Глава 2. Развитие психической деятельности в пренатальном периоде
Врожденное и приобретаемое в пренатальном развитии поведения
Морфофункциональные основы эмбриогенеза поведения
Поведение эмбриона является во многих отношениях основой всего процесса развития поведения в онтогенезе. Как у беспозвоночных, так и у позвоночных установлено, что развивающийся организм производит еще в пренатальном (дородовом) периоде движения, которые являются элементами будущих двигательных актов, но еще лишены соответствующего функционального значения, т. е. еще не могут играть приспособительную роль в общении животного со средой его обитания. Эта функция появляется лишь в постнатальном периоде его жизни. В таком смысле можно говорить о преадаптационном значении эмбрионального поведения.
Академик И. И. Шмальгаузен выделил несколько типов онто-генетических корреляций, среди которых особый интерес для познания закономерностей развития поведения представляют "эргонтические корреляции" (или функциональные в узком смысле), т. е. соотношения между частями и органами, обусловленные функциональными зависимостями между ними. Шмальгаузен имел при этом в виду типичные ("окончательные") функции органов. Таковыми являются уже на ранних стадиях эмбриогенеза, например, функции сердца или почек зародыша. В качестве характерного примера эргонтических корреляций Шмальгаузен указывает и на зависимость между развитием нервных центров и нервов и развитием периферических органов - органов чувств и конечностей. При экспериментальном удалении этих органов соответствующие элементы нервной системы недоразвиваются.
Как установил еще в 30-е годы Б. С. Матвеев, эмбриогенез характеризуется коррелятивными сдвигами в соотношениях между развивающимися органами, которые являются следствием нарушения взаимосвязей частей зародыша в развивающемся организме. В результате возникают функциональные изменения, определяющие характер деятельности формирующихся органов.
Эти выводы ведущих советских зоологов показывают всю сложность морфофункциональных связей и взаимозависимостей, определяющих формирование поведения животных в эмбриогенезе. Это нашло свое отражение и в концепции системогенеза П. К. Анохина. "Развитие функции, - говорил Анохин, - идет всегда избирательно, фрагментарно в отдельных органах, но всегда в крайней согласованности одного фрагмента с другим и всегда по принципу конечного создания работающей системы"*. В частности, на примерах развития эмбрионов кошки, обезьяны и человеческого плода он показал функциональную обусловленность асинхронности развития отдельных структур зародышей, подчеркнув, что "в процессе эмбриогенеза идет ускоренное созревание отдельных нервных волокон, которые определяют жизненные функции новорожденного" (например, сосательные движения), ибо для его выживания "система отношений" должна быть полноценной к моменту рождения"**.
* (П. К. Анохин. О морфологических закономерностях развития функции в эмбриогенезе животных и человека. В кн.: "Труды 6-го Всесоюзного съезда анатомов и эмбриологов", т. I. Харьков, 1961, стр. 25.)
** (П. К. Анохин. О морфологических закономерностях развития функции в эмбриогенезе животных и человека. В кн.: "Труды 6-го Всесоюзного съезда анатомов и эмбриологов", т. I. Харьков, 1961, стр. 27-28.)
Как показали исследования А. Д. Слонима и его сотрудников, внутриутробные движения влияют на координацию физиологических процессов, связанных с мышечной деятельностью, и тем самым способствуют подготовке поведения новорожденного. По данным Слонима, новорожденные козлята и ягнята способны не утомляясь бегать до двух часов подряд. Эта возможность обусловлена тем, что в ходе эмбриогенеза путем упражнения сформировалась координация всех функций, в том числе и вегетативных, необходимых для осуществления такой интенсивной деятельности уже в самом начале постнатального развития. В частности, к моменту рождения уже скоординированы регуляция минутного объема сердца и частота дыхания, как и других физиологических функций.
Говоря о принципе "конечного создания работающей системы" (Анохин) или эмбриональной преадаптации постнатального поведения, т. е. о преадаптационном значении эмбрионального поведения, нельзя забывать об относительности этих понятий. Эмбриональную преадаптацию нельзя понимать как некую изначальную, "фатальную" предопределенность. Условия жизни взрослого животного, его взаимодействие с компонентами окружающей среды в конечном итоге определяют условия развития зародыша, функционирование его развивающихся органов. Общие же закономерности и направление развития функции детерминированы сложившимися в филогенезе и генетически фиксированными факторами.
Эмбриоральное научение и созревание
В указанной связи приобретает особый интерес вопрос об "эмбриональном научении", считавшемся некоторыми исследователями преимущественным, если не единственным фактором всего сложного процесса первоначального формирования экзосоматических функций. К числу этих исследователей относится уже упоминавшийся известный американский ученый Цин-янг Куо, который еще в 20-30-х годах нашего века пытался объяснить весь процесс формирования поведения животных исключительно накоплением двигательного опыта и изменениями в окружающей зародыш среде. Куо сумел одним из первых убедительно показать, что уже в ходе эмбриогенеза происходит упражнение зачатков будущих органов, постепенное развитие и совершенствование двигательных функций путем накопления "эмбрионального опыта".
Куо изучал эмбриональное поведение на многих сотнях куриных зародышей. Чтобы получить возможность непосредственно наблюдать за движениями эмбрионов, он разработал и осуществил поистине виртуозные операции: перемещал зародыш внутри яйца, вставлял окошки в скорлупу и т. д. Ученый установил, что первые движения зародыша цыпленка, наблюдающиеся уже на четвертый, а иногда и на третий день инкубации, - это движения головы к груди и от нее. Уже через сутки голова начинает поворачиваться в стороны, и эти новые движения головы вытесняют прежние к 6-9-м суткам. Очевидно, причина тому - отставание роста шейной мускулатуры от роста головы, которая к десятому дню составляет уже более 50% веса всего зародыша. Такую огромную голову мышцы могут только поворачивать, но не поднимать и опускать. Кроме того, по Куо, движения головы формируются под влиянием таких моментов, как ее положение по отношению к скорлупе, расположение желточного мешка, сердцебиение и даже движения пальцев ног, поскольку последние во второй половине инкубационного периода располагаются слева и сзади головы.
Во всем этом Куо видел проявления действия "анатомического фактора" развития поведения. Сходным образом специфические условия морфоэмбриогенеза, анатомические изменения, связанные с ростом и развитием зародыша, влияют, по его мнению, и на формирование других движений куриного зародыша. В итоге вылупившийся из яйца цыпленок обладает целым набором выработанных в период эмбриогенеза реакций, "механизмов", но они не направлены на необходимые Для поддержания жизни раздражители. Широкая генерализация характерна, по Куо, как для раздражителей (нет избирательного отношения к ним, на весьма различные раздражители следует одинаковая реакция), так и для двигательных реакций (всегда двигается все тело, движения отдельных его частей еще плохо или совсем не координированы, они нецелесообразны, неэкономичны).
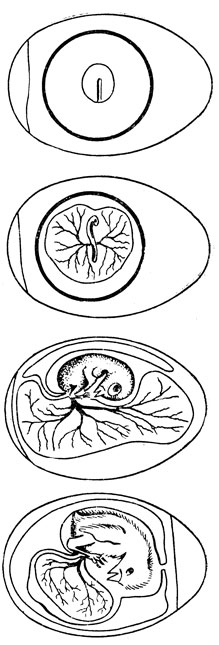
Рис. 18. Развитие куриного эмбриона. Сверху вниз: первые сутки инкубации (стадия первичной полоски - формируются зачатки головного и спинного мозга, скелета и скелетной мускулатуры), третьи сутки (зародыш на сосудистом поле желточного мешка), девятые сутки, пятнадцатые сутки
Из своих исследований Куо вывел заключение, что цыпленок должен всему научиться, что ни одна его реакция не появляется в готовом виде, а следовательно, нет врожденного поведения. Сорок лет спустя после опубликования своих первых работ Куо, уточняя свою точку зрения, указал на то, что генетически фиксированные предпосылки формирования поведения могут по-разному реализоваться в зависимости от конкретных условий развития зародыша, но важнейшую роль играет в этом процессе "расшифровки" генетической информации отношение эмбриона к окружающей его среде. Одновременно Куо подчеркивает, что эмбриональное научение не следует рассматривать в традиционном аспекте, поскольку в развитии поведения эмбриона существенную роль играет самостимулирование. Однако, как показывают результаты современных исследований, тактильная и проприоцептивная стимуляция, как представлял себе ее Куо, играет в развитии моторики в нормальном эмбриогенезе, очевидно, подчиненную роль.
Односторонний подход к проблеме становления поведения в онтогенезе, игнорирование врожденной основы индивидуального поведения, в том числе на эмбриональной стадии развития, являются, конечно, глубоко ошибочными. Если и можно говорить о некоем научении в период эмбриогенеза, то оно не происходит на пустом месте, а является развитием и видоизменением определенного генетического зачатка, воплощением и реализацией в индивидуальной жизни особи видового опыта, накопленного в процессе эволюции. Филогенез подготовил возможность развития поведения в онтогенезе в биологически полезном для особи и вида направлении. Наследственность проявляется не только в строении организма, его систем и органов, но и в их функциях.
Наследственная основа эмбриогенеза поведения выступает особенно отчетливо в тех случаях, когда элементы поведения новорожденного проявляются сразу как бы в "готовом виде", хотя возможность предшествующего "эмбрионального научения" исключается. К таким случаям, очевидно, относятся у млекопитающих поиск соска и сосущие движения новорожденных, звуковые реакции и др. Здесь можно говорить лишь о пренатальном созревании функции без эмбрионального упражнения, т. е. без пренатальной функциональной тренировки соответствующих морфологических структур. Для такого созревания явно достаточно одной лишь врожденной программы развития, возникшей и закрепившейся в ходе эволюции вида.
Хорошим примером, показывающим наличие и роль генетически фиксированных "программ действия", может служить поведение новорожденного кенгуру, который появляется на свет на столь незрелой стадии развития, что его можно сравнить с эмбрионом высших млекопитающих. В известной степени можно считать, что окончательное развитие эмбриона завершается в сумке матери. Но несмотря на состояние крайнего незрелорождения, детеныш совершенно самостоятельно перебирается в сумку матери, проявляя при этом поразительные двигательные и ориентационные способности. При этом нахождение сумки осуществляется на основе отрицательного гидротаксиса (а возможно, и хемотаксиса): оказавшись вне родовых путей, новорожденный, цепляясь за шерсть, поднимается по сухим ее участкам к сумке, находит ее вход, вползает в нее, находит там сосок, крепко присасывается к нему и остается длительное время висеть на нем. Шерсть матери смачивается перед этим на прихвостовых участках туловища родовыми водами, выливающимися после разрыва зародышевых оболочек, "дорожка" же, ведущая к сумке, остается сухой.
Здесь обращает на себя внимание строгая последовательность врожденных реакций. На эмбриональном этапе своего развития, который здесь резко укорочен, если не сказать прерван, детеныш не мог научиться ни отдельным поведенческим актам этой цепи, ни этой последовательности. До разрыва родовых оболочек он постоянно находился во влажной среде и не мог, таким образом, упражнять отрицательное гидротаксисное поведение. Никогда он не соприкасался с сухими объектами, в том числе с шерстью. Аналогично обстоит дело и с другими компонентами этого сложного поведенческого комплекса, формирование которых также невозможно объяснить "пренатальным научением".
Существенным для понимания процессов созревания элементов поведения в эмбриогенезе является то обстоятельство, что у позвоночных иннервация соматической мускулатуры туловища и конечностей предшествует замыканию рефлекторных дуг (у куриного эмбриона это замыкание происходит уже на 6-7-й день с начала инкубации). Однако сокращения этих мышц начинаются уже с момента их иннервации и носят, таким образом, на первых порах нерефлекторный характер. Эти движения являются ритмичными, так как они обусловливаются спонтанными нейрогенными ритмами ("импульсными ритмами"). Эндогенная ритмика в нервно-мышечных структурах сохраняется на протяжении всей жизни животного и представляет собой один из важных факторов созревания элементов врожденного поведения.
Все же едва ли можно говорить о некоем "чистом" созревании поведенческих актов, особенно если иметь в виду упомянутые выше коррелятивные морфофункциональные связи. К тому же, вероятно, никогда не удастся полностью исключить возможность и непосредственного пренатального упражнения тех или иных двигательных элементов поведенческого акта. Поэтому, возражая против одностороннего постулирования "эмбрионального научения" как единственного фактора пренатального развития поведения, невозможно соглашаться и с противоположной крайней точкой зрения, однобоко акцентирующей внимание на процессах эмбрионального (и постнатального) созревания. Эту точку зрения сформулировал видный американский исследователь онтогенеза поведения животных Л. Кармейкл, который допускал, что даже у человека поведение является на "девять десятых" врожденным. В том и другом случае неверным является противопоставление врожденного и приобретаемого в поведении, о чем уже многократно говорилось.
Что же касается термина "эмбриональное научение", то, очевидно, более точным будет выражение "эмбриональная тренировка", во всяком случае, когда речь идет о ранних стадиях эмбриогенеза. На первых стадиях, как уже упоминалось, даже еще нет рефлекторных дуг; сомнительно, чтобы условнорефлекторные связи образовались и на средних стадиях эмбриогенеза. Очевидно, отсутствуют на этих этапах и явления привыкания. Однако в каждом случае функционирование развивающегося органа или системы, безусловно, является индивидуальным "усвоением" и "прилаживанием" видового опыта, т. е. инстинктивного поведения, в виде тренировки. Последняя, разумеется, также относится к категории научения как одна из его элементарных форм. Полноценное же научение, как еще будет показано, встречается лишь на завершающих этапах эмбриогенеза.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'