Членистоногие Arthropoda (окончание)
Подтип Трилобиты (Trilobita) (А. Б. Ланге)
Трилобиты - вымершие морские членистоногие, особенно обильные в первой половине палеозойской эры и полностью исчезнувшие к ее концу. Панцири трилобитов находят в ископаемом состоянии в морских отложениях, их описано около 4000 видов. Трилобиты - одни из древнейших наиболее примитивных членистоногих, отличавшиеся однородностью (гомономностью) конечностей и первичным метаморфозом. Их изучение позволяет полнее понять организацию и развитие вышестоящих групп, в особенности родственных им хелицеровых.
Длина тела трилобитов - 2-10 см, редко до 75 см. У примитивных форм тело состоит из двух отделов: слитной головы и расчлененного многосегментного туловища (рис. 1). Тело заканчивается небольшой анальной лопастью (тельсон). У большинства трилобитов анальная лопасть и несколько задних сегментов сливаются, образуя третий, хвостовой отдел - пигидий. Тело делится продольными спинными бороздами на выпуклую срединную часть (ось, или рахис) и уплощенные боковые (плевры). С тройным продольным и поперечным делением тела связано название "трилобиты", что значит трехдольчатые. Покров спинной стороны очень плотный, а брюшной- тонкий перепончатый. Голова и пигидий были покрыты щитами, сегменты тела между ними соединялись подвижно, что позволяло трилобиту свертываться на брюшную сторону"
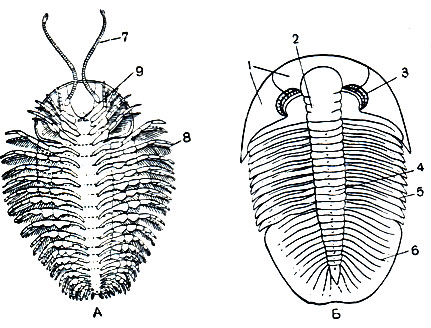
Рис. 1. Строение трилобитов: А - с брюшной, Б - со спинной стороны; 1 - головной щит; 2 - спинки сегментов головы; 3 - сложные глаза; 4 - спинки сегментов туловища (ось); 5 - плевры; 6 - пигидий; 7 - антеннулы; 8 - ноги; 9 - ротовое отверстие
В состав головы входит головная лопасть - акрон и 4 послеротовых сегмента. Акрон образует переднюю и боковые части полулунного головного щита, ему принадлежат на спинной стороне пара сложных фасеточных глаз и пара простых глаз, а на брюшной стороне - пара нитевидных членистых усиков - антеннул, прикрепленных перед ртом. Позади рта расположены 4 пары конечностей сегментов головы, которые по строению подобны друг другу и следующим за ними многочисленным ножкам туловища. Все они двуветвистые, состоят из основания и двух членистых ветвей.
Долгое время считалось, что двуветвистые ножки трилобитов и ракообразных соответствуют друг другу по строению. Однако, как установил Стермер, это не так. У трилобитов от основного членика отходит неветвистая расчлененная ножка, а вторая ветвь представлена жаброй - членистым придатком основного членика с жаберными листочками, У раков двуветвиста сама ножка. В то же время конечностям трилобитов подобны брюшные жаберные ножки водных хелицеровых, в частности мечехвостов, что служит одним из доказательств родства этих членистоногих. Ножки трилобитов, судя по их строению, выполняли различные функции: служили органами дыхания, плавания и ползания по грунту, по-видимому, гнали воду по брюшной стороне тела вперед, к ротовому отверстию, доставляя органические частицы и мелкие донные организмы, вероятно, служившие пищей трилобитам. Пища дополнительно измельчалась с помощью жевательных отростков - эндитов, имевшихся на члениках ножек у ряда форм.
Судя по ископаемым остаткам неполовозрелых стадий, развитие трилобитов представляло примитивный метаморфоз, который по некоторым признакам напоминает развитие кольчатых червей. Из яйца вылуплялась очень мелкая (до 1 мм) личинка - протаспис (рис. 2). Самые ранние известные стадии личинки имеют цельное тело. Далее оно сразу делится на головную и анальную лопасти и 4 сегмента между ними, которые являются сегментами головы трилобита и, в отличие от остальных сегментов, образуются все одновременно и называются личиночными или ларвальными. Дальнейшее развитие сопровождалось появлением туловищных сегментов, длительным ростом и линьками. Новые, послеличиночные, или постларвальные, сегменты, как и у аннелид, один за другим обособлялись вперед от зоны роста, расположенной у переднего края анальной лопасти, т. е. путем анаморфоза, ив развитии следовал длинный ряд стадий с неполным числом сегментов, вплоть до полносегментного состояния. После этого трилобит продолжал расти и линять, но новых сегментов обычно уже не появлялось, т. е. начинался период развития, называемый эпиморфозом.
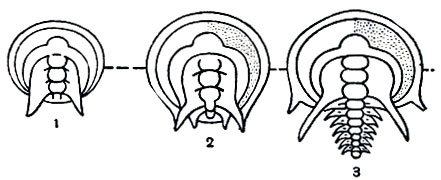
Рис. 2. Ранние стадии развития трилобитов: 1 - личинка протаспис; 2, 3 - начало образования туловищных (постларвальных) сегментов
Среди современных членистоногих первичный метаморфоз сохранился у некоторых ракообразных, у которых из яйца вылупляется личинка - науплиус, имеющая только ларвальные сегменты, а далее следует процесс наращивания постларвальных сегментов. Однако число ларвальных сегментов у протасписа и науплиуса различно: у первого 4, у второго 2. Это, а также различия в строении конечностей препятствуют сближению ракообразных с трилобитами. В то же время в эмбриональном развитии водных хелицеровых - мечехвостов - обнаружено 4 ларвальных сегмента, что в сочетании с другими признаками служит основанием для сближения хелицеровых с трилобитами.
Трилобиты были исключительно морскими формами, на что указывает характер отложений, содержащих их панцири и сопутствующие остатки других морских беспозвоночных. В большинстве трилобиты обитали в мелководных участках моря и были донными животными, медленно ползающими и плавающими у дна. На это указывает плоское снизу тело, тяжелый панцирь, относительно слабые конечности, вентральное положение рта. Многие могли зарываться в ил, особенно формы с острым краем головного щита. Обитателями толщи ила считаются некоторые формы с торчащими шипами и стебельчатыми глазами, которые они, видимо, выставляли над поверхностью дна наподобие перископов. Некоторые формы с длинными шипами на голове считают плавающими. Шипы увеличивали поверхность тела, повышая плавучесть, и такие виды, вероятно, могли парить в толще воды. Утрата глаз некоторыми группами трилобитов в одних случаях может быть связана с жизнью в иле или в сильно взмученных течениями водах в зонах илонакопления, в других - с жизнью в темноте на больших глубинах. Так, некоторые мелкие слепые трилобиты считаются обитателями больших глубин, измельчавшими в связи с недостатком пищи. Расселение трилобитов, вероятно, происходило на ранних личиночных стадиях. Мелкие личинки - протасписы, почти равные по размерам науплиусам современных веслоногих раков, очевидно, были планктонными организмами и могли далеко переноситься течениями, способствуя широкому распространению, которое характерно для многих видов трилобитов.
Трилобиты были очень многочисленны в кембрийский и ордовикский периоды, в силуре, девоне и карбоне число видов постепенно, но неуклонно сокращается, и в перми они полностью исчезают. Отчетливое изменение морфологических признаков во времени, обилие видов и хорошая сохранность панцирей сделали трилобитов важнейшей группой "руководящих ископаемых" при изучении, раннепалеозойских геологических систем. Наличие остатков трилобитов позволяет определять морские отложения, их возраст и условия образования, что важно при разведке полезных ископаемых.
При изучении трилобитов невольно возникает вопрос: что привело к полному вымиранию такой огромной группы организмов, некогда господствовавшей в морях? Обычно указывают на возможные изменения абиотических условий и на хищников. Врагами трилобитов считают ракоскорпионов, головоногих моллюсков и древних рыб, численность которых возрастала как раз с силура, а средствами защиты от хищников - наличие у многих трилобитов шипов и способность свертываться на брюшную сторону. Однако внешние факторы сами по себе вряд ли могут объяснить исчезновение трилобитов. Ведь рядом с ними жили другие группы беспозвоночных, в том числе не менее древние ракообразные, которые также подвергались действию абиотических перемен и уничтожались хищниками, но тем не менее благополучно дожили до нашего времени, а многие достигли расцвета. Видимо, дело не только в уничтожающих факторах, но и в самой природе трилобитов. Если сравнить их с ракообразными, то при всем обилии видов трилобитов бросается в глаза крайняя однотипность их примитивной организации и особенно развития. Как мы видели, формы индивидуального развития ракообразных эволюционировали с давних времен. Одни из них пошли по пути обогащения яиц желтком, благодаря чему из яйца стал вылупляться организм, сразу похожий на взрослого, у других возник сложный метаморфоз со сменой типов личинок и образа жизни, в частности у паразитов, третьи приобрели способность очень быстро размножаться, особенно мелкие планктонные формы, и т. д. Обогащение яиц желтком произошло и у водных хелицеровых, о чем речь пойдет ниже. Ничего подобного у трилобитов мы не находим. По-видимому, все они обладали первичным метаморфозом в его архаическом, так сказать, наиболее длинном варианте. Чтобы стать взрослым и способным размножаться, трилобит был вынужден пройти длиннейший ряд стадий со множеством линек, начиная от крошечного протасписа, через периоды наращивания сегментов тела и последующего роста в полносегментном состоянии. При этом, кроме самых ранних стадий, вероятно планктонных, на всех последующих образ жизни существенно не менялся, т. е. жизненный цикл был монотонным и в экологическом отношении. Естественно, что при таком способе развития противостоять уничтожающим факторам среды весьма трудно Быстрое восстановление численности вида невозможно, а сохранение хотя бы части особей в каких-то иных экологических условиях маловероятно благодаря однотипному образу жизни в течение большей части жизненного цикла. Иными словами, наиболее правдоподобно предположение, что трилобиты не смогли устоять против уничтожающих факторов изменявшейся среды, вероятно в основном против появившихся хищников, благодаря несовершенству метаморфоза, которое они так и не сумели преодолеть в своем историческом развитии,
Отметим еще, что в кембрийских отложениях открыты остатки странных членистоногих, по некоторым признакам похожих на трилобитов, а по другим на хелицеровых и на ракообразных. Эти недостаточно изученные формы рассматривают как представителей нескольких классов, которые при таком подразделении объединяются с трилобитами в подтип трилобитообразных (Trilobitomorpha). Возможно, что дальнейшее изучение этих животных позволит полнее осветить соотношения трилобитов и других членистоногих.
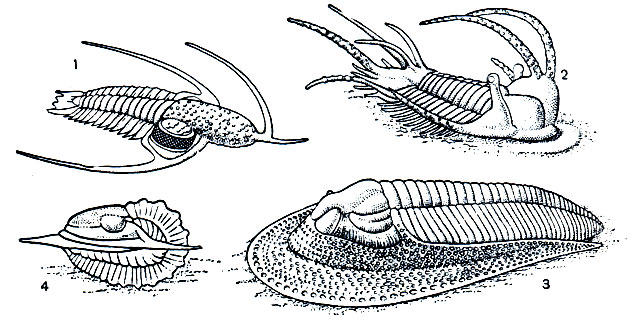
Рис. 3. Трилобиты: 1 - Lonchodomas, вероятно, плавающая форма; 2 - Ceratargus, форма с шипам и стебельчатыми глазами, вероятно, зарывавшаяся в ил; 3 - Hаrрes, типичная донная форма; 4 - свернувшийся трилобит
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'