящих из следующих отделов: тазик, вертлуг (у некоторых перепончатокрылых он двойной - есть еще вертлужок), бедро, голень и лапка. Число члеников лапки бывает 1-5, но не более 5. Если все пары ног развиты одинаково, конечности называются ходильными или бегательными (палочники, жуки-жужелицы, табл. 28; 39, 10). Такие конечности приспособлены к бегу по твердой поверхности. Иногда "твердая" поверхность может быть и очень зыбкой - вспомним водомерок, у которых на лапках выделяется жировой слой, препятствующий смачиванию и прорыву поверхностной пленки воды.
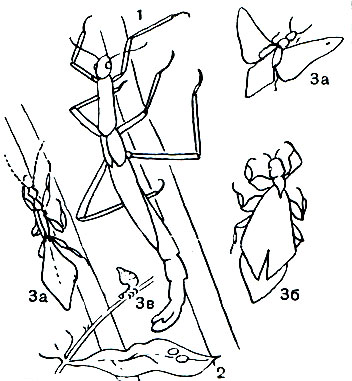
Таблица 28. Тропические палочники: 1, 2 - гигантский индонезийский палочник (Cyphocrania gigas) и его яйца; 3 - цейлонская листовидна (Phyllium crurifolium); а - самец, б - самка, в - личинка

Таблица 28. Тропические палочники: 1, 2 - гигантский индонезийский палочник (Cyphocrania gigas) и его яйца; 3 - цейлонская листовидна (Phyllium crurifolium); а - самец, б - самка, в - личинка
При обитании в траве или листве деревьев много преимуществ дает движение сильными прыжками. При прыжке наибольший эффект обеспечивает быстрое распрямление задних конечностей, толкающих тело вперед (это наблюдается и у четвероногих: лягушка, заяц, тушканчик имеют более развитые задние конечности, чем передние). У многих насекомых, передвигающихся таким образом, сильно удлиненные мощные бедра и длинные голени задних ног. Такие задние ноги, как у саранчовых, кузнечиков, жуков земляных блошек, клопов-прыгунов (Halticus), называются прыгательными (табл. 29, 31).
Жизнь взрослых насекомых в почве приводит к усилению значения передних ног при движении - к развитию роющих конечностей, характеризующихся расширением лопатообразных голеней (у медведок, у южноамериканских цилиндрохет, жуков-карапузиков и др., рис. 111).
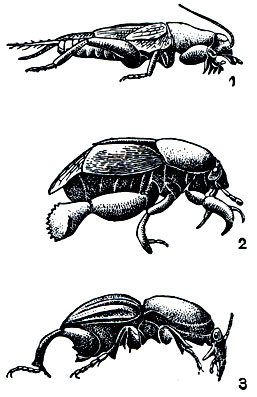
Рис. 111. Конвергентное сходство у роющих насекомых: 1 - медведка (Gryllotalpa); 2 - клоп Stibaropus; 3 - южноамериканский жук-усач Hypocephalus
У тех насекомых, которые передними ногами ловят добычу, бедра и голени передних ног удлиненные, с шипами на сторонах, сближаемых при складывании; такие ноги называются хватательными (у мантисп, богомолов, табл. 25, рис. 239).
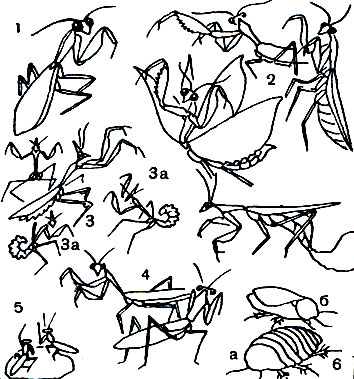
Таблица 25. Богомолы и тараканы: 1 - древесный богомол (Hierodula tenuidentata); 2 - обыкновенный богомол (Mantis religiosa); 3, 3а - рогокрылая эмпуза (Empusa pennicornis) и ее личинки; 4 - риветина (Rivetina baetica); 5 - богомол-крошка (Armene); 6 - таракан-черепашка Соссюра (Polyphaga saussurei); а - самка, б - самец

Таблица 25. Богомолы и тараканы: 1 - древесный богомол (Hierodula tenuidentata); 2 - обыкновенный богомол (Mantis religiosa); 3, 3а - рогокрылая эмпуза (Empusa pennicornis) и ее личинки; 4 - риветина (Rivetina baetica); 5 - богомол-крошка (Armene); 6 - таракан-черепашка Соссюра (Polyphaga saussurei); а - самка, б - самец
У насекомых, перешедших к жизни в воде, ноги часто бывают с удлиненными плоскими голенями, покрытыми длинными волосками (табл. 41). Такие плавательные ноги работают как весла.
Редукция ног у взрослых насекомых наблюдается при переходе к неподвижному образу жизни и может быть полной (у настоящих щитовок - Diaspinae). У личинок многих насекомых утрата конечностей бывает связана с приспособлением к червеобразному передвижению в почве и в сходных субстратах (например, у личинок двукрылых, долгоносиков и др.). У насекомых с полным превращением (жуки, сетчатокрылые), если личинки даже имеют развитые бегательные ноги, они никогда не имеют такого полного расчленения, как у взрослых форм,- часто происходит слияние голени и лапки (образуется голенелапка), лапка всегда одночлениковая и имеет один коготок (иногда двойной). Иногда ноги у личинок сохраняются только в виде одночлениковых образований (например, у зерновок).
Для высших насекомых характерна способность к активному полету с помощью крыльев. Крылья развиваются полностью только при переходе насекомого во взрослое состояние, а закладываются как плоские листовидные выпячивания покровов средне- и заднегруди. Это выпячивание разрастается, его противоположные стенки плотно сближаются, в результате чего образуется эластичная пластинка, укрепленная жилками, представляющими собой затвердевшие участки тех путей, по которым во время формирования крыла в него нагнеталась гемолимфа, расправляя крыловую пластинку. Жилки располагаются так, что становятся механической опорой крыла и помогают наилучшим образом преодолевать сопротивление воздуха при полете. В частности, у хорошо летающих насекомых передний край крыла укреплен мощной жилкой. Расположение и число жилок - признаки, хорошо характеризующие каждую систематическую группу насекомых. Строение крыльев в значительной мере кладется в основу систематики крылатых насекомых, а так как в ископаемом состоянии от насекомых лучше всего сохраняются крылья, палеоэнтомология - наука об ископаемых насекомых - особенно широко использует признаки строения крыла. В крыле различаются продольные жилки, частично ветвящиеся, и поперечные, соединяющие продольные друг с другом. Просветы перепончатой части крыла, заключенные между жилками, называются ячейками. Каждая жилка соответственно ее расположению на крыловой пластинке имеет определенное наименование, так же как и ячейки.
У более примитивных групп насекомых сеть жилок гуще, чем у представителей высших отрядов. Кроме того, у представителей высших отрядов передние и задние крылья обычно значительнее отличаются друг от друга, чем у более примитивных, у которых обе пары крыльев довольно сходны. Нередко развивается лишь одна пара крыльев, чаще передние (у мух, самцов щитовок, некоторых поденок), но бывает и наоборот - развиты только задние крылья (у веерокрылых). Нередко крылья, особенно передние, принимают на себя дополнительные функции, а часто такие дополнительные функции становятся основными, а летная функция утрачивается. Например, передние крылья у прямокрылых (саранча, кузнечик) более кожистые, чем задние; в сложенном состоянии более тонкие перепончатые задние крылья прикрыты передними, защищающими от повреждений при передвижениях в зарослях растений. У клопов защитная функция передних крыльев, превратившихся в надкрылья, выражена еще ярче: у них только самая вершина передних крыльев перепончатая (рис. 220). А для самого богатого видами отряда насекомых - жуков характерно то, что передние крылья целиком превратились в жесткие защитные надкрылья. При полете надкрылья жуков играют роль только планирующих выростов, не производящих собственных движений, летные движения совершают задние крылья. Вероятно, это наблюдали многие, кто видел, как летают божьи коровки, жуки-нарывники, жуки-олени и др. А иногда у жуков при полете надкрылья даже не расправляются - задние крылья только высовываются в стороны из-под надкрылий (у бронзовок, рис. 112).
Рис. 112. Летящая бронзовка Cetonia aurata
Нередки случаи утраты летной функции крыльев и их исчезновение. Для многих жуков, живущих в сухих местностях, характерно, что у них надкрылья полностью срастаются друг с другом, образуя защитный "шлем" над брюшком. Естественно, что со срастанием надкрылий задние крылья не могут действовать и исчезают. Это наблюдается у многих долгоносиков-скосарей, жужелиц (хлебная жужелица, рис. 251, 2) и особенно часто у степных и пустынных чернотелок.
У некоторых широко распространенных видов чернотелок можно проследить разные степени редукции крыльев. Так, например, песчаный медляк (рис. 113) во Франции местами совершает перелеты, а на нашей территории это жук нелетающий, у которого можно проследить все переходы от развитых, но бездействующих крыльев под надкрыльями до жалких, почти полностью исчезнувших рудиментов.
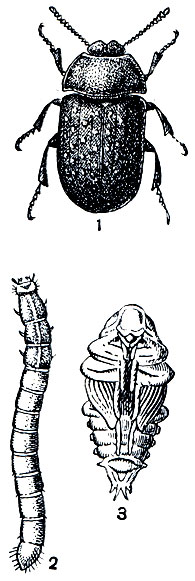
Рис. 113. Песчаный медляк (Opatrum sabulosum): 1 - взрослое насекомое; 2 - личинка; 3 - куколка
В разных группах насекомых можно столкнуться с бескрылыми формами, целиком утратившими крылья. Это относится и к тем группам, большинство представителей которых хорошо летает: среди двукрылых, бабочек, прямокрылых, клопов известны формы с исчезнувшими крыльями. В таких случаях бывает ясен факт утраты крыльев, существовавших у предков этих насекомых. И рассматривая черты внутреннего строения, и сравнивая бескрылые виды с рядом представителей той же группы, имеющих развитые и редуцированные органы полета, можно установить, что бескрылые формы произошли от крылатых предков. Такая утрата крыльев может быть вызвана разными причинами (переходом к паразитизму, к жизни в почве, при обитании на островах или в горах и т. д.).
Даже в тех случаях, когда целые отряды включают совершенно бескрылых насекомых (вши, пухоеды, блохи), изучение их строения и развития в сравнении с другими группами насекомых позволяет считать у них отсутствие крыльев следствием редукции и утраты. Вероятно, у предков, например, блох были крылья, исчезнувшие с приспособлением взрослой стадии к паразитизму.
Но среди низших насекомых есть и такие отряды, представители которых никогда и не имели крыльев: отряд щетинохвосток, например, заслуживает названия первичнобескрылых. Все особенности представителей этого отряда дают основание считать его происшедшим от далеких бескрылых предков, бывших, однако, и предками крылатых насекомых.
У насекомых, имеющих более или менее сходные передние и задние крылья, снабженные густой сетью жилок, т. е. у более примитивных (стрекозы, сетчатокрылые, термиты и др.), передние и задние крылья при полете двигаются независимо друг от друга, а у бабочек, перепончатокрылых, цикад и других более высокоорганизованных форм переднее и заднее крылья сцепляются так, что образуют единую согласованно двигающуюся поверхность. У мух, имеющих только передние развитые крылья, задние крылья превращены в небольшие булавовидные придатки, богатые чувствующими волосками. Они работают как гироскопы, служат стабилизаторами при полете. Изменение направления при полете достигается изменением положения плоскости передних крыльев (у дневных бабочек) или амплитуды и плоскости вибрации (у пчел).
Скорость вибрации крыльев у разных насекомых значительно варьирует. Бабочка махаон (табл. 43, 2) при полете делает только 5-9 взмахов крыла в секунду, пчела около 200, а комары-звонцы - 1000. Такая скорость сокращений обеспечивается резко и густо исчерченными поперечнополосатыми мышцами, на долю которых у хорошо летающих форм приходится 15-25% веса тела.
Скорость полета у насекомых очень различна: у златоглазки - 0,5 м, у пчелы - 3 м, у бабочек бражников - 15 м в секунду. Самые сильные насекомые, такие, как бражники, без посадки летят на сотни километров. Известны, например, случаи залета олеандрового бражника (Deilephila nerii), распространенного в средиземноморских странах (где растет олеандр), в Ленинград и Эстонию.
На втором - девятом брюшных сегментах только у первичнобескрылых есть одночлениковые рудименты конечностей. Кроме того, на конце брюшка у низших насекомых бывают придатки (церки), выполняющие роль органов чувств (особенно это выражено у эмбий) или хватательных органов (у уховерток). Видоизмененные зачатки конечностей сегментов рядом с половым отверстием служат вспомогательными органами при оплодотворении (гоноподнями). У низших групп насекомых самки гоноподиями подхватывают пакет с семенной жидкостью - сперматофор.
Кутикула многих насекомых покрыта различной длины волосками, иногда очень густо. Те волоски, которые представляют просто выросты кутикулы, служат главным образом для того, чтобы при затоплении насекомого (во время дождя и. т. п. ) создать вокруг тела слой воздуха, изолировать тело от соприкосновения с водой. Но кроме них, на теле насекомых много волосков и щетинок, подвижно сочлененных с ямкой в кутикуле, к основанию которых подходят окончания нервов - это основные осязательные элементы насекомого.
Нередко тело (у чешуйниц) и крылья (у жуков, бабочек и др.) бывают покрыты чешуйками. Волоски и чешуйки определяют у многих насекомых окраску. Например, у бабочек яркая пестрая окраска крыльев зависит не от присутствия какого-либо пигмента, а от явлений светового резонанса при преломлении световых лучей через чешуйки - сами чешуйки бесцветны. От структуры кутикулы зависит и металлическая окраска жуков, перепончатокрылых и других насекомых. Такая окраска, зависящая не от пигмента, а от структуры покровов, называется оптической. Она была расшифрована в 1902 году киевским физиком И. И. Косовоговым. Наряду с оптическими явлениями окраску многих насекомых определяют и отлагаемые клетками гиподермы пигменты. Черная окраска чаще всего зависит от отложений меланина. Распространенная у насекомых зеленая окраска (у саранчовых, палочников и др.) долгое время рассматривалась как связанная с хлорофиллом растений, но, по новым данным, она определяется особыми пигментами.
Окраска многих насекомых, особенно определяемая преломлением лучей в хитиновых частях покровов, сохраняется и у мертвых высушенных экземпляров. Так, бабочки, собранные и наколотые уже более 200 лет тому назад К. Линнеем, сохраняют такую же свежесть и яркость тонов, как живые. Прекрасно сохраняют металлический блеск и многие жуки, например бронзовки. А яркая окраска надкрылий божьих коровок, прямокрылых и других насекомых, у которых она зависит от органических пигментов, у собранных высушенных экземпляров тускнеет и выцветает. Выцветает иногда и металлическая окраска, например у жука щитоноски (Gassida nobilis).
Изумительно многообразие расцветок насекомых. Многие из них по окраске могут соперничать с пестрыми и яркими цветами, а многие имеют тусклую неяркую окраску, делающую их непривлекательными и незаметными. Чем же объясняется такое многообразие окрасок насекомых?
Окраска имеет огромное биологическое значение. Не защищенные другими способами насекомые, если их окраска напоминает тот фон, на котором они держатся, становятся незаметными для истребляющих их хищников, в первую очередь для птиц. Каждый собиратель насекомых знает, как трудно бывает обнаружить сидящую на вбитом в землю потемневшем деревянном столбике озимую совку, как незаметны бывают сидящие на покрытом лишайниками стволе дерева бабочки пяденицы, как легко пройти мимо зеленой гусеницы, сидящей на листе, как ускользает из поля зрения прыгающая на поверхности серой почвы серая саранча и т. п.
Итальянский ученый Чеснола делал такие опыты. Он собирал крупных богомолов, имеющих сероватую или зеленую окраску. Сероватые незаметны на фоне сухой травы, где они держатся, а зеленые - на зеленой. Он привязывал равное количество богомолов разной окраски к травинкам такого же цвета или к травинкам другого (половину сероватых к сухой траве, половину к зеленой, половину зеленых к сухой, половину к зеленой). Оказалось, что количество склеванных птицами богомолов было больше в тех случаях, когда они были привязаны к травинкам другого цвета, что больше выжило тех, которые были на "своем цвете". Опыты Чеснолы у нас были проверены М. М. Беляевым. Окраска, делающая насекомое незаметным на окружающем фоне, называется покровительственной (табл. 16).
Металлически блестящие места покровов насекомых в естественной обстановке отражают преобладающий окружающий фон и таким образом служат тоже своеобразной защитной окраской. Часто покровительственной бывает не только окраска, но и форма тела - у тех же богомолов вытянутое тело напоминает стебелек травинки. Еще ярче выражена покровительственная окраска и форма у палочников, у гусениц пядениц, тело которых часто похоже на сучок, и т. д.
Интересна так называемая криптическая окраска у насекомых. Бывает, что причудливый узор, например на крыльях бабочки, не привлекает внимания хищника, а, наоборот, делает насекомое незаметным. Так, глазчатые пятна на крыльях большого ночного павлиньего глаза делают крылья похожими на поверхность коры с углублениями - рисунок имитирует, если смотреть издалека, неровности поверхности, создает стереоскопический эффект. Такие примеры "стереоморфизма" изучал ленинградский профессор Б. Н. Шванвич.
Но бывает, что окраска насекомого как бы нарочно привлекает внимание. В таких случаях насекомое обычно надежно защищено от хищников отталкивающим запахом или вкусом, ядовитыми железами и т. п. Так, яркую окраску имеют многие клопы, например солдатики; жуки-нарывники, божьи коровки выделяют едкие вещества, например кантаридин. Ярко окрашены и защищены ядовитым жалом осы, например, шершни. Такая яркая окраска носит название предупреждающей (табл. 16). Действительно, замечено, что птицы научаются не трогать таких защищенных насекомых. Следует, правда, иметь в виду, что нередко окраска, воспринимаемая нами как пестрая, для хищника, истребляющего данный вид насекомого, может представляться однотонной и быть защитной. Известно, что способность различать цвета неодинакова у разных видов животных и, например, собака цвета не различает. Способность же разных видов птиц различать цвета специально не изучалась.
Большое распространение в мире насекомых имеет так называемая мимикрия (табл. 17) - сходство очертании и окраски двух видов, из которых один характеризуется предупреждающей окраской и несъедобен или опасен для преследователей, а другой, незащищенный, имитирует защищенный. Неопытному наблюдателю бывает трудно отличить мух-сирфид от пчел и ос - настолько совершенно и расцветка, и характер движений этих беззащитных мух имитируют жалоносных ядовитых перепончатокрылых. Неискушенный собиратель насекомых нередко опасается взять в руки сирфиду, думая, что имеет дело с пчелой, осой или шмелем. Похожи на ос и бабочки-стеклянницы. Очень интересен пример мимикрии южноамериканских бабочек-белянок, "подражающих" ядовитым бабочкам-геликонидам, около 100 лет тому назад описанный Бэтсом (подробнее см. выше).
Во всех таких случаях незащищенное насекомое меньше истребляется птицами, ящерицами и другими врагами, так как они принимают его за ядовитую "модель".
Иногда, конечно, окраска имеет и прямое приспособительное значение. Так, жуки-чернотелки в пустынях характеризуются либо белой окраской (парадокс - чернотелки, а белые!), отражающей лучи солнца, либо чаще (отсюда и название семейства) интенсивно черной - плотные черные покровы задерживают губительные короткие лучи спектра.

Рис. 114. Скопление божьих коровок на зимовке
На поверхности тела у насекомых особыми канальцами открываются различные железы, выделяющие пахучие вещества, иногда воспринимаемые только другими насекомыми, иногда доступные и нашему обонянию (например, у различных клопов, жуков-медляков рода Blaps и т. д.). Часто пахучие выделения помогают особям одного вида находить друг друга (выделения самок привлекают самцов, запахи залегающих на зимовку клопов-черепашек или божьих коровок обеспечивают скопление зимующих особей и т. д.). Некоторые выделения желез имеют пищевую привлекательность для особей того же вида (например, выделения шейных желез самцов кузнечиков слизываются самками) или для других видов (выделения жуков, обитающих в муравейниках, охотно слизываются муравьями). Иногда железы выделяют ядовитые вещества. Поступающие в полые волоски ядовитые выделения служат средством защиты - известно, что ядовитые волоски многих гусениц, например златогузки, могут вызывать воспалительные процессы кожи человека. Едкие защитные вещества выделяют многие жуки (кантаридин - жуки-нарывники, педерин - некоторые стафилиниды). Такие яды могут вызывать явления воспаления и омертвения покровов, если попадают на кожу. Нередки у насекомых железы, выделяющие воск и воскоподобные вещества, часто служащие насекомым для защиты от потери воды (щитки у щитовок, восковые нити у червецов и подушечниц и т. д.).
Пищеварительная система у насекомых, как и у всех членистоногих, состоит из передней, средней и задней кишки (рис. 115).
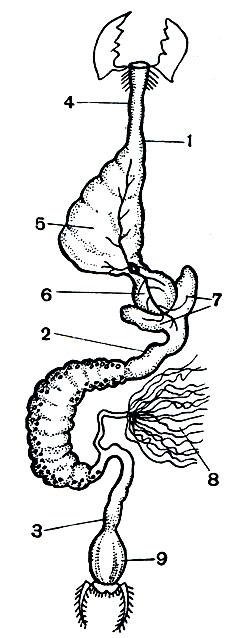
Рис. 115. Пищеварительная система сверчка: 1 - передняя кишка; 2 - средняя кишка; 3 - задняя кишка; 4 - пищевод; 5 - зоб; 6 - железистая часть желудка; 7 - слепые выросты средней кишки; 8 - мальпигиевы сосуды; 9 - прямая кишка
Передняя кишка начинается ротовой полостью, в которую впадают слюнные железы. Слюнные железы у разных насекомых вырабатывают разные ферменты. У растительноядных это чаще всего ферменты, переводящие дисахариды в моносахариды (инвертазы). У хищников, например у водных клопов, в слюне содержится фермент, расщепляющий белки на аминокислоты (протеаза). У кровососов часто в слюне есть фермент, препятствующий свертыванию крови (например, у малярийного комара).
На нижней губе у личинок бабочек, ручейников и ряда других насекомых открываются железы, секрет которых быстро затвердевает; это шелкоотделительные железки, продукт которых употребляется для изготовления различных защитных (коконы) или ловчих (тенета ручейников) образований. Трубчатый пищевод в задней части нередко расширяется в зоб, иногда приобретающий вид мешка, особенно развитый у тех насекомых, которые питаются жидкой пищей.
Средняя кишка является тем местом, где в основном переваривается пища. Железистые стенки среднего отдела кишечника (его расширенная часть называется желудком) вырабатывают ферменты, которые расщепляют и разжижают служащие пищей вещества.
Кроме ферментов (вырабатывающихся и у нас), расщепляющих белки, жиры, крахмал и сахар, многие насекомые соответственно роду пищи вырабатывают и другие энзимы. Многие личинки жуков-дровосеков вырабатывают целлюлазу, разрушающую клетчатку, личинки кожеедов выделяют кератиназу, действующую на роговые вещества, и т. п. Очень часто такие трудноразрушаемые соединения, как клетчатка, разрушаются с помощью симбиотических бактерий и простейших. Известно, что термиты потребляют в основном клетчатку, но своих разлагающих ее ферментов не выделяют - у них в кишечнике много симбиотических многожгутиковых (Trichonympha и др.), усваивающих целлюлозу. У тех насекомых, которые используют кишечную микрофлору для переваривания пищи, кишка имеет большие слепые выросты или достигает очень большой длины и пища задерживается в кишечнике на длительные сроки (например, у личинок майского хруща). Недавно было выяснено, что те насекомые, которые питаются бедными белками соками растений (например, тли), усваивают азот воздуха с помощью симбиотических азотфиксирующих бактерий.
В средней кишке пищевая масса обычно одевается особой легкопроницаемой белковой оболочкой, защищающей нежные клетки кишки от повреждений. Оболочка выделяется самым передним участком средней кишки. Средняя кишка не имеет кутикулы, а задняя, как и пищевод, покрыта проницаемой кутикулой. В задней кишке происходит всасывание переваренной пищи и, главное, всасывание воды. Расположенные в конце пищеварительного тракта у многих насекомых так называемые "ректальные железы" служат для интенсивного всасывания воды.
Выделительными органами у насекомых служат мальпигиевы сосуды - тонкие трубчатые выросты кишечника на границе средней и задней кишки (рис. 115). Число их варьирует от 2 до 150, их свободные слепые концы лежат в полости тела, не сообщаясь с нею. Растворимые продукты обмена поглощаются стенками мальпигиевых сосудов и по их просветам проходят в заднюю кишку, обычно еще в них переходя в мелкие кристаллики мочевой кислоты. У тех более примитивных крылатых насекомых, которые живут во влажной среде, особенно у водных насекомых (стрекозы, поденки), или у тех, которые потребляют много сочной растительной пищи (прямокрылые), мальпигиевых трубочек много - они принимают на себя роль осморегуляторных образований, выводящих избыток воды. Слепые концы мальпигиевых трубочек у насекомых, живущих в сухих условиях,- у жуков и личинок чернотелок, гусениц, личинок муравьиных львов - прирастают к стенке задней кишки ниже того места, где эти трубочки впадают в кишечник. Просвета в месте присоединения нет, поглощение воды концом мальпигиева сосуда из задней кишки происходит путем диффузии. Этим обеспечивается непрерывное промывание мальпигиева сосуда одним и тем же небольшим объемом воды (рис. 116).
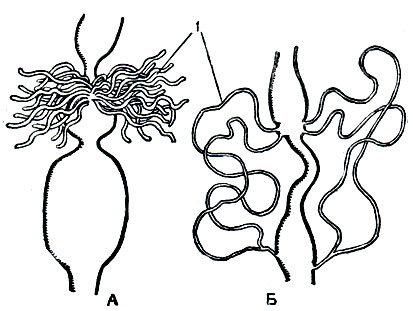
Рис. 116. Выделительная система насекомых: А - стрекозы; Б - жука-листоеда; 1 - мальпигиевы сосуды
У некоторых первичнобескрылых насекомых (Machilis) наряду с мальпигиевыми трубочками некоторую роль в выделении играют открывающиеся на нижней губе железы - органы выделения, напоминающие максиллярные железы ракообразных.
Основная дыхательная система у насекомых - трахеи. У живущих в воде, во влажной почве и в тканях растений личинок насекомых большую роль играет также кожное дыхание, от которого и произошло трахейное дыхание.
У некоторых двукрылых вышедшая из яйца личинка еще не имеет трахей, которые развиваются только после линьки. Но чаще кожное дыхание существует наряду с трахейным, или, когда насекомые вторично переходят к кожному дыханию, у них сохраняется хорошо развитая система трахей, не имеющих сообщения с внешней средой.
Трахейная система у насекомых представляет сообщающуюся систему трахейных трубочек, открывающихся по бокам тела отверстиями - дыхальцами. Только у махилисов (Machilis) пучки трахей не сообщаются друг с другом. Наибольшее число дыхалец у насекомых - 10 пар (по паре на первых 8 брюшных сегментах, 2 пары на грудных), но количество действующих дыхалец (у личинок насекомых, живущих в воде, во влажной земле и т. п.) может уменьшаться до 1 пары.
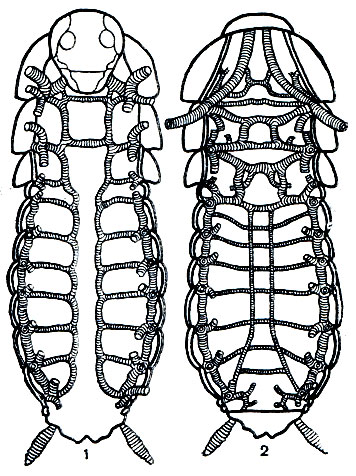
Рис. 117. Трахейная система таракана: 1 - брюшные, 2 - спинные трахейные стволы
У насекомых, как и у других наземных членистоногих, кислород попадает от дыхалец к клеткам тела путем диффузии по просветам трахей и их тончайших веточек - трахеол. Но у многих высших насекомых имеется система вентиляции, проталкивания воздуха через крупные стволы трахей, что ускоряет газообмен. При движениях брюшка то расширяются, то сжимаются широкие трахейные стволы или связанные с ними воздушные мешки. А определенное направление тока воздуха в трахеях при таких дыхательных движениях обеспечено тем, что у таких насекомых дыхальца снабжены клапанами, то закрывающимися, то открывающимися. У тех личинок насекомых, которые живут в воде, во влажной земле и в других влажных средах, дыхальца всегда открыты для воздуха. Хорошо приспособившиеся к жизни в воде личинки поденок, веснянок, ручейников и других насекомых не имеют открытых дыхалец. Кислород у них проникает через поверхность всех участков тела, где покровы достаточно тонки, особенно через поверхность листовидных выростов, пронизанных сетью слепо заканчивающихся трахей. У таких личинок хорошо развита трахейная система, но механизм дыхания сложный - кислород проникает через покровы, растворяется в полостной жидкости, затем из полостной жидкости диффундирует в трахеи, затем уже по трахеям доходит до всех внутренних органов.
Многие живущие в воде насекомые (водные жуки и клопы, личинки и куколки комаров и др.) должны время от времени подниматься к поверхности, чтобы захватить воздуха, т. е. у них дыхание воздушное. Личинки комаров, водные личинки долгоножек и др. на время обновления запаса воздуха в своей трахейной системе подвешиваются с помощью венчиков несмачиваемых жирных волосков к поверхностной пленке воды. Водные жуки - водолюбы, плавунцы,- и клопы, например гладыши, обновив запас воздуха у поверхности, уносят его с собой вглубь- воздух уносится под надкрыльями и в виде слоя, приставшего к несмачиваемым волоскам на поверхности тела.
Оказалось, что, находясь под водой, такие насекомые тратят гораздо больше кислорода, чем содержится в запасе, унесенном ими из атмосферы. Дело в том, что между содержанием кислорода в соприкасающемся с водой слое воздуха на теле нырнувшего насекомого и окружающей водой устанавливается некоторое равновесие: по мере того как потребляется кислород из этого слоя, в него проникает кислород, находящийся в растворенном виде в окружающей воде!
Таким образом, слой воздуха на теле водного насекомого играет роль как бы особого приспособления к дыханию под водой (как водолазный колокол) и даже называется "физической жаброй". Вспомним, что несмачиваемые волоски на теле наземных насекомых имеют такое же значение в случае, например, затопления талыми водами в конце зимовки в почве: вокруг тела насекомого сохраняется слой воздуха - "физическая жабра".
У многих насекомых, например у личинок комаров-мотылей (Chironomus), нет трахейных жабр, но на заднем конце тела есть отростки с тонкими покровами, которые считались тоже жабрами ("кровяные жабры"). А оказалось, что эти отростки - места поглощения солей из воды, а не жабры; дыхание у личинок мотылей кожное, всей поверхностью тела. Тело их кажется красным от просвечивающего гемоглобина. Но гемоглобин у таких личинок служит не для переноса кислорода, а для более прочного его связывания и запасания. Это бывает важно для живущих в илистом грунте личинок - там часто создаются анаэробные условия.
Кровяная жидкость - гемолимфа, циркулирующая в полости тела насекомых,- бесцветная. Гемолимфа не имеет большого значения для снабжения всех тканей кислородом - она служит для транспортировки растворенных в ней поступающих из кишечника питательных веществ, для выноса из клеток продуктов обмена, для распределения в теле гормонов. В гемолимфе есть клетки-фагоциты, помогающие насекомым справляться с попадающими в их тело микроорганизмами, а иногда и с более крупными паразитами. Фагоциты окружают и часто "инкапсулируют" паразита, попавшего в тело насекомого, что вызывает его гибель.
Циркуляция гемолимфы обеспечивается чередующимися расширениями и сокращениями трубчатого спинного сосуда-сердца. При растягивании стенок сосуда, осуществляемого крыловидными мышцами (рис. 118), гемолимфа, обогащенная питательными веществами, через боковые отверстия (остии) попадает в сердце, а при сокращении стенок сосуда клапаны в остиях закрываются и гемолимфа выталкивается через аорту к головному мозгу.
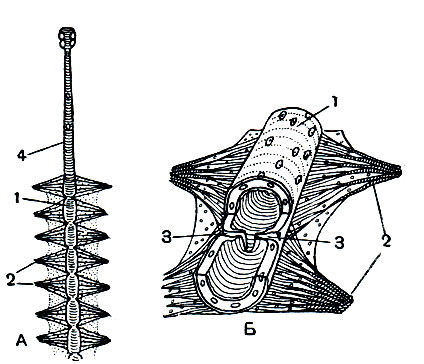
Рис. 118. Кровеносная система насекомого: А - спинной сосуд (сердце) сверху; Б - участок спинного сосуда в разрезе; 1 - сосуд; 2 - крыловидные мышцы; 3 - остии; 4 - аорта
Нервная система у насекомых построена сходно с нервной системой низших членистоногих и даже кольчатых червей: есть головной мозг - надглоточные скопления нервных клеток - и брюшная нервная цепочка. Большого развития и усложнения достигает у насекомых головной мозг. Брюшная же цепочка состоит из подглоточного узла и обычно 10 грудных и брюшных ганглиев, причем соседние ганглии брюшной цепочки часто сближаются и полностью сливаются друг с другом. В головном мозге, всегда представляющем слитное образование, различают 3 части: "первичный" мозг, обычно самый большой отдел, связанный с органами зрения, "вторичный", связанный с усиками, и "третичный", дающий ветви к верхней губе и передней части кишечника. В мозге развиты в основном ассоциативные клетки, к которым подходят нервы от органов чувств. В центре "первичного" мозга есть "грибовидные тела", состоящие из скоплений ассоциативных клеток.
Чем сложнее поведение насекомого, тем сильнее у него развит головной мозг.
У пчелы он составляет 1/174 объема тела, а у жука-плавунца всего 1/420. У маленькой пчелы мозг даже абсолютно больше, чем у крупного жука-оленя (рис. 119).
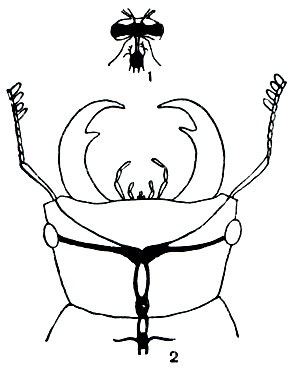
Рис. 119. Головной мозг пчелы (Apis mellifera) (1) и американского жука-оленя (Lucanus dama) (2)
Чем многообразнее раздражения, получаемые насекомыми, чем детальнее насекомое способно их различать и чем совершеннее возможности отвечать на раздражение многообразными движениями, тем сложнее поведение. Особенно сложным бывает поведение многих насекомых в период размножения. Какие же раздражения ощущаются насекомыми? Они очень многообразны. У большинства ведущими органами чувств оказываются органы зрения, о чем можно судить и на основании особо сильного развития "первичного" мозга (табл. 18, 19).

Таблица 18а. Вид бабочки, воспринимаемый глазом человека
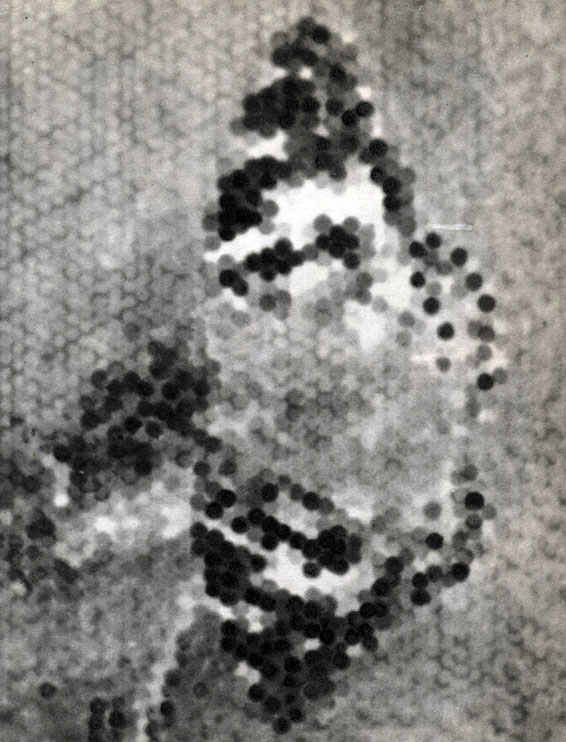
Таблица 18б. Вид бабочки, воспринимаемый фасеточным глазом насекомого

Таблица 18в. Вид цветка мака, воспринимаемый глазом человека

Таблица 18г. Вид цветка мака, воспринимаемый фасеточным глазом насекомого

Таблица 19. Зрительное восприятие человека и насекомых: справа - рисунок крыльев бабочек, цветков и листьев растений, видимый человеком; слева - те же объекты, видимые насекомым, воспринимающим ультрафиолетовые лучи
Конечно, не у всех насекомых глаза развиты одинаково, очень немногие виды подвергались изучению, но те материалы, которые есть, позволяют отметить, что насекомые (личинка стрекозы, пчела) воспринимают смену изображений как мелькание при несколько даже большей частоте, чем воспринимаем мы. Сложные глаза дают достаточно четкую картину близких предметов и позволяют различать силуэты отдаленных. Простые глазки, а у многих бабочек и сложные глаза хорошо воспринимают изменение интенсивности освещения. Для многих насекомых доказана способность различать цвета. Эта способность не всегда одинакова, но экспериментально показано, что тли распознают длинноволновую и коротковолновую части спектра (красный - зеленый участок отличают от синего-фиолетового), что шведскую мушку привлекают голубые оттенки на зеленом фоне, что муравьи различают ультрафиолетовые лучи, а пчелы хорошо различают четыре цвета, но иные, чем различаем мы. Один цвет для пчел - это красный, желтый и зеленый; в этом участке спектра пчелы, выражаясь нашими понятиями, дальтоники. Другой цвет - сине-зеленый, третий - сине-фиолетовый, а четвертый - недоступный нам ультрафиолетовый!
Удивительная способность многих членистоногих различать поляризованный свет и воспринимать плоскость поляризации хорошо показана на насекомых: в глазу мух есть структуры, примерно так же действующие, как в полярископе Николевы призмы! Выяснилось, что глаз насекомого легче различает двигающиеся предметы, чем неподвижные, что запоминание пчелой летка ее улья связано с видом не только улья и самого летка, но и с восприятием всего ландшафта. Если на земле передвинуть улей на 2 м, возвращающиеся в него пчелы испытывают замешательство, стремясь туда, где леток был до перестановки улья. Но если держать улей на плоту на озере и переплыть на километр, пчелы легко находят свой улей, так как на фоне водной глади плот с ульем - единственный ориентир.
Не так давно выяснили, что пчелы могут различать и форму предметов, причем легче они различают фигуры с изрезанными, как у цветка или снежинок, краями (сходные с венчиками цветов), а труднее, но все же различают геометрические фигуры, на распознавание которых пчел можно выдрессировать.
Реакции насекомых на зрительные раздражения очень быстрые - ориентируясь по виду летящего насекомого, стрекоза меняет направление полета и схватывает добычу на лету.
Еще не все выяснено в области изучения зрения насекомых, но и то, что известно, поразительно.
Зрение не единственный путь получения информации насекомыми.
Все насекомые очень чувствительны к изменениям температуры, влияние которой на активность, длительность развития и длительность жизни насекомых исключительно велико. Каким образом насекомые, одетые в плотный панцирь, быстро воспринимают изменения температуры? Оказалось, что у насекомых есть особые теплочувствительные участки на поверхности тела, например у перелетной саранчи у основания усиков есть места с особо тонкой кутикулой, подстилаемой чувствительными клетками (рис. 120). У многих такими органами оказываются особые короткие толстостенные волоски. Такие волоски на лапке таракана воспринимают изменение температуры поверхности, по которой он ходит, в 1° - точность, труднодоступная нашему осязанию.
Рас. 120. Температурный орган на усике саранчи (в разрезе): 1 - экзокутикула; 2 - чувствующие клетки; 3 - полость органа; 4 - жировое тело
Живущие в воде насекомые хорошо реагируют на изменение давления, по величине которого они получают информацию о той глубине, на которой находятся, а также определяют положение тела. Например, у клопа, называемого водяным скорпионом (Nepa cinerea, табл. 38, 4), дыхальца на 3 сегментах ведут в камеру, выстланную очень тонкой кутикулой (мембраной), под которой лежат чувствительные клетки. В горизонтальном положении тела в воде мембраны на всех сегментах испытывают одинаковое давление, а при изменении положения на одни мембраны давление усиливается, а на другие ослабляется. Эта разница (равная всего 0,00015 ат!) уже улавливается водным клопом! Очень часто у насекомых бывают развиты органы слуха. Следует только оговориться, что слух у насекомых - это способность к восприятию не только звуковых колебаний, воспринимаемых нами, но и любых колебаний среды.
Рис. 121. Схема строения органа равновесия у водяного скорпиона (Nepa cinerea): 1 - при горизонтальном положении; 2, 3 - при наклоне налево и направо
Насекомые - единственные беспозвоночные, для которых точно доказана способность различать звуки. Классическое доказательство-опыты со сверчками, когда в одном помещении перед микрофоном сажали стрекочущего самца, а в другом - самку перед телефоном. Когда включали микрофон, самка устремлялась к телефону. Не надо только думать, что органы слуха у насекомых похожи на наше ухо. Наше ухо воспринимает изменение давления воздуха, вызываемое источником звукового колебания, а у насекомых чаще воспринимается движение воздуха.
Наиболее соответствуют нашему уху тимпанальные органы, состоящие из тонких перепонок, натянутых на склеротизованные кольца (вроде нашей барабанной перепонки), под которыми находятся пузыревидно расширенные участки трахей с подходящими к ним нервами. Такие органы есть у кузнечиков и сверчков на голенях передних ног, у саранчовых на боках первого брюшного сегмента (рис. 158), у совок на боках заднегруди, у огневок на первом брюшном сегменте, у цикад на втором и т. д.
Проще построенные хордотональные органы - натянутые эластичные волокна, к которым подходят нервные окончания,- видимо, есть у всех насекомых. Интересно, что по опытам, проведенным над несколькими видами бабочек совок, оказалось, что они реагируют на многие звуковые колебания (бабочки, улавливая звук, ускоряют полет и меняют его направление или "притворяются мертвыми"). Частота колебаний, на которую они реагируют, от 15 000 до 175 000 в секунду, причем сильнее всего реакция на колебания порядка 30 000-80 000, т. е. такого порядка, как частота ультразвуков, испускаемых летучими мышами. Известно теперь, что ориентировка летучих мышеи при полете происходит по принципу радара: они испускают ультразвуки и улавливают их отражение от летящих насекомых. И у ночных бабочек выработалась защитная реакция - изменение полета - в ответ на эти "радарные лучи" их врагов. А некоторые ночные бабочки и сами издают ультразвуки, помогающие распознавать друг друга.
У насекомых органами слуха служат и специальные волоски, располагающиеся на всей поверхности тела или на отдельных участках. Точными опытами установлено, что, хотя у сверчка основные слуховые органы - тимпанальные органы находятся на голенях передних ног, ослабленный слух сохраняется у него и после их удаления. Волоски, расположенные на церках прямокрылых, улавливают звуки частотой 50-400 колебаний в секунду, а при синхронном колебании - до 800.
Подвижно причлененные волоски - это и органы для восприятия направления ветра, тока воздуха. У основания усиков крылатых насекомых есть сложно устроенные органы чувств - джонстоновы органы, с помощью которых насекомые контролируют скорость и направление полета. Те группы насекомых, у которых наиболее хорошо развиты слух и органы слуха, способны и издавать звуки. Это относится к прямокрылым, у которых звуки издаются путем трения переднего края заднего крыла о нижнюю поверхность переднего крыла или о внутренний край бедер задних ног (у саранчовых) либо путем трения особых участков передних крыльев друг о друга (у сверчков, кузнечиков, рис. 149), к цикадам, у которых вибрирует особая мембрана, и т. д.
Явственно выражены сходные по строению "стридуляционные" органы у многих насекомых, например у личинок жуков (жуков-оленей, навозников, пассалид - рис. 248), которые живут в небольших объемах пищи. Их звуков мы не слышим, но они явно предупреждают друг друга о взаимно опасном сближении! До сих пор никто не изучал издаваемые ими не слышимые нами звуки. Следует учесть, что часто степень слышимости разных звуков неодинакова. Довольно много людей в общем с нормальным слухом, которые не слышат стрекота кузнечиков; правда, и кузнечики, вероятно, не слышат орудийных выстрелов, так как их стрекот при стрельбе не прекращается.
При восприятии звуков волосками трудно провести грань между слухом и осязанием. Многие насекомые воспринимают колебания той поверхности, на которой сидят. Полевой сверчок при частоте колебаний около 1500 в секунду воспринимает их даже при амплитуде в 0,1 миллимикрона (миллимикрон - одна миллионная миллиметра!). А садовая жужелица (Carabus hortensis) воспринимает колебания частоты 200-400 в секунду при амплитуде начиная с 5500 миллимикрон! Органы осязания насекомых - волоски, особенно чувствительные волоски на усиках и церках.
Исключительно сильно развиты у насекомых органы химического чувства. В большинстве случаев они разбросаны по всему телу (тонкие полые волоски, в полость которых заходят окончания чувствующих клеток), но основная их масса сосредоточена на усиках и щупиках. Чувствительность насекомых к запахам много выше, чем наша, например пчелы обнаруживают метилгептанон при концентрации в 40 раз меньшей, чем та, при которой его начинаем чувствовать мы.
Живущие в земле проволочники имеют обоняние в 10 раз более острое, чем наше, даже в отношении тех веществ, запах которых мы легко обнаруживаем. Именно по запаху обнаруживает и находит пищу большинство насекомых. Меченые самцы бабочки Actias selene прилетали на запах самки за 11 км, а самцы непарного шелкопряда - за 3,8 км.
Растворенные вещества могут восприниматься насекомыми не только ротовыми частями, но и лапками. Именно лапками передних ног мухи и бабочки могут пробовать, сладок ли раствор. При этом оказывается, что бабочки "ногами" чувствуют концентрации сахара в воде, в 2000 раз меньшие, чем те, начиная с которых мы распознаем сладковатый привкус! Выяснено, что насекомые могут различать сладкое, соленое, горькое и кислое. У большого водолюба, например, органы чувств на нижнечелюстных щупиках различают сладкий, горький и соленый вкус, а для распознавания кислого служат нервные окончания на вершине нижнегубных. Многие живущие в почве насекомые ориентируются по концентрации растворенных в почвенной влаге веществ, а недавно было показано, что они воспринимают и ничтожные изменения концентрации углекислоты: выделение углекислоты корнями растений привлекает личинок хрущей, проволочников и других живущих в почве вредителей.
На усиках находятся и удивительные по чувствительности органы восприятия влажности воздуха. Это небольшие бугорки и ямки (сенсиллы), которые позволяют, например, личинкам щелкунов (проволочникам) различать 0,5% относительной влажности воздуха (100% и 99,5%). Такие органы имеются и на щупиках. Неясно, каким образом некоторые насекомые, например термиты, могут воспринимать магнитное поле, но индийские энтомологи установили, что самки термитов, находящиеся в подземных гнездах, располагаются в гнезде так, что ось тела проходит по магнитному меридиану.
Насекомые улавливают и воздействие электростатического поля, что также пока не может быть объяснено. А. Ф. Лебедев полагал, что основная функция покрова волосков и выростов на поверхности тела насекомого - отведение электрического заряда.
Приведенный краткий обзор показывает, что насекомые имеют очень развитую систему органов чувств - анализаторов. Поведение насекомых носит ярко выраженный рефлекторный характер: на раздражение, воспринимаемое тем или иным органом чувств (или их комплексом), насекомое реагирует теми или иными движениями, из совокупности которых формируется поведение насекомых. Иногда поведение имеет характер ярко выраженных реакций на раздражение - характер таксисов. Так, например, живущие в почве личинки насекомых стремятся с поверхности уйти вглубь (положительный геотаксис); активные днем насекомые, попав в темное помещение, стремятся к окну, к свету (положительный фототаксис); многие скрытоживущие насекомые, например чешуйницы, уходят от света (отрицательный фототаксис) и т. п. Часто в зависимости от условий, от физиологического состояния и т. д. меняется реакция на раздражитель. Чернотелки пимелии на севере ареала в нашем полушарии дневные, а на юге ночные. Чернотелки блапсы при более низкой температуре проявляют положительный фототаксис, а при более высокой - отрицательный. Мухи формии (Phormia) привлекаются альдегидом изовалериановой кислоты при концентрации его менее 1:50 000, а при более высокой - отпугиваются им.
Очень часто определенный раздражитель является сигналом для начала той или иной активности насекомого. Например, перед заходом солнца снижение освещенности до определенной степени интенсивности является тем сигналом, по которому начинается лёт многих хрущей. В Средней Азии, где в мае часты безоблачные дни, можно с точностью до минуты предсказать, когда начнется лёт июньского хруща.
Реакции типа тропизмов у насекомых выражены недостаточно четко; сильнее выражены так называемые кинезисы. Как, например, происходит выбор насекомыми места с определенной температурой? Обычно насекомое не просто переходит, допустим, из более жаркого в более прохладное место. Находясь там, где температура выше оптимальной, насекомое быстро и беспорядочно бегает, пока не наткнется на место с более благоприятной температурой, где беспорядочные движения становятся замедленными. Если случайно насекомое выходит за пределы такой благоприятной зоны, оно снова начинает беспокойно передвигаться, пока не попадет в благоприятную зону, где постепенно замедляет движение и наконец, попав в наиболее благоприятную обстановку, останавливается. Итог - "выбрано" наиболее благоприятное место, но не путем прямого к нему устремления, а методом многочисленных "проб и ошибок".
Часто действия насекомого, которые кажутся простыми, представляют сложную цепь закономерно меняющихся друг за другом действий. Например, при движении жука-короеда к пригодному для откладки яиц дереву должно смениться в совершенно определенной последовательности не менее 7 реакций на различные раздражители (температура, влажность, цвет, запах и др.). Не только внешние, но и внутренние причины - состояние насекомого - определяют поведение. Самка жука, сегодня летающая, завтра, после оплодотворения, в точно таких же условиях среды не летает. Рабочая пчела, которая вчера не оставляла улей, несмотря на летную погоду, сегодня начинает летать за взятком.
Поведение насекомых носит характер в основном наследственно закрепленных инстинктов, представляющих определенную очень длинную и сложную цепь безусловных рефлексов. Инстинктивная деятельность многих насекомых (жуков-навозников, пчел, термитов) очень сложная, их поведение соответствует обычным условиям их жизни и производит впечатление "разумности". Однако такое наследственно закрепившееся поведение при изменении условий становится часто нецелесообразным и насекомое или его потомство погибает. Много примеров этому привел замечательный наблюдатель природы французский энтомолог Ж. А. Фабр, очерки которого ("Жизнь насекомых") следует прочесть каждому, кто интересуется насекомыми.
Однако после работ Фабра долгое время недооценивалась роль индивидуального опыта, возможность выработки условных рефлексов у насекомых. При работах с пчелами выяснилась возможность выработки у них условных связей, "обучения" пчел. Оказалось возможным при дрессировке пчел научить их распознавать цвета и некоторые геометрические фигуры, брать пищу из сосудов определенной формы и цвета.
Кроме того, оказалось, что пчелы могут, общаясь, передавать друг другу индивидуальный опыт. Если научить пчелу узнавать чашки с сахарным сиропом определенной формы и цвета, а затем поместить эти чашки в районе облета пчелы, пчела, прилетев в улей, совершая движения определенным образом, указывает другим пчелам своего улья направление, по которому нужно лететь за пищей (рис. 122).
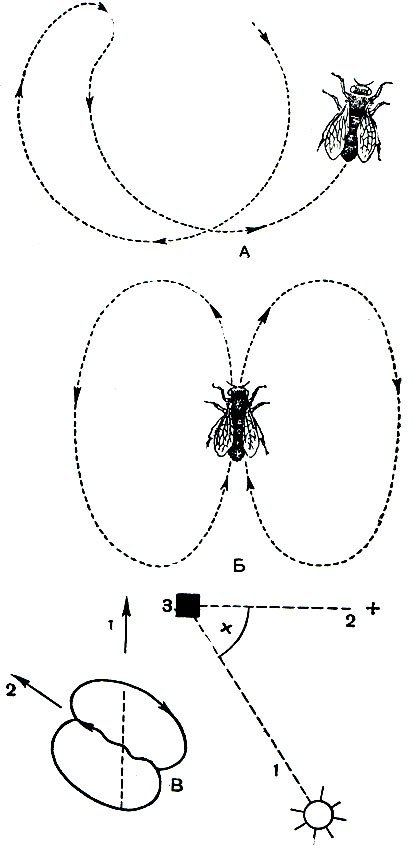
Рис. 122. Танец пчелы: А - круговой; Б - восьмерочный; В - направленный; 1 - направление к солнцу; 2 - направление к цветам; 3 - улей
Некоторые исследователи, например у нас П. И. Мариковский, изучая поведение муравьев, нашли у них целый ряд жестов, которые служат сигналами, побуждающими других особей к определенному поведению ("дай пищи", "опасность" и т. п.).
За последнее десятилетие были проведены исследования, показавшие, что у многих насекомых, особенно у общественных (муравьи, термиты и др.), выделяются биологически активные вещества, известные под общим названием "феромоны", которые влияют на развитие других особей и определяют их поведение. Так, например, насекомые могут оставлять определенные следы, по которым двигаются другие особи того же вида. Выделения матки пчелы в небольшом улье тормозят поведение рабочих особей, направленное на воспитание других маток, и т. п.
В данном случае речь идет об определенных химических соединениях, выделяемых одной особью и влияющих на поведение других. Такая химическая регуляция поведения представляет одну из очень интересных и еще малоисследованных особенностей поведения насекомых.
Очень интересна зависимость поведения от степени скученности насекомых данного вида. Поведение же в свою очередь, как оказалось, определяет и многие черты строения насекомого, что было впервые установлено для многих саранчовых Б. П. Уваровым. За последние 10 лет выяснилось, что влияние друг на друга особей одного вида у многих гусениц тоже приводит к различию и их поведения и их строения (как это, например, хорошо заметно на гусеницах ильмового ногохвоста, окраска которых при одиночном и скученном образе жизни различна). В этих случаях имеет значение уже не выделение каких-либо веществ, а воздействие особей друг на друга через нервную систему.
Особенно сложное поведение наблюдается у так называемых общественных насекомых, т. е. у тех, которые всегда живут колониями и иначе существовать не могут. В сущности, колония общественных насекомых - это одна семья, потомство одной самки (у термитов, пчел, муравьев). Для общественных насекомых характерно разделение функций, связанное с различными физиологическими особенностями и различиями в строении отдельных групп особей. В этом коренное отличие общественных насекомых от человеческого общества. В колониях общественных насекомых есть способные к размножению особи - самцы и самки (самок часто называют "матками" или "царицами") - и особи с недоразвитыми половыми железами, неспособные к размножению, но способные заготовлять пищу, выращивать личинок и т. д. Колония общественных насекомых существует именно благодаря такому разделению функций между различными группами особей. Естественно, что слаженная целостная жизнь колонии может обеспечиваться только при определенным образом координированном поведении всех входящих в нее особей. (Подробнее этот вопрос разобран в главе о перепончатокрылых)
Почти все насекомые раздельнополы. Только немногие, например обитающая в гнездах термитов муха Termitoxenia, гермафродиты. В некоторых местах в Калифорнии гермафродитом оказался и австралийский желобчатый червец : у него часть половых клеток превращается в сперматозоиды, часть в яйца, и самооплодотворение происходит внутри особи, имеющей вид самки; иногда часть яиц бывает не оплодотворена - из них развиваются самцы,утратившие биологическое значение. Но в общем для насекомых характерна раздельнополость.
У многих насекомых хорошо выражен половой диморфизм, т. е. самцы и самки сильно отличаются друг от друга. Иногда это признаки, связанные у самок с приспособлением к откладке яиц. Так, очень длинные яйцеклады, отсутствующие, конечно, у самцов, есть у самок наездников (рис. 364), рогохвостов (табл. 50, 5), кузнечиков (табл. 29) и других насекомых. У многих хрущей самки, зарывающиеся в землю для откладки яиц, имеют голени более расширенные, чем самцы. Таких примеров много. Нередко при большой массе откладываемых яиц самки бывают более грузными, чем самцы, и хуже летают, что в итоге приводит к тому, что у самок редуцируются крылья, и тогда самки нередко деградируют. Так, например, у бабочек-мешечниц (Psychidae) и у наших жуков-светляков (Lampyris noctiluca) самки личинкообразные, без всяких зачатков крыльев, у ложнощитовок (Lecaniinae) тело неподвижной самки утрачивает сходство с насекомыми вообще - кажется выступом коры (рис. 215), а у щитовок (рис. 214) - утрачивает все конечности. Самцы же у перечисленных насекомых имеют облик нормальных насекомых. Редко бывают обратные соотношения (например, у инжирного опылителя - рис. 363, 7, 8), когда самец бескрылый, а самки крылаты.

Таблица 29. Кузнечики: 1 - четырехточечный пластинокрыл (Phaneroptera quadripunctata); 2 - обыкновенный пластинохвост (Leptophyes albovittata); 3 - короткокрылый мечник (Gonocephalus dorsalis); 4 - шиповатый пластинокрыл (Tylopsis lilifolia); 5 - он же с распростертыми крыльями; 6 - зеленый кузнечик (Tettigonia viridissima); 7 - певчий кузнечик (T. cantans); 8 - хвостатый кузнечик (T. caudata); 9 - серый кузнечик (Decticus verrucivorus); 10 - виноградный эфиппигер (Ephippiger ephippiger); 11 - степная дыбка (Saga pedo); 12 - степной толстун (Bradyporus multituberculatus)

Таблица 29. Кузнечики: 1 - четырехточечный пластинокрыл (Phaneroptera quadripunctata); 2 - обыкновенный пластинохвост (Leptophyes albovittata); 3 - короткокрылый мечник (Gonocephalus dorsalis); 4 - шиповатый пластинокрыл (Tylopsis lilifolia); 5 - он же с распростертыми крыльями; 6 - зеленый кузнечик (Tettigonia viridissima); 7 - певчий кузнечик (T. cantans); 8 - хвостатый кузнечик (T. caudata); 9 - серый кузнечик (Decticus verrucivorus); 10 - виноградный эфиппигер (Ephippiger ephippiger); 11 - степная дыбка (Saga pedo); 12 - степной толстун (Bradyporus multituberculatus)
Иногда различия в строении самцов и самок не имеют такого явного приспособительного значения. Так, у бабочек-голубянок (Lycaena) голубые крылья имеют только самцы, а у самок крылья коричневые, незаметные.
Иногда самцы и самки различаются по строению ротовых органов. Так, у самки жука-оленя жвалы развиты нормально, а у самца гипертрофированно изменены, превратясь только в "турнирное оружие", как и у настоящих оленей. У самца жука-носорога на голове рогообразный вырост, которого нет у самки, и т. п.
Нередко у насекомых наблюдается полиморфизм, когда, например, у общественных насекомых (пчелы, термиты) разные особи выполняют разные функции, обеспечивающие сохранение вида Так, у пчел есть способные к размножению самки (матки) и рабочие особи - самки с недоразвитыми половыми железами, обеспечивающие снабжение пищей всей колонии. По всему облику эти самки сильно разнятся друг от друга. Еще более разительны различия у муравьев, у которых самки крылаты, а рабочие совершенно бескрылы; у термитов много форм: рабочие, солдаты и др. Резко выражен полиморфизм у тлей, у которых самки бывают бескрылыми или крылатыми, партеногенетическими или оплодотворяемыми в зависимости от условий освещения, питания, температуры.
Обычным способом размножения у насекомых является обычное половое размножение, но нередко наблюдается и партеногенез, чаще встречающийся у насекомых, самки которых бескрылы. Партеногенез обычен у тлей, щитовок, долгоносиков-скосарей, бескрылых эмбий, палочников и др.
Оплодотворение у низших насекомых (чешуйницы) наружно-внутреннее: самец откладывает на землю сперматофор, а самка подбирает его с земли при помощи половых придатков. Близкий способ оплодотворения у низших групп крылатых насекомых как с неполным (таракановые, богомолы, кузнечики, сверчки), так и с полным (верблюдки, вислокрылки, сетчатокрылые) превращением. У них самка подхватывает находящийся ниже ее брюшка сперматофор своими половыми придатками в момент его откладки самцом (рис. 123). Этим достигается сокращение срока пребывания сперматофора во внешней среде. У более высокоорганизованных групп насекомых (бабочки, жуки, двукрылые) самец вводит сперматофор в половое отверстие самки с помощью совокупительного органа, благодаря чему семенная жидкость ни мгновения не остается во внешней среде, т. е. оплодотворение внутреннее. А так как при этом защитная роль оболочки сперматофора становится излишней, при внутреннем оплодотворении часто в половое отверстие самки вводится не сперматофор, а непосредственно семенная жидкость.
Рис. 123. Спаривание кузнечиков
Большинство насекомых откладывает яйца. У разных видов форма яиц очень разнообразна (табл. 20). У насекомых яйца покрыты скорлупой, защищающей их от неблагоприятных воздействий, причем скорлупа образуется до оплодотворения яйца.
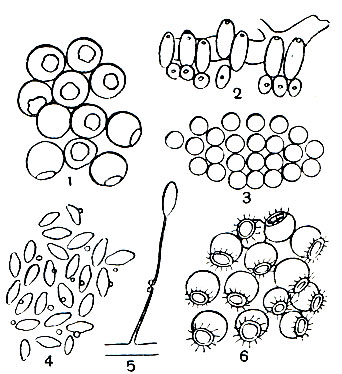
Таблица 20. Многообразие форм яиц насекомых: 1, 6 - яйца клопов (Hemiptera); 2 - яйца колорадского жука (Leptinotarsa decemlineata); 3 - яйца совок (Noctuidae); 4 - яйца стрекоз (Odonata); 5 - яйцо златоглазки (Chrysopa)

Таблица 20. Многообразие форм яиц насекомых: 1, 6 - яйца клопов (Hemiptera); 2 - яйца колорадского жука (Leptinotarsa decemlineata); 3 - яйца совок (Noctuidae); 4 - яйца стрекоз (Odonata); 5 - яйцо златоглазки (Chrysopa)
Сперматозоид проникает в яйцо через особое отверстие в скорлупе, находящееся против ядра яйцеклетки,- через так называемое микропиле. При вылуплении личинка обычно прогрызает оболочку яйца (табл. 21).
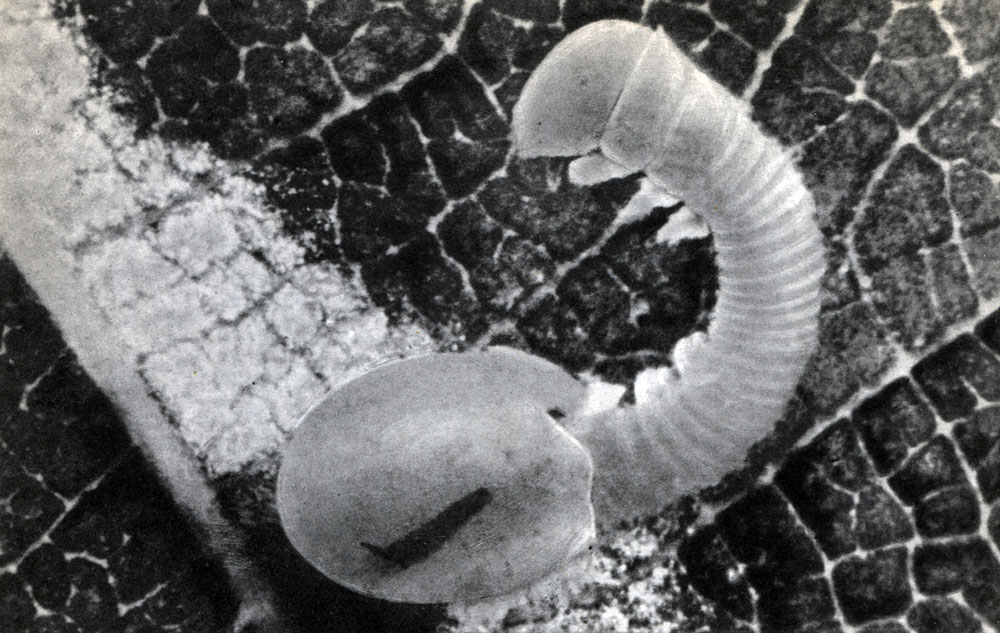
Таблица 21. Вылупление личинки насекомого из яйца. Липового бражника (Mimas tiliae)

Таблица 21. Вылупление личинки насекомого из яйца. Медведки обыкновенной (Gryllotalpa gryllotalpa)
В некоторых случаях можно заставить яйца развиваться и без оплодотворения у тех видов, у которых нормально требуется оплодотворение. Такой "искусственный партеногенез" был впервые получен московским зоологом А. А. Тихомировым на яйцах тутового шелкопряда в конце прошлого века. А недавно тоже в Москве Б. Л. Астауров добился развития шелкопряда только за счет ядра сперматозоида, убив в яйце рентгеновскими лучами ядро, полученное от материнского организма. Такой "андрогенез" представляет, огромный теоретический интерес для определения роли ядра в наследственности. У шелкопряда роль ядра оказалась решающей, полученный андрогенетический шелкопряд имел все признаки отца и никаких признаков матери, хотя материнской плазмы в яйцеклетке много больше, чем отцовской в сперматозоиде. Для представителей всех групп водных беспозвоночных, перешедших к жизни на суше (в почве), а также, для всех групп низших наземных членистоногих, тоже тесно связанных с почвой, характерно то, что у них из яйца выходит молодой организм, очень похожий на взрослый. Это относится и к связанным с почвой низшим насекомым - щетинохвосткам, у которых рост молодого организма не сопровождается существенными изменениями строения. Вероятно, так развивались и предки насекомых, которые жили в таких же условиях. Такой тип развития условно можно назвать прямым развитием. Такое развитие возможно, если в течение всего периода после выхода из яйца животные ведут примерно однотипный образ жизни, обитая в одной и той же среде.
Правда, например, даже у чешуйниц наблюдается, что взрослые формы чаще покидают почву и другие укрытия, чем молодь. Выходы взрослой стадии из почвы на ее поверхность у предков современных насекомых были очень кратковременными, но биологическое значение их было велико - на открытой поверхности облегчается возможность встречи полов и легче происходит расселение. Взрослые формы, имеющие относительно меньшую поверхность тела, чем мелкие молодые, всегда легче выносят временный дефицит влаги (даже у самых влаголюбивых видов). От временных переползаний на поверхности почвы взрослая стадия предков насекомых перешла к более длительным выходам из почвы, к расселению сперва путем такого же переползания, как по ходам в почве, а затем прыжками (махилисы). Более подверженные иссушению спинные участки тела стали более плотными, над боками образовались боковые защитные выросты тергитов - "паранотальные выросты", характерные для многих наземных членистоногих; такие выросты могли дать начало зачаткам крыльев, с помощью которых взрослые насекомые могли сперва перейти к планирующему полету, а затем и к активному. На рисунке 85 приведена гипотетическая схема происхождения насекомых от червеобразных предков.
При переходе взрослой стадии насекомых к жизни в воздушной среде возможны были два пути эволюции.
В одном случае и личинки стали переходить к такому же образу жизни, как взрослые. Но жизнь на открытой поверхности требует более сложного строения. Поэтому у тех насекомых, у которых личинки ведут такой же образ жизни, как взрослые, но уже в открытой воздушной среде, они из яйца выходят на более высокой ступени развития, чем выходили их жившие в укрытиях предки. Это путь, по которому шло развитие насекомых с неполным превращением.
Другой путь эволюции был связан с тем, что личинки продолжали вести скрытый образ жизни в почве и в других укрытиях, приспосабливаясь к нему всё совершеннее, а взрослые насекомые вели открытый образ жизни, активно летая и отыскивая подходящие места для развития потомства. Приспособление ранних стадий к одному образу жизни, а взрослых к другому сделало невозможным переход из личиночного состояния во взрослое путем обычной линьки. Потребовалась стадия куколки - стадия перестройки. Это путь развития насекомых с полным превращением. У насекомых с полным превращением личинки часто имеют очень упрощенное строение; например, у личинок мух нет ни ног, ни головы, весь их облик скорее напоминает червя. Такая простота строения - своеобразное приспособление к развитию в запасах пищи. Более примитивные личинки насекомых с полным превращением похожи по внешнему виду на первичнобескрылых насекомых - у них 3 пары ног, а на конце брюшка бывают церки. Куколка насекомых - непитающаяся стадия, по общему облику более напоминающая взрослое насекомое, чем личинку. На стадии куколки происходит разрушение специфических личиночных органов, осуществляемое в процессе разрушения личиночных тканей (гистолиз), и образование из скоплений особых клеток (имагинальные диски) тканей и органов взрослого насекомого (гистогенез). Чем больше отличается строение личинки от строения взрослого насекомого, тем сильнее процессы гистолиза в теле куколки. Особенно сложные перестройки происходят у таких насекомых, как мухи, у которых в первые дни после окукливания основная масса внутренних органов куколки представляет жидкую кашицу.
Основные типы куколок - свободная (рис. 261, 266, 317) и покрытая (рис. 327, 412). Свободная куколка характеризуется тем, что у нее все придатки (ноги, усики, ротовые части, крылья) не только хорошо различимы, но и легко могут быть отделены иголочкой от поверхности тела, даже если они плотно прижаты (табл. 22). У покрытых куколок придатки, хотя и заметны, но сливаются с общей поверхностью тела и не могут быть отведены иголкой от тела без повреждения насекомого (табл. 22). Особый тип куколки - бочонковидный. Куколки этого типа характерны для высших мух, у которых роль оболочки куколки принимает последняя личиночная шкурка, которая перед окукливанием личинки сильно раздувается и затвердевает (рис. 423, 5).
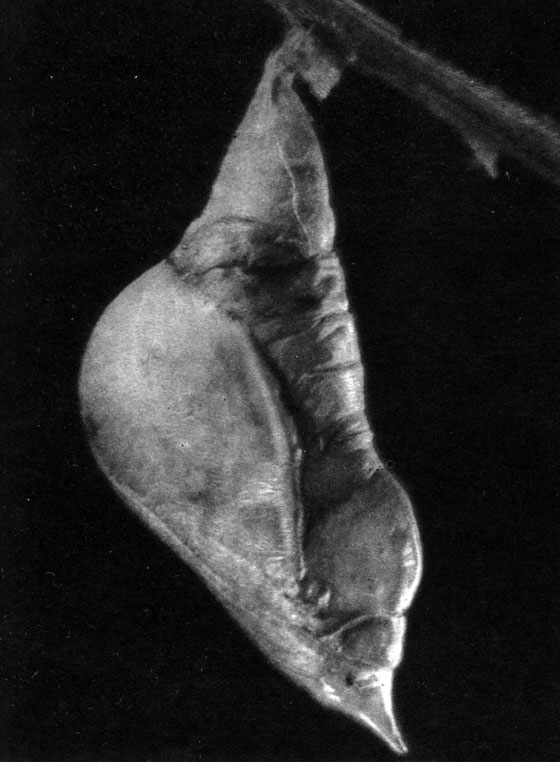
Таблица 22. Типы куколок насекомых и способы окукливания. Покрытая куколка бабочки-лимонницы (Gonepteryx rhamni), окукливающейся открыто на раcтениях
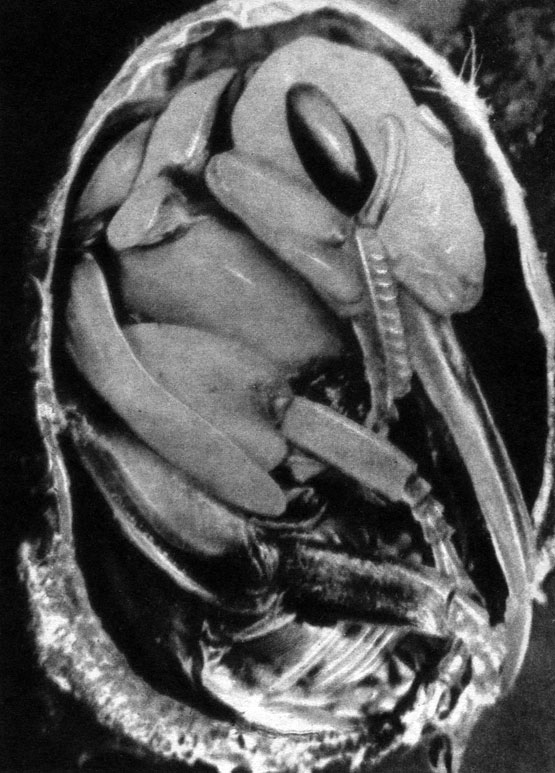
Таблица 22. Типы куколок насекомых и способы окукливания. Свободная куколка шмеля Bombus hypnorum во вскрытой колыбельке
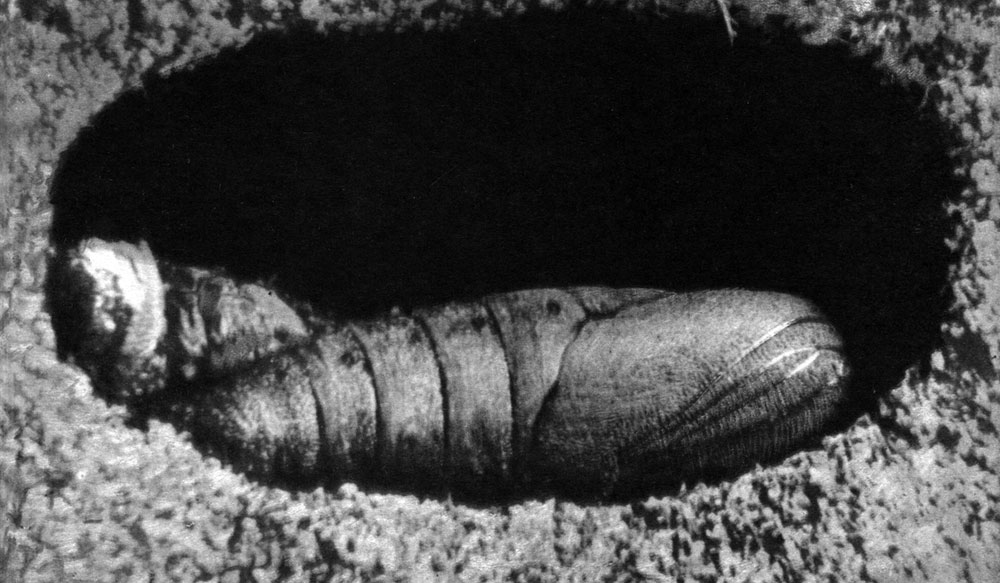
Таблица 22. Типы куколок насекомых и способы окукливания. Покрытая куколка липового бражника (Mimas tiiiae) в колыбельке в почве
Нередко стадия куколки называется "покоящейся". Такое определение неверно, так как, во-первых, в теле куколки идут интенсивные процессы перестройки организма, а во-вторых, у многих насекомых куколки производят энергичные движения. Только бочонковидные куколки совершенно неподвижны, остальные же способны двигать хотя бы брюшком. Развивающиеся в воде покрытые куколки ручейников и комаров превосходно и очень активно плавают. Куколки бабочки древесницы въедливой активно проползают из глубины хода, проложенного гусеницей в древесине, к тому месту, где находится отверстие, через которое выходит бабочка. У верблюдок куколка бегает не менее проворно, чем личинка, а у некоторых мошек, как показал английский ученый Хинтон, куколка даже способна прогрызать шелковый чехлик, который перед окукливанием сплетает личинка.
У некоторых насекомых, ведущих очень сложный образ жизни в стадии личинки, наблюдается так называемый гиперметаморфоз. В тех случаях, когда личинки разного возраста имеют резко различающееся строение, переход одной формы личинки в другую сопровождается прохождением стадии перестройки, обычно неподвижной ("ложнокуколка"). Таким образом, при гиперметаморфозе имеются две стадии перестройки организма - ложнокуколка и куколка.
Рост насекомых, как и всех членистоногих, сопровождается линьками (табл. 23, 24). Из всех открыточелюстных насекомых только у щетинохвосток линьки продолжаются и во взрослом состоянии, да у поденок есть одна линька уже окрылившихся насекомых. Остальные же насекомые во взрослом состоянии не линяют. В личиночном возрасте обычно бывает 4-5 линек, но иногда их не бывает вовсе {некоторые паразитические перепончатокрылые) или их более 20 (личинки некоторых щелкунов, цикад).

Таблица 23. Перелинявшая пустынная capанча (Scliistocerca gregaria), справа - пустая шкурка
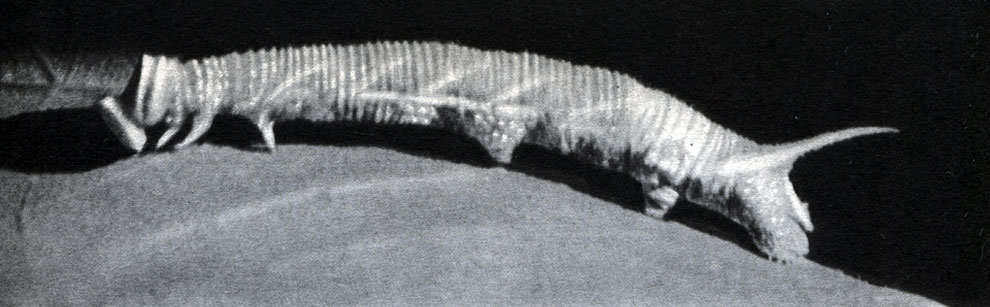
Таблица 24. Последовательные этапы сбрасывания шкурки гусеницей липового бражника (Mimas tiliae) при линьке
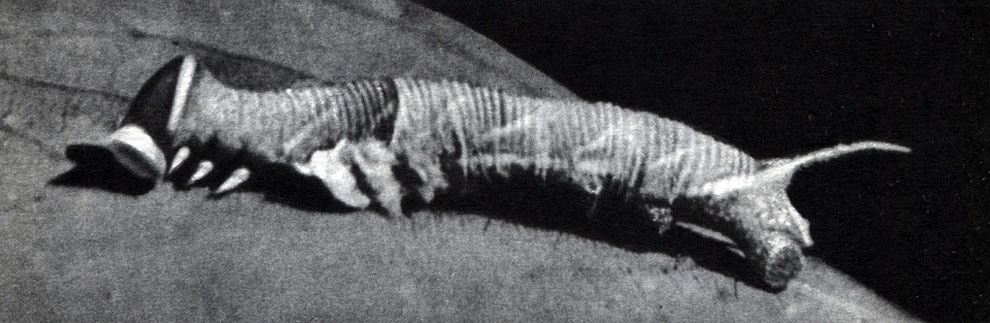
Таблица 24а. Последовательные этапы сбрасывания шкурки гусеницей липового бражника (Mimas tiliae) при линьке
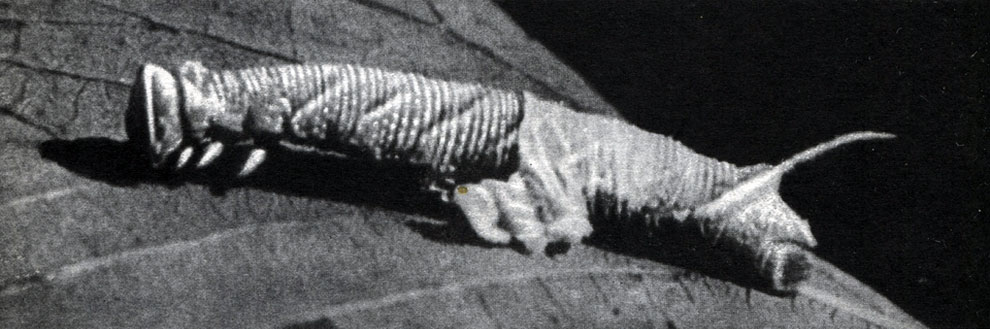
Таблица 24б. Последовательные этапы сбрасывания шкурки гусеницей липового бражника (Mimas tiliae) при линьке

Таблица 24в. Последовательные этапы сбрасывания шкурки гусеницей липового бражника (Mimas tiliae) при линьке
Как и у всех животных с непостоянной температурой тела, скорость развития у насекомых зависит от температуры. В пределах примерно от 6 до 36° С при повышении температуры на каждые 10° скорость развития удваивается. Опытным путем было установлено, что для каждого вида насекомых можно установить то количество "тепла", которое требуется для развития. Наименьшая температура, при которой уже происходит развитие насекомого, называется температурным порогом развития (у большинства насекомых это 6-10°). Среднесуточная температура выше этого "порога" называется эффективной. Для многих видов насекомых умеренной полосы определена та сумма среднесуточных эффективных температур, которая требуется для развития. Как правило, в пределах области распространения того или иного вида в зависимости от местных условий года меняется конкретное число дней, требуемых для развития данного насекомого, но сумма эффективных температур остается величиной более или менее постоянной. Поэтому в зависимости от длины теплого (вегетационного) периода и от среднесуточных температур (обе величины закономерно изменяются при переходе из одних широт в другие) многие виды имеют разное число поколений. Например, луговой мотылек (Loxostege sticticalis) - один из наших хорошо изученных широко распространенных видов вредителей - на севере области распространения (в нечерноземной зоне) дает одно поколение в году, в зоне лесостепи - два, на юге степной зоны - три, а в Закавказье даже четыре. Но многие виды дают меньше поколений в году, чем можно определить по сумме эффективных температур. Бывает, что при благоприятных температуре и влажности развитие насекомого вдруг останавливается. Такая остановка развития имеет приспособительное .значение для насекомого. Например, на Украине гусеницы люцерновой совки {Chloridea dipsacea), развивающиеся из яиц, отложенных в начале лета, окукливаются и через 2 недели из куколок вылетают бабочки. Гусеницы, вышедшие из яиц, отложенных этими бабочками, развиваются примерно столько же времени, сколько развивались и гусеницы первого поколения, тоже окукливаются, но бабочки из этих куколок через 2 недели не выходят. Куколки второго поколения, несмотря на благоприятные условия, останавливаются в развитии и зимуют. Такая остановка развития носит название глубокого покоя или диапаузы.
Диапауза имеет явно приспособительный характер. Если бы у люцерновой совки из куколок второго поколения бабочки вылетали в тот же год, из отложенных ими яиц вышли бы гусеницы, которые не успели бы закончить свое развитие.
Уход в диапаузу связан с существенными изменениями обмена веществ насекомых - с накоплением избытка жира и т. п. "Сигналом", определяющим уход в диапаузу люцерновой совки, оказывается длина светового дня. Если гусениц держать при длинном световом дне, они дают быстро развивающихся куколок, а при сокращенном - диапаузирующих. Температура, влажность - факторы очень изменчивые, зависящие от погоды. А вот длина дня для каждой зоны - величина строго определенная для каждого дня в году. Поэтому естественно, что приспособление к уходу в зимовку в подготовленном состоянии определяется таким постоянно действующим фактором. Ну, а выход из диапаузы в разбираемом случае связан с теми изменениями в организме, которые определяются действием зимних температур. Только после морозного периода диапаузирующая куколка люцерновой совки снова способна к развитию. Так достигается то, что превращение куколки в бабочку осуществляется весной, когда ее потомству обеспечена обильная пища.
В состояние диапаузы разные насекомые могут впадать на разных стадиях развития. У саранчовых известна зародышевая диапауза (в стадии яйца), у желудевого долгоносика диапаузируют личинки, у люцерновой совки - куколки, у колорадского жука - взрослая стадия.
Таким образом, развитие насекомых регулируется теми изменениями их состояния, которые вызываются условиями среды. Но, кроме того, развитие насекомых управляется и внутренними факторами. За последние 10 лет установлен ряд гормонов, регулирующих развитие насекомых. Эти гормоны выделяются нервной системой (мозг, подглоточный ганглий) и специальными железами внутренней секреции - придаточными телами мозга. Для некоторых гормонов, например для гормона линьки (экдизона), уже известно химическое строение, тогда как другие гормоны, например "ювенильный гормон" (неотин), выделяемый придаточными телами, тормозящий созревание, известен только по физиологическому действию.
О влиянии на развитие "феромонов" (называемых также "телергонами") - веществ, выделяемых одними особями и влияющих на развитие других представителей того же вида, упоминалось ранее. Насекомые очень быстро растут и очень плодовиты. Естественно, что и влияние полчищ насекомых на природу огромно. Многие насекомые - важные почвообразователи, разрушители растительного опада. Известно, что в основном насекомыми разлагается, например, опад в зоне хвойных лесов и что там, где нет насекомых, накапливаются волокнистые торфообразные неплодородные слои. В тропиках основными почвообразователями являются термиты, перерабатывающие в лесах всю отмирающую древесину. За свою жизнь насекомые перерабатывают количество пищи, в десятки раз превышающее их вес. Насекомые быстро уничтожают накапливающиеся на поверхности почвы остатки растений, трупы и экскременты животных. Некоторые насекомые, питающиеся разлагающимися растительными и животными остатками, неразборчивы в пище, а некоторые очень специализированы; есть, например, жуки-навозники, развивающиеся только в заячьем помете. Осуществляемую насекомыми очистку почвы от скоплений остатков организмов трудно переоценить. Еще К. Линней подсчитывал, что в тропиках потомство трех мух съедает труп лошади быстрее, чем лев! В известных опытах почвоведа П. А. Костычева листья, подвергавшиеся разложению микроорганизмами, 3 года сохраняли свое строение, а когда на них отложил свои яйца грибной комарик, за 3 дня превратились в перегнойную труху!
Среди насекомых огромное количество видов приспособилось к питанию высшими (цветковыми) растениями. По ископаемым остаткам удалось установить, что наиболее прогрессивные группы насекомых развивались параллельно с цветковыми растениями. Цветковые растения - основной источник и пищи и влаги для большинства насекомых. В то же время большинство цветковых растений (около 80%) в свою очередь непосредственно зависит от насекомых. Оказывается, что привлекающие наш взор пестрота и многообразие цветов обязаны тому, что растения приспособились к опылению насекомыми. Посещающие цветы насекомые обеспечивают перенос пыльцы с пыльников одних цветков на рыльца других, обеспечивая перекрестное опыление. Особенно велика роль как опылителей растений представителей трех высших отрядов насекомых с полным превращением - перепончатокрылых, двукрылых и бабочек - тех групп, в отношении которых и по геологическим наслоениям прослеживается связь с цветковыми растениями.
Многие растения имеют такое строение цветка, какое требует обязательного участия в опылении какого-либо определенного вида или группы видов насекомых.
Насекомые посещают цветки для сбора пыльцы (некоторые жуки, многие перепончатокрылые) или ради сладкого выделения - нектара (мухи, бабочки, многие перепончатокрылые). Иногда насекомые используют цветки как убежище или забираются в цветки и соцветия для пристраивания там своего потомства. И все типы посещения цветков насекомыми, оказывается, используются растениями для обеспечения перекрестного оплодотворения.
Часто цветы бывают ярко окрашенные, но без особого запаха. Они привлекают насекомых, которые ориентируются в основном с помощью зрения. Цветки с мелкими венчиками, как правило, бывают собраны в соцветия, чтобы привлекать насекомых, которые их лучше в таком случае замечают. Иногда рядом с невзрачным цветком для привлечения насекомых развиваются яркие прицветные или кроющие листья, как у белокрыльника, иван-да-марьи. Для привлечения насекомых, летающих ночью или в сумерки, особое значение имеет запах; обычно открывающиеся в темноте цветки имеют белую окраску, даже для нас более заметную при недостатке освещения.
Не всегда запах цветов бывает приятен для человека - цветы, привлекающие падальных мух, обладают трупным запахом и иногда имеют окраску венчиков мясного цвета (например, тропическая раффлезия). У многих цветков нектар находится вблизи поверхности и достать его нетрудно (бузина, липа, селезеночник). Такие цветки опыляются случайными посетителями, нередко мухами и даже мелкими жуками. Цветки с полускрытым нектаром, такие, как у различных крестоцветных, у которых нектарники находятся на дне короткой трубочки венчика или защищены волосками, как у розоцветных (яблоня, земляника), опыляются мухами и некоторыми перепончатокрылыми, имеющими короткий хоботок. Обычно такого типа цветки правильные (с радиально расположенными лепестками венчиков) и имеют белую, желтую или розовую окраску. Есть же такие цветки, и их много, у которых нектар спрятан глубоко в недрах сросшихся лепестков венчика. Такие цветки чаще бывают неправильными (двустороннесимметричными), и окраска таких цветков чаще бывает синих, пурпурных и фиолетовых тонов. Нектар таких цветков доступен не для любых насекомых, а только для определенных, имеющих длинный хоботок и нередко достаточно сильных, чтобы раздвинуть лепестки при доставании нектара. Такая форма цветка заставляет определенный вид насекомого, высасывающего нектар, занять на цветке определенное положение, которое обеспечит покрытие пыльцой тех частей тела крылатого гостя, которые войдут в контакт с рыльцем следующего посещаемого цветка.
Приспособления к переносу пыльцы бывают различными. Иногда пыльца просто легко прилипает к телу насекомого, посещающего цветок, как например у лилии, мака. В этих случаях почти любое насекомое, привлекаемое цветком и переходящее с него на другой, обеспечивает опыление. Но не так легко достать нектар из цветка, например, шалфея, доступ к нектару которого закрыт особыми пластинчатыми выростами оснований тычинок. Только сильное насекомое, например пчела, садясь на цветок, может раздвигать лепестки шалфея. Устремляясь к нектару, пчела головой нажимает на вырост тычинки, пыльник которой ударяет пчелу по спинке. При посадке на следующий цветок шалфея пчела именно тем местом, на которое попала пыльца, соприкасается с рыльцем и переносит таким образом на него пыльцу (рис. 124). Сходно приспособление к опылению у дельфиниума.
Рис. 124. Опыление шалфея: 1 - пчела, опыляющая цветок (пыльник прикасается: к спинке насекомого); 2 - тычинка цветка шалфея
Обычно привлечение насекомых цветком вызывается пищевыми раздражителями, но не всегда. Например, некоторые орхидеи опыляются самцами шмелей. Было выяснено, что цветок таких орхидей напоминает очертания самки шмеля и привлекательность цветка связана с половым инстинктом насекомого.
Очень своеобразные отношения сложились между американским растением юккой и юкковой молью (Pronuba juccasella). Самка юкковой моли по вечерам посещает свисающие цветки юкки и собирает комок пыльцы с помощью специально видоизмененных челюстей, в которых переносит этот комок на другой цветок юкки. Своим яйцекладом она прокалывает завязь, откладывает в нее яйца, а после этого спускается по столбику и засовывает комок пыльцы в рыльце (рис. 125). Таким образом моль обеспечивает опыление юкки, а тем самым и развитие своего потомства - выходящие гусеницы питаются завязывающимися семенами, часть которых съедают. Не менее удивительна взаимная приспособленность фигового дерева и его опылителя - хальциды-бластофаги.
Рис. 125. Опыление юкки юкковой молью (Pronuba juccasella)
Нередко недостаток опылителей определяет неурожай многих растений. Это хорошо известно тем, кто имел дело с плодовыми деревьями - яблони, груши без достаточного количества пчел не дают плодов. Некоторые культурные растения зависят не от домашней пчелы, а от диких опылителей. Например, клевер опыляется в основном шмелями. Не всякая домашняя пчела обеспечивает опыление клевера. Хоботок нашей среднерусской пчелы недостаточно длинен, чтобы доставать нектар из глубоких нектарников цветков клевера; только длиннохоботковая кавказская раса пчелы может опылять клевер. Но, кроме того, пчел на клевер нужно дрессировать. Известно, что пчела, раз начав посещать цветки определенного вида растений, продолжает посещать преимущественно их весь период цветения. Подкормка пчел сиропом, изготовленным на цветках клевера, повышает посещаемость клевера пчелами. Иногда приходится дрессировать пчел и на посещение таких цветов, у которых нектар легкодоступен. Например, чтобы получить хороший урожай семян рапса, недостаточно расположить вблизи посева этой культуры пасеку; надо предварительно провести дрессировку пчел на рапс.
Есть много культурных растений, например люцерна, опыляющихся дикими пчелами, целиком заменить которых домашней пчелой нельзя.
Не могут опыляться пчелами и такие цветы, которые открываются в темное время суток. Такие цветы, особенно имеющие глубоко расположенные нектарники, как, например, душистый табак, опыляются ночными бабочками, преимущественно бражниками и совками.
Если питание нектаром и избыточной пыльцой растений насекомых-опылителей приносит пользу растениям, то поедание насекомых других частей растений причиняет им в большинстве случаев явный ущерб.
Характер повреждений различными насекомыми тканей растений очень многообразен. Он зависит от того, какие органы растения повреждаются насекомым и где находится насекомое в момент повреждения, и от строения ротового аппарата насекомого. Для вредящих растениям насекомых характерны 2 типа ротового аппарата - грызущий и колюще-сосущий. Насекомые с грызущим ротовым аппаратом грубо откусывают участки тканей растения. Если грызущие насекомые питаются листьями растений, они либо "неправильно" объедают лист, съедая кусок за куском, как это делают крупные жуки, например свекловичный долгоносик на свекле, либо оставляют только толстые жилки ("скелетируют" лист), как это делает тополевый листоед на тополе, либо прогрызают в листьях окошечки, как жуки-щитоноски и их личинки на лебеде, либо обгрызают края листовой пластинки, как жуки клубеньковые долгоносики на листьях гороха. Всех способов нанесения повреждений (рис. 126) не перечислить, но по характеру повреждений, которые бывают обычно очень типичны для каждого вредителя, питающегося на данном виде растения, определить вид насекомого бывает даже легче, чем по признакам строения самого вредителя. Те насекомые, которые питаются твердыми или мясистыми частями растений (стволы, стебли, плоды), чаще вгрызаются в ткани, прокладывая там ходы. Такие ходы тоже бывают настолько индивидуальны, что специалисту спутать, какому виду принадлежит ход, например, короеда, невозможно.

Рис. 126. Типы повреждений различных частей растений насекомыми: 1 - грубое скелетирование листа капусты гусеницей капустной белянки; 2 - дырчатое повреждение молодого листа свеклы личинкой щитоноски; 3 - повреждение стеблей пшеницы личинкой узловой толстоножки; 4 - неправильное объедание листа свеклы гусеницей лугового мотылька; 5 - окончатое повреждение листа редиса земляной блошкой; 6 - вздутая пузыревидная мина на листе крестоцветного, поврежденного минирующей мухой; 7 - скручивание листа яблони, поврежденного яблоневой тлей; 8 - скручивание листьев свеклы, поврежденных свекловичным клопиком; 9 - тонкое скелетирование листа яблони молодыми гусеницами яблоневой моли; 10 - повреждение корзинки подсолнечника гусеницей подсолнечниковой моли; 11 - повреждение початка кукурузы гусеницей хлопковой совки; 12 - повреждение коробочки льна гусеницей льняной листовертки; 13 - личинка люцернового долгоносика и поврежденный ею корень люцерны; 14 - мины на листе брюквы, поврежденном минирующей молью; 15 - краевое объедание листьев гороха клубеньковыми долгоносиками
Большие систематические группы насекомых приспособились к развитию в лубе древесных пород (лубоеды), в древесине (многие усачи, златки, рогохвосты), в плодах (долгоносики-ринхиты), в желудях и орехах (долгоносики-плодожилы), в головках клеверов (долгоносики-семееды), в соломинах злаков (хлебные пилильщики), в толстых стеблях травянистых растений (горбатки, долгоносики-фрачники), в клубеньках на корнях бобовых (долгоносики-ситоны) и т. д.
Многие мелкие насекомые поселяются в листовых пластинках, проделывая в них ходы - мины, которые снаружи хорошо заметны и на зеленом фоне листа кажутся белыми. Есть целые семейства минирующих мух, минирующих молей и др., именно таким образом вредящих растениям.
Развитие внутри растительных тканей создает очень благоприятные условия для многих насекомых - внутри растения воздух насыщен водяным паром, как и в почве. Поэтому обитающие внутри растения личинки насекомых избавлены от самого неблагоприятного воздействия которому могут подвергаться наземные насекомые,- им не угрожает высыхание. Поэтому, например, в пустынях, где воздух очень сух, большинство растительноядных насекомых либо обитает в почве, повреждая корни растений, либо развивается внутри растительных тканей. Не надо думать, однако, что в сухих местностях нет открыто живущих вредных для растений насекомых,- их очень много и они в этих условиях особенно опасны. Теряя влагу, они стремятся восполнить ее запасы за счет сочных тканей растений, и потому именно в сухих местностях вредные насекомые наносят особо сильный ущерб растениям. Достаточно напомнить, что именно в сухих местностях распространены и сильно вредят различные саранчовые, съедающие в годы массовых размножений всю зелень, встречающуюся на их пути.
Те насекомые, которые высасывают соки из растений, не причиняют таких бросающихся в глаза нарушений повреждаемым органам, однако в результате их деятельности растения тоже очень страдают. Если вредитель, например тля, поселяется на листьях, листья начинают сперва светлеть, затем желтеть, а потом и завядать. Нередко происходит скручивание листьев - это наиболее простая деформация листа, вызываемая сосущими насекомыми. При массовом поселении сосущих насекомых на корнях (например, кровяной тли на яблоне) или на стволах (например, запятовидной щитовки на саженцах яблони) растения могут засохнуть. Нередко поселение на растении сосущих, а также развитие в тканях растений и некоторых грызущих насекомых вызывает разрастание тканей растений, которое либо имеет вид неправильных наростов, либо напоминает какие-либо плоды.
Такие разрастания тканей растений под влиянием механических раздражений и химических воздействий, причиняемых насекомыми (или другими организмами), носят название галлов (рис. 127, табл. 58). Так, галлы, вызываемые орехотворками на листьях дуба, напоминают плод типа костянки: внутри одетого румяной кожуркой мягкого, похожего на яблоко шаровидного галла вокруг тела личинки даже образуется затвердение тканей листа, напоминающее косточку. Галлы, вызываемые хермесами на молодых побегах ели, несколько напоминают шишку, а на листьях береста, на которых поселяется берестовая тля, вокруг тли образуются орешкоподобные или стручкообразные галлы, которые растрескиваются, когда тля закончит свое развитие и должна вылетать. Образование галлов - результат длительной эволюции, приспособления друг к другу растения и насекомого. Растение "создает" благоприятные условия для существования насекомого, но как бы изолирует насекомое, защищая этим ткани, удаленные от места сосания, от его воздействия.
Рис. 127. Галлы и мины, вызываемые насекомыми: А - мешетчатые галлы тли Tetraneura ulmi на листе вяза: 1 - пораженный лист, 2 - наружный вид галла, 3 - галл в разрезе; Б - мешетчатые галлы тли Pemphigus bursarius на листе тополя; В - галлы дубовой орехотворки (Diplolepis quercusfolii) на листе дуба; Г - корневые галлы личинки скрытнохоботника Ceuthorrhynchus pleurostigma на корнях сурепки: 1 - общий вид галла, 2 - галл в разрезе; Д - галлы (бедегуар) орехотворки Rhodites rosae на побегах розы: 1 - общий вид пораженного побега, 2 - галл в разрезе; Е - мины насекомых на листьях: 1 - узкая мина личинки мухи Ophiomyia maura на листе золотой розги, 2 - широкая мина личинки мухи Pegomyia nigritarsis на листе щавеля, 3 - широкая мина гусеницы Phyllocnistis sorhageniella на листе осины
Важным моментом, определяющим галлообразование, является выделение насекомыми ростовых веществ типа ауксинов. Известно, что, применяя обработку растений гетероауксином, можно вызвать усиленный рост некоторых органов, например органов плодоношения у томатов, что применяется в нашей практике. Воздействие насекомых на формообразовательные процессы у растений гораздо тоньше и гораздо эффективнее - изучение галлов может дать много нового для понимания процессов развития растений.
Те насекомые, которые питаются культурными растениями, причиняют огромный вред народному хозяйству. Достаточно указать, что по подсчетам, сделанным 10 лет назад, только сельское хозяйство нашей страны несет ежегодно потери порядка 4,5 миллиарда рублей (в современном исчислении) от вредителей и болезней. Это без учета потерь от вредителей леса! Только от вредных насекомых (не считая потерь от болезней растений) потери сельского хозяйства в США составляют 4 миллиарда долларов в год. Эти поистине чудовищные цифры в большой мере объясняют то, что на изучение насекомых направлены усилия огромного числа специалистов, работающих во всех странах мира. Достаточно сказать, что каждые несколько минут из печати выходит научная работа по энтомологии - разделу теоретической и прикладной зоологии, который посвящен насекомым.
Но никогда не следует забывать, что, если бы провести точные подсчеты той пользы и того вреда, который приносится насекомыми, польза бы значительно перевесила. Достаточно вспомнить, что без насекомых не давали бы урожая многие важные культурные растения - плодовые, гречиха, крестоцветные, подсолнечник и др. и многие просто бы погибли. Если бы не было жуков-навозников, погибли бы пастбища.
Что это не досужее предположение, показывает такой интересный случай из истории сельского хозяйства Австралии. В Австралии некоторые пастбища стали погибать из-за того, что на поверхности почвы стали скапливаться неразлагающиеся и затрудняющие рост травы кучи коровьего навоза. Оказалось, что там не было навозников. Завоз навозников и их акклиматизация быстро привели к разложению скоплений навоза и к большому повышению урожайности начавших приходить в запустение пастбищ.
Когда мы говорим о насекомых-вредителях, мы имеем в виду не тот вред, который насекомые причиняют растениям, а тот урон, который они наносят нашей экономике. Например, когда в Австралии в массе размножились на пастбищах колючие кактусы, мешавшие пастьбе скота, туда были завезены насекомые, истребляющие кактусы, которые рассматривались как полезные насекомые. А когда были выведены бесколючковые формы кактусов, которые охотно поедаются скотом, виды насекомых, истребляющих кактусы, из полезных стали вредителями.
Из насекомых, дающих человеку непосредственно используемые им продукты, только медоносная пчела приносит пользу и растениям. Кстати сказать, польза от пчелы как от опылителя культурных растений во много раз превышает ту пользу, которую мы получаем, используя мед и еще более ценный продукт, широко применяемый в технике,- воск. Другие непосредственно полезные для нас насекомые повреждают те растения, на которых питаются. Их не так много. Это тутовый шелкопряд, гусеницы которого выделяют при постройке кокона шелк. Несмотря на замечательные достижения современной химии синтетических волокон, шелк оказывается непревзойденным еще материалом. Шелк по степени полимеризации и по зависящим от нее механическим качествам нити превосходит лучшие сорта капрона, нейлона и других синтетических волокон. Поэтому для наиболее ответственных, требующих высокой прочности изделий,- для парашютов, для герметических прокладок -натуральный шелк представляет пока лучший материал. Другой шелкопряд - китайский дубовый шелкопряд - дает очень прочную, но более грубую нить, из которой изготавливается чесуча.
Очень ценный продукт - шеллак дают обитающие на ряде древесных пород на юго-востоке Азии лаковые червецы. Воскоподобные вещества, выделяемые этими червецами, обладают исключительными изоляционными свойствами и находят широкое применение в электротехнике, особенно в радиотехнике.
Другие червецы дают краску: как источник ценной краски кармина известны обитающие на средиземноморском дубе червецы кермесы (Kermes), значение которых было особенно велико в древности, но снизилось, когда стали добывать кармин из живущих в Мексике на кактусе опунции кошенилей (Coccus cacti). Теперь с развитием химии красителей значение этой краски несколько уменьшилось.
Некоторое значение в медицине имеет кантаридин, содержащийся в жуках "шпанских мушках", обгрызающих листья ясеня. Кантаридин - активное вещество нарывного пластыря. Вот в сущности и все те непосредственно полезные для нас насекомые, которые дают используемые человеком вещества.
Медоносная пчела, тутовый и дубовый шелкопряды, лаковые червецы и кошениль - немногие примеры полуодомашненных насекомых, разводимых человеком ради доставляемых ими продуктов.
Среди остальных питающихся растениями видов насекомых мы сталкиваемся с более или менее безразличными для нас, которые питаются не используемыми человеком видами растений, и с вредителями, которые повреждают культурные растения, становясь нашими конкурентами.
Среди вредных для культурных растений насекомых можно выделить две биологические группы. Одна включает многоядных вредителей, которые малоразборчивы в выборе пищи и повреждают целый ряд культурных (полевых, огородных и др.) растений. Таковы, например, личинки щелкунов, большинство саранчовых, гусеницы лугового мотылька, озимой совки и многие другие. Другая группа - специализированные вредители, питающиеся одним или немногими видами культурных растений. Это, например, такие насекомые, как вредящий сахарной свекле свекловичный долгоносик, вредящая зерновым культурам шведская мушка, повреждающая виноградную лозу филлоксера и др.
Все вредители культурных растений перешли на них с дикорастущих. Как правило, многоядные вредители переходят на посевы культурных растений быстрее. Когда, например, распахиваются целинные земли, развивающиеся в почве и питающиеся корнями разных растений личинки щелкунов принимаются за питание корнями посеянного по нови растения. Переход специализированных вредителей с дикорастущих растений на родственные культуры иногда совершается очень медленно. Так, например, в Красноярском крае долгоносик льняной скрытнохоботник стал вредить льну только лет через сто после введения его там в культуру, а до этого существовал за счет дикорастущего желтого льна.
Мальвовая моль в Армении недавно перешла с дикорастущей мальвы на хлопчатник и стала опасным вредителем его коробочек, а в Средней Азии она хотя и встречается на мальве, но на хлопчатник не переходит. А, например, при широком введении в культуру кок-сагыза в нашей стране в 30-х годах с первых же лет на его посевах в массе встречалось несколько видов вредителей, перешедших с родственных кок-сагызу дикорастущих одуванчиков. Большинство вредителей культурных растений - представители местной фауны, ранее питавшиеся за счет местных дикорастущих растений. Часто новые культурные растения, которые не имеют близкородственных дикорастущих видов среди местной флоры, оказываются почти неповреждаемыми вредителями. Например, эвкалипты на родине, в Австралии, страдают от многих вредителей, а в Европе и Азии практически не повреждаются вредителями. В таких случаях очень важно для защиты посевов не допустить вредителя из удаленных местностей на территорию, где вредителя нет. Это достигается путем применения карантинных мер, контроля за тем, чтобы с посадочным материалом и с разными грузами вредитель не последовал за растением. История знает случаи завоза очень опасных вредителей, которые в массе размножились и стали причинять большие опустошения там, где их ранее не было. Так, в Северную Америку был завезен из Европы стеблевой, или кукурузный, мотылек, ставший там бичом кукурузы. Еще ранее из Европы туда попала сильно вредящая пшенице гессенская мушка. Около 100 лет назад в Америку был завезен непарный шелкопряд, объедающий там леса и сады сильнее, чем на родине, в Европе. В Европу из Америки тоже около 100 лет назад завезли виноградную филлоксеру, разорившую виноградное хозяйство Франции, и широко распространившуюся кровяную тлю, губящую яблони у нас на юге.
В нашей стране нет многих опасных вредителей, таких, например, как "розовый червь" (гусеница) хлопчатника, уничтожающий хлопковые семена и волокна в коробочках (плодах).
Строгий карантинный контроль, облегчаемый у нас монополией внешней торговли, позволяет нашей стране не допускать многих опасных вредителей, в других странах причиняющих большой ущерб.
Следует иметь в виду, однако, что и активные перелеты насекомых-вредителей, и расширение транспортных связей делают задачи карантина все более сложными. И некоторые вредители, еще недавно у нас отсутствовавшие, проникают на нашу территорию. Примером может служить колорадский жук. У нас картофель не знал серьезных вредителей. Около 100 лет тому назад в Америке в Скалистых горах с дикого картофеля на культурный перешел колорадский жук, быстро приспособившийся к новому для себя виду растений и распространившийся по всей территории США до Атлантического океана уже к 1874 году. В начале XX века неоднократно его завозили случайно в Европу, но быстро уничтожали в местах появления. Во время первой мировой войны он был завезен во Францию, но обратили на него внимание только тогда, когда он размножился на значительной территории. За период с 1922 года (когда его заметили) до второй мировой войны колорадский жук достиг границ Германии, а во время второй мировой войны он проник далеко на восток. Фашистская оккупация способствовала его завозу в Чехословакию, Югославию, Польшу, даже на запад УССР. В СССР его очаги были ликвидированы, но постепенное распространение жука (несмотря на строгие мероприятия по борьбе с ним) в ГДР и Польше привело к тому, что он продвинулся на восток и проник на нашу территорию на Украине, в Белоруссии, Прибалтике (рис. 128). Так колорадский жук стал объектом не только внешнего, но внутреннего карантина в нашей стране.
Рис. 128. Пути распространения колорадского жука (по Венгореку)
Чтобы стать вредителем культурного растения, насекомое должно не только питаться им, но и приспособиться к существованию в тех условиях, которые создаются при его возделывании. А эти условия в значительной мере в руках человека. Правильно организуя севообороты, выбирая приемы агротехники, которые благоприятны для растения, но неблагоприятны для вредителя, можно подавить развитие вредителя и даже свести на нет наносимый им вред.
Замена бессменной культуры пшеницы правильными севооборотами на юге УССР и Северном Кавказе привела к снижению численности хлебного жука-кузьки, к тому, что перестали вредить земляные усачи, пырейные огневки и другие виды. Распашка меж, связанная с переходом от единоличных форм землепользования к коллективным, привела к ликвидации вреда от жука-кравчика. Зяблевая вспашка и ликвидация послеуборочных остатков уничтожают зимующие яйца капустной тли, хлебных пилильщиков и других вредителей. Занятые пары сдерживают размножение озимой совки и т. д. Большое значение имеет выведение устойчивых к вредителям сортов растений-подсолнечника к подсолнечниковой огневке (рис. 129), яблони к кровяной тле и т. д. Комплекс агротехнических мероприятий, меняя среду обитания насекомого и растения, является решающим звеном в деле защиты растений.
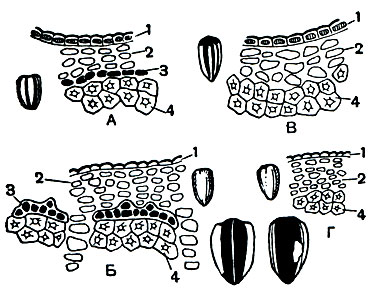
Рис. 129. Сорта подсолнечника, устойчивые (панцирные - А, Б) и неустойчивые (беспанцирные - В, Г) к повреждениям гусеницей подсолнечниковой огневки: 1 - эпидермис; 2 - пробковая ткань; 3 - панцирный слой; 4 - склеренхима
Большую роль в снижении численности вредителей играют их естественные враги - хищники, паразиты, возбудители болезней.
Среди насекомых большое число видов - хищники, а многие развиваются паразитируя в других насекомых. Использование таких естественных врагов насекомых-вредителей позволяет не допускать потери от них урожая. Эти мероприятия составляют основу биологического метода борьбы с вредителями.
Особенно хорошие результаты дает биологический метод против тех вредителей, которые были завезены и не встретили эффективных местных врагов. Показательны такие примеры, как кровяная тля. Этот вредитель корней яблони-американского происхождения, и, например, в СССР не было такого естественного врага тли, который бы сдерживал ее размножение. Ввоз ее паразита наездника афелинуса (Aphelinus mali) дал во многих районах прекрасные результаты. Блестящий пример успешной биологической борьбы дает история австралийского желобчатого червеца. Но бывает и так, что ввоз естественных врагов не дает эффекта. Например, в США для борьбы с непарным шелкопрядом, завезенным из Европы, были завезены все главные враги этого вредителя, сдерживающие его размножение. Но на новой родине хищники и паразиты либо не прижились, либо перешли на питание другими насекомыми - биологическая борьба с непарным шелкопрядом не удалась. Были такие примеры и у нас. Например, против калифорнийской щитовки в Краснодарский край были завезены с Дальнего Востока ее враги - хищные божьи коровки хилокорусы. Хилокорусы прижились и размножились, но... перешли к питанию другими щитовками, а не калифорнийской.
Для борьбы с вредителями можно использовать и местных хищных и паразитических насекомых. Например, для борьбы с тлями в предгорных равнинах используют местных божьих коровок - хищников, в массе уничтожающих тлей. Божьи коровки в горах зимуют большими скоплениями. Их собирают и держат в холодильниках, пока не появятся на культурных растениях тли, а затем выпускают.
В некоторых случаях в борьбе с многими вредными гусеницами хороший результат дает использование наездника-яйцееда трихограммы, развивающейся в яйцах многих чешуекрылых. В лабораторных условиях разводят трихограмму на яйцах зерновой моли, а в период массовой яйцекладки вредителей (озимой совки, яблонной плодожорки) яйца моли с находящейся в них трихограммой выносят на защищаемые участки: трихограмма вылетает и заражает яйца вредителя. Очень важно для подавления численности вредителей не только разводить его паразитов, но и создавать для них благоприятные условия. Например, большинство наездников, личинки которых развиваются во вредных насекомых, убивая их, во взрослом состоянии нуждаются в питании нектаром. Поддерживание вблизи полей и садов цветущих круглое лето растений повышает число наездников.
Конечно, важное звено в системе мер борьбы с вредными насекомыми представляет привлечение насекомоядных птиц. Обнадеживающие результаты дает применение культур микроорганизмов, вызывающих заболевания и гибель вредных насекомых. К ним относится препарат "энтобактерин", с успехом испытанный против ряда вредных гусениц, культура грибка боверии (Beauveria bassiana), опыливание которой во влажные годы вызывает гибель колорадского жука, и др.
Однако часто ни агротехнических мероприятий, ни использования естественных врагов насекомых не бывает достаточно для защиты культурных растений. В практике все шире применяется химический метод борьбы с вредными насекомыми, чаще всего основанный на том, что защищаемые растения и находящиеся на них вредители покрываются тончайшим слоем ядовитого для насекомых вещества. Яды наносятся на растения либо путем опрыскивания, обычно в водном растворе или в виде растворов в маслах и эмульсиях, либо, теперь чаще, в виде мельчайшего порошка (опыливание). А за последние 10-15 лет внедряется и использование ядов в виде мельчайшей взвеси в воздухе (аэрозоли).
Для химической борьбы с вредными насекомыми либо применяют специальные наземные машины (опрыскиватели), либо яды распыляют с помощью самолетов - метод, особенно распространенный у нас для борьбы с вредителями лесов, полевых культур и садов. Авиаметод в борьбе с вредителями был впервые введен в практику в СССР. Для борьбы с вредителями, имеющими грызущий ротовой аппарат, например против саранчовых, применяются яды кишечного действия, попадающие в организм насекомого вместе с пищей. Это в основном различные неорганические соединения - соли мышьяковой и мышьяковистой кислот, фтористые соединения и др.
За последние десятилетия было синтезировано много хлорорганических соединений, очень эффективных в борьбе с вредителями. Это прежде всего ДДТ (дихлордиэтилтрихлорэтан), ГХЦГ (гексахлорциклогексан), алдрин, диэлдрин, гептахлор и др. Они действуют не только попадая в кишечник, но и контактно, попадая в организм насекомого через покровы. На многих насекомых эти яды действуют в совершенно ничтожных количествах и могут применяться и против грызущих, и против сосущих насекомых. Другая группа синтетических ядов - фосфорорганические (меркаптофос, тиофос, вофатокс и др.). Многие из них впитываются растением (не вызывая его гибели) и отравляют насекомое, высасывающее из него соки, действуя как "системный яд", а не только при контакте с вредителем. Это позволяет бороться даже со скрытоживущими вредителями.
Фосфорорганические яды очень ядовиты и для человека, что ограничивает их применение. Некоторые хлорорганические яды (ДДТ) сначала считались безвредными для теплокровных организмов. Однако теперь выяснилось, что ДДТ, например, накапливается в организме человека, вызывая тяжелые изменения в печени и в нервной системе, что заставляет ограничивать применение таких препаратов.
Хлорорганические и фосфорорганические препараты отличаются тем, что действуют не только на вредителей, но и на других насекомых, в том числе и полезных - опылителей и энтомофагов (хищников и паразитов вредителей), а также на многих почвообразователей.
Поэтому, несмотря на высокую эффективность и широкое применение химических мер борьбы с вредителями, в последние годы все больше специалистов высказывается за ограничение применения современных синтетических ядов. Нередко в местностях, где регулярно проводится борьба с вредителями химическими методами, оказываются уничтоженными естественные враги и того вредителя, с которым боролись, и враги тех вредителей, борьба с которыми не предполагалась. Например, в плодовых садах, регулярно обрабатываемых ДДТ против плодожорки и других насекомых, в массе размножаются паутинные клещики, не погибающие от ДДТ, а даже становящиеся от него более плодовитыми. Это связано с тем, что ДДТ убивает тех хищных насекомых, которые сдерживают размножение клещиков.
Неизбирательное уничтожение большинства насекомых при химической борьбе приводит к гибели насекомоядных птиц - в итоге уничтожаются факторы естественной регулировки численности вредителей. На последнем Международном энтомологическом конгрессе в 1964 году в Лондоне многие крупнейшие специалисты высказывались за разработку той системы мероприятий, которая исключала бы массовое применение химических мер борьбы с вредителями. Такую интегральную борьбу с вредителями приветствовали и участники II Международного конгресса по защите растений в 1965 году в Неаполе. Основой в деле борьбы с вредителями должно быть то правило, что легче не допускать размножения вредителя, чем потом его уничтожать на больших площадях. Надо создать такой комплекс агротехнических (а в лесу - лесохозяйственных) условий и такой биологический режим, при котором бы не было предпосылок для массовых размножений вредителей. Химическую борьбу надо проводить не на больших площадях, когда вредитель размножился, как это часто во всем мире делают сейчас, а в самом начале вспышек размножения, на тех небольших участках, где вредитель только начинает появляться в больших количествах. Это важно и потому, что для успешного уничтожения вредителя химическим методом нужно, чтобы мероприятие было проведено в определенный срок, когда вредитель наиболее доступен для уничтожения и наиболее чувствителен к ядам. Своевременное проведение борьбы на ограниченном участке - выполнимая задача. На больших же площадях, даже при использовании самолетов, борьба растягивается и может совпасть с периодами, когда яд на вредителя не действует. При таком синтезе биологических и химических мер борьбы следует ожидать хороших результатов. Но организовать такую борьбу можно только при глубоком знании особенностей и вредителей, и их паразитов, и хищников, хорошо зная жизнь животных!
Нередко применение химических мер борьбы с вредителями встречается с неожиданным затруднением. Вредители, прежде чувствительные к яду, становятся устойчивыми к нему. Например, во многих местностях чувствительность комнатной мухи к ДДТ уменьшилась в 10 раз. В Западной Европе во многих местностях резко снизилась чувствительность колорадского жука к ДДТ и линдану. Такие изменения наследственных особенностей насекомых под влиянием регулярно действующего нового фактора естественного отбора представляют не только серьезную практическую проблему (время от времени приходится менять применяемые яды), но и большой биологический интерес, так как показывают, что у насекомых за короткое время могут измениться наследственные свойства. Большая плодовитость и высокая скорость их размножения недаром сделали многих насекомых теми объектами, на которых решаются закономерности наследственности (дрозофила, мучной хрущак, шелкопряды и др.). Конечно, вредители растений и борьба с ними - серьезная экономическая проблема, стоящая перед человечеством. Но не менее, а часто более важны те насекомые, биология которых связана с теплокровными животными и непосредственно с самим человеком.
Например, только от кожных оводов ежегодная потеря продукции крупного рогатого скота в СССР составляет около 0,5 миллиарда рублей. Огромные убытки животноводству и снижение работоспособности людей причиняет в осваиваемых таежных и других лесных районах так называемый "гнус" - совокупность различных кровососущих насекомых, наносящих болезненные зудящие уколы. Это слепни, мошки, мокрецы - двукрылые разных размеров, связанные с неосвоенными территориями. Убытки от гнуса огромны, не говоря о том угнетающем действии, которое он производит на работающих, например в тайге, людей - лесозаготовителей, геологов, строителей. При строительстве, например, Братской ГЭС много усилий и средств пришлось затратить на борьбу с насекомыми-кровососами.
Еще опаснее те кровососы, которые переносят возбудителей различных заболеваний. Тут уж причиняемый ими вред исчисляется не в денежном выражении. а человеческими жизнями. До приобретения независимости в Индии от малярии, возбудитель которой передается малярийным комаром (род Anopheles), ежегодно умирало около 200 тысяч человек.
Большое снижение работоспособности людей вследствие заболевания малярией было у нас в рисосеющих районах Средней Азии до того, как малярия как массовое заболевание в СССР была ликвидирована. В тропической Америке не поддается учету количество человеческих жизней, уносимых передаваемой комарами желтой лихорадкой. В Западной Африке бичом населения является передаваемая мухой це-це сонная болезнь.
В наших широтах огромное эпидемиологическое значение имеют вши, передающие сыпной и возвратный тифы, и блохи, передающие чуму. Если наша страна не знает теперь вспышек этих болезней, то это потому, что налажен строгий контроль за этими заболеваниями и созданы санитарно-гигиенические условия, препятствующие распространению насекомых-паразитов. Огромна роль мух в переносе различных кишечных инфекций, в том числе и таких, как брюшной тиф, дизентерия и др. Только насекомыми передаются менее опасные, но тяжелые болезни, такие, как пендинская язва, лихорадка папатачи и др.
Приведенные данные показывают, насколько велика роль насекомых в природе, в экономике общества, в жизни людей, насколько необходимо всестороннее изучение огромной армии наших шестиногих друзей, союзников и врагов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'