Класс саркодовые (Sarcodina)
К классу саркодовых относятся наиболее просто устроенные среди простейших организмы. Как уже было сказано в общем очерке организации типа простейших, в подразделении его на классы существенную роль играют органоиды движения. Для саркодовых характерной формой их являются ложноножки, или псевдоподии, которые представляют собой временно образующиеся выросты цитоплазмы. Псевдоподии служат вместе с тем для захвата пищи.
Строение саркодовых, несмотря на относительную простоту их организации, отличается большим разнообразием. Главным образом это касается скелетных образований, которые, как увидим дальше, достигают у саркодовых большой сложности и совершенства.
Общее количество видов живущих в настоящее время саркодовых измеряется числом 8-10 тыс. Очень большое количество видов известно в ископаемом состоянии благодаря хорошей сохранности скелетов многих групп саркодовых.
Свыше 80% всех современных саркодовых являются обитателями моря. Некоторые виды живут в пресной воде, немногие приспособились к жизни в почве. Имеются также паразитические виды.
Класссаркодовых состоит из трех подклассов, весьма неравноценных по количеству относящихся к ним видов:
1. Корненожки (Rhizopoda) - около 2 тыс. видов;
2. Солнечники (Helizoa) -всего несколько десятков видов;
3. Лучевики (Radiolaria) - 7-8 тыс. видов.
Рассмотрим наиболее интересных и характерных представителей этих трех подклассов.
Подкласс корненожки (Rhizopoda)
Наиболее просто устроенными организмами среди корненожек являются голые амебы (Amoebina), образующие первый отряд подкласса корненожек.
Чтобы познакомиться со строением и образом жизни голых амеб, рассмотрим сначала какого-нибудь одного характерного и часто встречающегося представителя.
Амеба протей (Amoeba proteus). В пресных водах, в небольших прудах и канавах с илистым дном, нередко удается обнаружить амебу протея (Amoeba proteus). Культуру этого вида легко развести в лабораторных условиях. Амеба протей - одна из крупных свободноживущих амеб. В активном состоянии она достигает размера 0,5 мм, ее видно простым глазом. Если наблюдать под микроскопом за живой амебой (рис. 23, 24), видно, что она образует несколько довольно длинных лопастных, тупо заканчивающихся псевдоподий. Псевдоподии все время меняют свою форму, часть их втягивается внутрь, часть, напротив, удлиняется, иногда разветвляется. Тело амебы как бы переливается в псевдоподии, которые в нескольких точках прикрепляются к субстрату, и благодаря этому образование ложных ножек приводит к поступательному движению всей амебы. Псевдоподии служат не только для движения, но и для заглатывания пищи. Если псевдоподия в процессе своего образования наталкивается на какую-либо органическую частицу (водоросль, мелкое простейшее и т. п.), она "обтекает" ее со всех сторон (рис. 25) и включает внутрь цитоплазмы вместе с небольшим количеством жидкости.
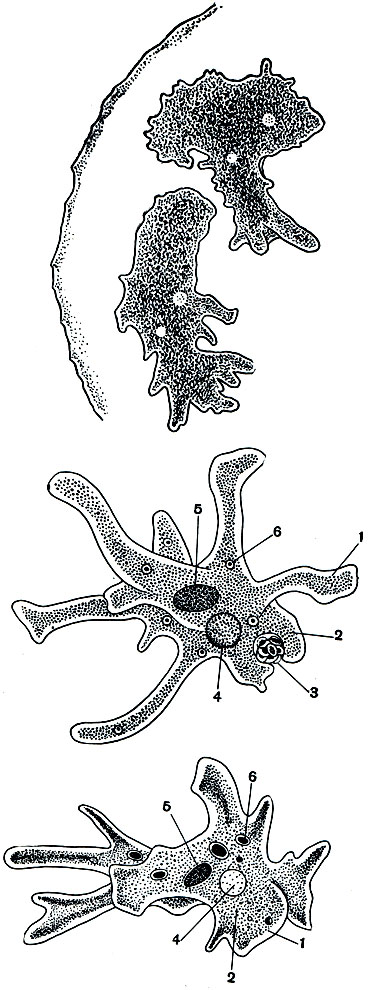
Рис. 23. Амебы в капле воды (Amoeba proteus) (вверху)
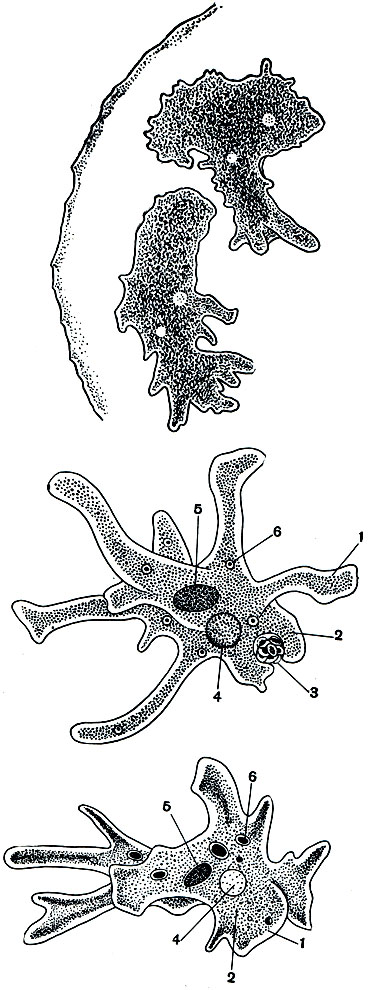
Рис. 24. Амеба протей (Amoeba proteus): 1 - эктоплазма; 2 - эндоплазма; 3 - непереваренные частицы пищи, выбрасываемые наружу; 4 - сократительная вакуоля; 5 - ядро; 6 - пищеварительная: вакуоля (внизу)
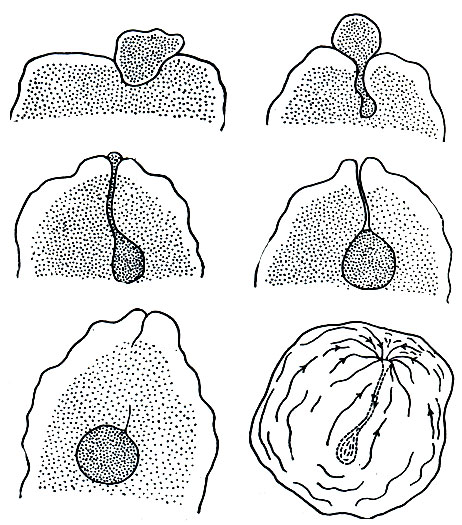
Рис. 25. Последовательные стадии заглатывания пищи амебой (Amoeba terricola)
Таким образом в цитоплазме образуются пузырьки с пищевыми включениями, которые называют пищеварительными вакуолями. В них происходит переваривание пищи (внутриклеточное пищеварение).
Не переваренные остатки пищи через некоторое время выбрасываются наружу (см. рис. 24).
Вся цитоплазма амебы ясно подразделена на два слоя. Наружный, светлый, вязкий, всегда лишенный пищеварительных вакуолей, носит название эктоплазмы. Внутренний, зернистый, гораздо более жидкий, несущий многочисленные пищевые включения, называют эндоплазмой. В состав псевдоподий входят оба слоя цитоплазмы. Эктоплазма и эндоплазма не представляют собой резко разграниченных частей тела амебы. Они могут превращаться друг в друга. В области образования и нарастания псевдоподии, куда устремляется жидкая эндоплазма, периферические части ее желатинизируются (уплотняются) и превращаются в эктоплазму.
Напротив, на противоположном конце тела протекает обратный процесс - разжижение эктоплазмы и частичное превращение ее в эндоплазму. Это явление обратимого превращения эндоплазмы в эктоплазму и обратно лежит в основе образования псевдоподий.
Кроме пищевых включений (часто сосредоточенных в пищеварительных вакуолях), в цитоплазме амебы протея обычно отчетливо бывает виден светлый пузырек, который периодически то появляется, то исчезает. Это сократительная вакуоля, играющая очень важную роль в жизненных отправлениях амебы. Сократительная вакуоля заполняется жидкостью (в основном водой), которая поступает в нее из окружающей цитоплазмы. Достигнув определенного, характерного для данного вида амеб размера, сократительная вакуоля сокращается. Ее содержимое при этом изливается наружу через пору. Весь период наполнения и сокращения вакуоли при комнатной температуре длится у амебы протея обычно 5-8 минут.
Концентрация различных растворенных органических и неорганических веществ в теле амебы выше, чем в окружающей пресной воде. Поэтому в силу законов осмоса вода проникает в протоплазму амебы. Если бы избыток ее не выводился наружу, то через короткий промежуток времени амеба "расползлась" бы и растворилась в окружающей воде. Благодаря деятельности сократительной вакуоли этого не происходит. Таким образом, сократительная вакуоля - это, прежде всего-органоид осморегуляции, регулирующий постоянно осуществляемый ток воды через тело простейшего. Однако наряду с этим она связана и с другими жизненными функциями. Вместе с выводимой из тела амебы жидкостью выводятся и продукты обмена веществ. Следовательно, сократительная вакуоля участвует в функции выделения.
Постоянно поступающая в цитоплазму вода содержит кислород. Поэтому сократительная вакуоля косвенно участвует и в функции дыхания.
Как и во всякой клетке, в теле амебы есть ядро. На живом объекте оно почти не видно. Для выявления ядра применяют некоторые красители, избирательно окрашивающие нуклеиновые вещества ядра. У амебы протея ядро довольно крупное, расположено в эндоплазме, примерно в центре тела.
Как размножаются амебы? Единственной известной у них формой размножения является деление надвое в свободноподвижном состоянии. Процесс этот начинается с кариокинетического деления ядра. Вслед за тем на теле амебы появляется перетяжка, которая в конце концов перешнуровывает тело ее на две равные половинки, в каждую из которых отходит по одному ядру. Темп размножения амебы протея зависит от условий, и прежде всего от питания и температуры. При обильном питании и температуре 20-25° С амеба делится один раз в течение 1 - 2 суток.
В пресной и морской воде живет несколько десятков видов амеб. Они различаются размерами, формой псевдоподий (рис. 26). Ложные ножки могут сильно отличаться по форме и размерам. Есть виды амеб (см. рис. 26), у которых образуется всего одна толстая короткая псевдоподия, у других - несколько длинных заостренных, у третьих - много коротких тупых и т. п. Следует отметить, что даже в пределах одного вида амеб форма псевдоподий может довольно широко варьировать в зависимости от условий окружающей среды (солевой состав, кислотность среды и т. п.).
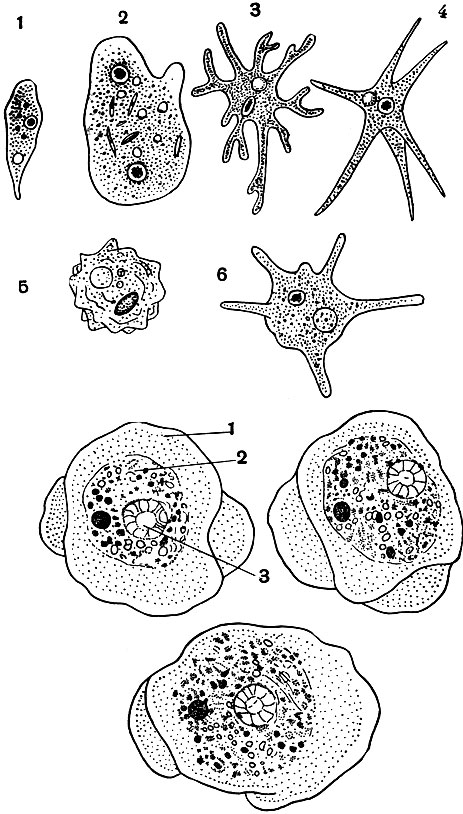
Рис. 26. Разные виды амеб с различной формой псевдоподий: 1 - Amoeba limax; 2 - Pelomyxa binucieata; 3 - Amoeba proteus; 4 - Amoeba radiosa; 5 -Amoeba verrucosa; 6 - Amoeba polypodia (вверху)
Паразитические амебы. Некоторые виды амеб приспособились к паразитическому образу жизни в кишечнике позвоночных и беспозвоночных животных. В толстых кишках человека живет пять видов паразитических амеб. Четыре вида их являются безобидными "квартирантами". Они питаются бактериями, которые в огромном количестве населяют толстую и слепую кишку человека (так же как и всех позвоночных животных), и не оказывают никакого влияния на хозяина. Но один из видов паразитирующих в кишечнике человека амеб - дизентерийная амеба (Entamoeba histolytica) - при определенных условиях может вызвать у человека тяжелое заболевание - особую форму кровавого поноса (колита), болезни, носящей название амебиаза.
Что же представляет собой дизентерийная амеба человека, почему она вызывает заболевание, как она проникает в кишечник?
Дизентерийные амебы живут в толстом кишечнике человека. Это очень мелкие (по сравнению, например, с только что описанной амебой протеем) простейшие. Размеры их - 20-30 мк. При изучении живой амебы под микроскопом ясно видно, что у нее резко разграничены эктои эндоплазма, причем зона эктоплазмы относительно широка (рис. 27). Дизентерийная амеба характеризуется очень активной подвижностью. Она образует немногочисленные короткие широкие псевдоподии, в формировании которых принимает участие почти исключительно эктоплазма.
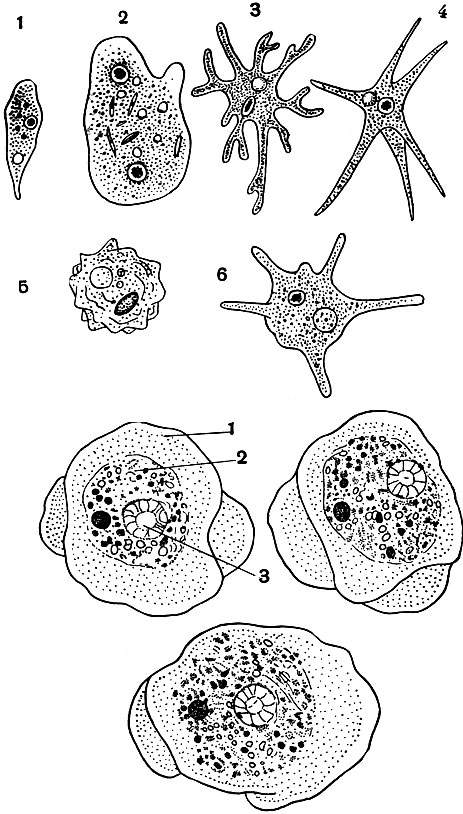
Рис. 27. Дизентерийная амеба (Entamoeba histolytica), рисунки с живого объекта в различных стадиях движения: 1 - эктоплазма; 2 - эндоплазма; 3 - ядро (внизу)
Дизентерийная амеба широко распространена по всему земному шару. В зависимости от географического положения процент зараженности людей этим паразитом варьирует в среднем от 10 до 30. Но заболевание амебиазом встречается очень редко и приурочено преимущественно к субтропическим и тропическим районам земного шара.
В умеренных и северных широтах в подавляющем большинстве случаев дело ограничивается носительством и клинически выраженные формы амебиаза представляют большую редкость.
Почему же имеется такое расхождение между частотой встречаемости паразита и частотой вызываемого им заболевания? Оказывается, дело в том, что далеко не всегда наличие дизентерийной амебы в кишечнике человека сопровождается болезненными явлениями. В большинстве случаев амеба не приносит своему хозяину-человеку никакого вреда. Она живет в просвете кишечника, активно двигается и питается бактериями. Это явление, когда возбудитель какого-либо заболевания присутствует в организме хозяина, но не вызывает патологических явлений, носит название носительства. В отношении дизентерийной амебы имеет место именно носительство.
Иногда амеба меняет свое поведение. Она активно внедряется в стенки кишечника, разрушает эпителий, выстилающий кишку, и проникает в соединительную ткань. Происходит изъязвление стенки кишечника, которое приводит к тяжелой форме кровавого поноса. Амебы, проникшие в ткани, меняют и характер своего питания. Вместо бактерий они начинают активно пожирать красные кровяные клетки (эритроциты). В цитоплазме амеб скапливается большое количество эритроцитов на разных стадиях переваривания (рис. 28). Медицине в настоящее время известны некоторые специфические лекарственные вещества, применение которых убивает амеб, что приводит к выздоровлению. Если не прибегать к лечениюг то амебиаз переходит в хроническую форму и, вызывая тяжелое истощение организма человека, иногда приводит к смертельному исходу.

Рис. 28. Дизентерийная амеба (Entamoeba histolytica): А - амеба с заглоченными красными кровяными клетками; В - амеба без эритроцитов. 1 - ядро; 2 - эритроциты
До сих пор остаются неизвестными причины, которые превращают безобидного "квартиранта" кишечника в "агрессивного" пожирателя тканей. Высказывалось предположение, что существуют разные формы дизентерийной амебы, не отличающиеся друг от друга по своему строению.
Одни из них, распространенные в умеренном и северном поясе, редко переходят к паразитизму в тканях и почти всегда питаются бактериями. Другие - южные - относительно легко становятся "агрессивными" пожирателями тканей.
Каким образом дизентерийная и другие амебы, паразитирующие в кишечнике человека, попадают в организм хозяина?
Активно подвижные формы амеб могут жить только в кишечнике человека. Будучи выведены из него, например в воду, в почву, они погибают очень быстро и не могут служить источником заражения. Заражение осуществляется особыми формами существования амеб - цистами. Посмотрим, как происходит у дизентерийной амебы процесс формирования цист. Попадая вместе с содержимым толстого кишечника в его нижние отделы и в прямую кишку, амебы претерпевают значительные изменения. Они втягивают псевдоподии, выбрасывают пищевые частицы, округляются. Затем эктоплазма выделяет тонкую, но весьма прочную оболочку. Этот процесс представляет собой инцистирование.
Одновременно с выделением оболочки цисты претерпевает изменение и ядро. Оно дважды последовательно делится, причем деление ядра не сопровождается делением цитоплазмы. Таким образом образуются столь характерные для дизентерийной амебы четырех ядерные цисты (рис. 29).
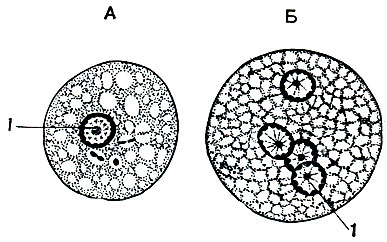
Рис. 29. Стадии инцистирования дизентерийной амебы (Entamoeba histolytica): А - одноядерная предцистная форма; Б - че-тырехъядерная циста. 1 - ядра
В таком виде вместе с фекальными массами цисты выводятся наружу. В отличие от активно подвижных вегетативных форм цисты обладают большой стойкостью. Попадая в воду или в почву, они долгое время сохраняют жизнеспособность (до 2-3 месяцев).
Подсыхание и нагревание гибельны для цист. Доказано, что цисты могут распространяться мухами, сохраняя при этом жизнеспособность.
Попадая в кишечник человека с пищей, водой и т. п., амеба эксцистируется. Ее наружная оболочка растворяется, после чего следуют два деления, не сопровождающиеся делением ядра (цисты, как мы видим, четырехъядерны). В результате получаются четыре одноядерные амебы, которые переходят к активной жизни.
Другие, не патогенные виды амеб кишечника человека распространяются таким же путем при помощи цист. По своему строению (размеры, число ядер) цисты разных видов несколько отличаются друг от друга. На этом основывается их диагностика.
Раковинные корненожки. Кроме амеб, в пресных водах встречаются представители и другого отряда корненожек - раковинные корненожки (Testacea). В море они не встречаются.
По своему строению раковинные корненожки напоминают амеб. В отличие от них часть протоплазматического тела корненожек заключена внутри раковинки, играющей роль защитного образования. В раковинке есть отверстие (устье), через которое наружу выдаются псевдоподии. У арцеллы (Arcella, рис. 30) раковинка имеет форму блюдечка. Устье ее расположено в центре. Раковинка, часто коричневой окраски, состоит из органического вещества, напоминающего по консистенции рог. Выделяется она веществом цитоплазмы подобно тому, как выделяется оболочка цисты. У диффлюгии (Difflugia, рис. 30) раковинка грушевидная. Она состоит из песчинок - мелких посторонних частичек, заглоченных, а затем отложенных на поверхности тела. У эуглифы (Euglypha) раковинка башневидная (рис. 30), но, в отличие от диффлюгии, она слагается из кремневых пластиночек правильной овальной формы. Эти пластиночки образуются в толще цитоплазмы корненожек, а затем выделяются на поверхность. Размеры раковинных корненожек невелики. Обычно они варьируют в пределах 50-150 мк.
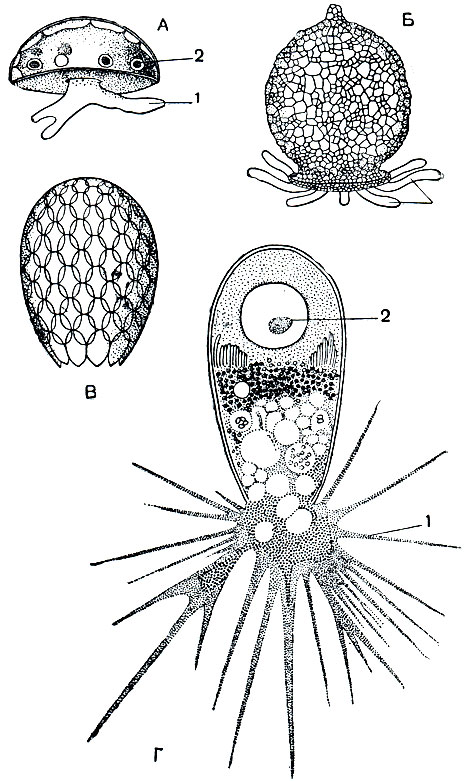
Рис. 30. Разные виды раковинных корненожек:: А - Arcella; Б - Difflugia; В - Euglypha - раковинка; Г - Euglypha с псевдоподиями. 1 - псевдоподии; 2 - ядро
Выдающиеся из устья наружу псевдоподии выполняют двоякую функцию. Они служат органоидами движения и захвата пищи. Последнее осуществляется по тому же типу, как и у голых амеб.
В связи с наличием раковины несколько видоизменяется, по сравнению с амебами, способ бесполого размножения - деления. Раковинка служит прочным скелетным образованием, и понятно, что она не может перешнуроваться пополам. Поэтому процесс деления раковинных корненожек связан с развитием новой раковины. Обычно он осуществляется следующим образом. Сначала примерно половина цитоплазмы выступает из устья. Вокруг этой части образуется новая раковинка. Одновременно с этим процессом делится ядро и одно из ядер переходит в дочернюю особь (рис. 31). На этой стадии обе особи оказываются еще связанными друг с другом мостиком цитоплазмы и обе раковинки (старая и вновь образовавшаяся) направлены одна к другой устьями. Вскоре после этого цитоплазматический мостик между особями утончается и перешнуровывается и обе корненожки переходят к самостоятельному существованию. По существу этот процесс мало чем отличается от деления амеб, он несколько усложнен лишь процессом образования новой раковинки.
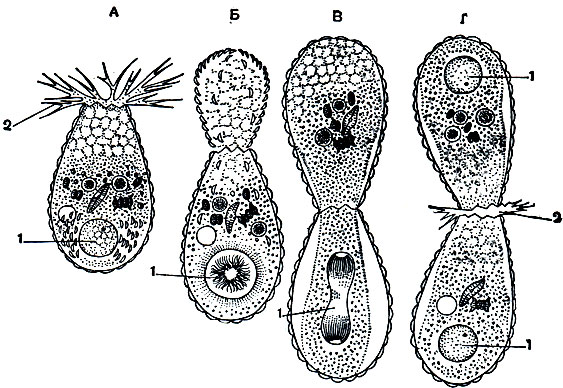
Рис. 31. Бесполое размножение путем деления раковинной корненожки Euglypha alveolata: А - корненожка перед делением; Б-образование цитоплазматической почки, на поверхности которой располагаются скелетные пластинки; В - деление ядра, скелетные пластинки образуют новую раковину; Г-конец деления, одно из ядер переместилось в дочернюю особь. 1 - ядро; 2 - псевдоподии

Таблица 2. Различные радиолярии, зарисованные с живых объектов с их естественной окраской: 1 - Acanthodesmia prismatium (отряд Nasselaria). Тонкие радиально расходящиеся псевдоподии и желтые сферические симбионты. Кремневый скелет в форме трех колец с короткими отростками; 2 - Euchitonia virchovi (отряд Spumellaria). Многочисленные тонкие псевдоподии, сетчатый трехлопастный кремневый скелет; цитоплазма окрашена в красный цвет благодаря пигменту; 3 - Auloceras arborescens (отряд Pheodaria). Коричневая центральная капсула, зеленый феодий; кремневый скелет в форме радиально ветвящихся на концах игл и поверхностно расположенных тонких иголочек (спикул); 4 - Diplocercus fuscus (отряд Acantharia). Нитевидные псевдоподии, скелет, состоящий из сернокислого стронция из радиальных неравномерно развитых игл; зеленые симбионты зоохлореллы; 5 - Arachnocorys circumtexta (отряд Nasselaria). Нитевидные псевдоподии, кремневый скелет в форме шлема с расходящимися иглами; красная центральная капсула, желтые симбионты; 6 - Tuscarilla nationalis (отряд Pheodaria). Нитевидные псевдоподии, кремневый скелет в форме конуса с отходящими от него иглами, две центральные капсулы, темно-зеленый феодий; 7 - Lithoptera miilleri (отряд Acantharia). Скелет из сернокислого стронция в форме радиальных неравномерно развитых игл с сетчатыми выростами на концах, центральная капсула крестообразной формы с зелеными симбионтами зоохлореллами; 8 -Acanthometra tetracopa (отряд Acantharia). Немногочисленные радиальные псевдоподии; скелет состоит из сернокислого стронция, слагается из 20 радиально расположенных одинаково развитых игл; цитоплазма прикрепляется к иглам при помощи сократимых волоконец (миофрисков). В центре ярко окрашенная центральная капсула с зернами пигмента и зоохлореллами
Как уже говорилось выше, раковинные корненожки - обитатели пресных вод. Они входят в состав донного населения, причем большая часть видов приурочена к прибрежной зоне. Преимущественно это обитатели мелких стоячих водоемов - прудов, канав, богатых органическими веществами.
Довольно богатая фауна корненожек (несколько десятков видов) встречается в сфагновых болотах, в самом сфагновом мху. Этот мох очень гигроскопичен и всегда впитывает большое количество воды. В прослойках воды, между стебельками и листочками мха, живут многочисленные раковинные корненожки. Здесь же встречаются и некоторые виды инфузорий.
Таким путем создается чрезвычайно характерный биоценоз обитателей сфагновых мхов.
Отряд фораминиферы (Foraminifera)
Самым обширным отрядом среди корненожек являются обитатели моря - фораминиферы (Foraminifera). В составе современной морской фауны известно свыше 1000 видов фораминифер. Небольшое число видов, представляющих, вероятно, остаток морской фауны, обитает в подпочвенных соленых водах и солоноватых колодцах Средней Азии.
Подобно раковинным корненожкам, все фораминиферы имеют раковину. Строение скелета достигает здесь большой сложности и огромного разнообразия.
В океанах и морях фораминиферы распространены повсеместно. Их находят во всех широтах и на всех глубинах, начиная от прибрежной литоральной зоны и кончая самыми глубокими абиссальными впадинами. Все же наибольшее разнообразие видов фораминифер встречается на глубинах до 200-300 м. Подавляющее большинство видов фораминифер является обитателями придонных слоев, входят в состав бентоса. Лишь очень немногие виды живут в толще морской воды, являются планктонными организмами.
Познакомимся с некоторыми наиболее характерными формами скелета фораминифер (рис. 32).

Рис. 32. Раковинки различных фораминифер: 1 - Saccamina sphaerica; 2 - Lagena plurigera; 3 - Hyperammina elongata; 4 - то же в разрезе; 5 - Rhabdam-mina linearis; в - то же в разрезе; 7 - Astrorhiza limicola; 8 - Ammo discus incertus, вид сбоку; 9 - то же со стороны устья; 10 - Cornuspira involvens; 11 - Rheopax nodulosus; 12 - Nodosaria hispida; 13 - Haplophragmoides canariensis, вид сбоку; 14 - то же со стороны устья; 15 - Nonion umbilicatulus; 16 - то же со стороны устья; 17 - Discorbis vesicularis; 18 - тоже, вид со стороны основания; 19 - Quinqueloculina seminulum (вид сбоку); 20 -то же со стороны устья; 21 - Spiroloculina depressa; 22- Textularia sagittula; 23- Globigerina sp
Среди огромного разнообразия строения раковин фораминифер можно различить по составу их два типа. Одни из них состоят из посторонних телу корненожки частиц - песчинок. Подобно тому как мы это видели у диффлюгии (см. рис. 30), фораминиферы, обладающие такими агглютинированными раковинами, заглатывают эти посторонние частицы, а затем выделяют их на поверхности тела, где они закрепляются в тонком наружном кожистом слое цитоплазмы. Такой тип строения раковины имеют часто встречающиеся представители родов Hyperammina, Astrorhiza (см. рис. 32, 3-7) и др. Например, в некоторых районах наших северных морей (море Лаптевых, Восточно-Сибирское море) эти крупные фораминиферы, достигающие 2-3 см длины, почти сплошным слоем покрывают дно.
Число видов фораминифер с агглютинированной раковиной относительно невелико (хотя число особей этих видов может быть огромным). Большая часть обладает известковыми раковинами, состоящими из углекислого кальция (СаСО3). Эти раковины выделяются цитоплазмой корненожек, которые обладают замечательной особенностью концентрировать в своем теле кальций, содержащийся в морской воде в небольших количествах (соли кальция в морской воде составляют немногим более 0,1%). Размеры известковых раковин разных видов фораминифер могут быть очень различны. Они варьируют в пределах от 20 мк до 5-6 см. Это примерно такое же соотношение размеров, как между слоном и тараканом. Наиболее крупных из фораминифер, раковина которых имеет 5-6 см в диаметре, уже нельзя назвать микроскопическими организмами. Наиболее крупные (роды Cornuspira и др.) живут на больших глубинах.
Среди известковых раковинок фораминифер в свою очередь можно различить две группы.
Однокамерные фораминиферы имеют одну-единственную полость внутри раковины, которая сообщается с наружной средой устьем. Форма однокамерных раковин разнообразна. У одних (например, Lagena) раковинка напоминает бутылочку с длинным горлышком, иногда снабженную ребрами (см. рис. 32, 2).
Очень часто происходит спиральное закручивание раковины, и тогда внутренняя полость ее становится длинным и тонким каналом (например, Ammodiscus, см. рис. 32, 8, 9).
Большинство известковых раковин корненожек является не однокамерными, а многокамерными. Внутренняя полость раковины поделена перегородками на ряд камер, число которых может достигать нескольких десятков и сотен. Перегородки между камерами не сплошные, в них имеются отверстия, благодаря чему протоплазматическое тело корненожки не расчленено на части, а представляет собой единое целое. Стенки раковинок не у всех, но у многих фораминифер пронизаны мельчайшими порами, которые служат для выхода наружу псевдоподий. Об этом подробнее будет сказано ниже.
Число, форма и взаимное расположение камер в раковинке может быть очень различным, что и создает огромное разнообразие фораминифер (см. рис. 32). У некоторых видов камеры расположены в один прямой ряд (например, Nodosaria, см. рис. 32, 12), иногда же их расположение двурядное (Textularia, см. рис. 32, 22). Широко распространена спиральная форма раковины, когда отдельные камеры расположены по спирали, причем по мере приближения к камере, несущей устье, размеры их возрастают. Причины этого постепенного нарастания размеров камер станут понятны, когда мы рассмотрим ход их развития.
В спиральных раковинках фораминифер имеется несколько оборотов спирали. Наружные (более крупные) обороты могут быть расположены рядом с внутренними оборотами (см. рис. 32,17,18) так, что все камеры видны снаружи. Это эволютный тип раковины. У других форм наружные (более крупные) камеры целиком или частично охватывают внутренние камеры (рис. 33, 1). Это инволютный тип раковины. Особую форму строения раковины находим у фораминифер милиолид (семейство Miliolidae, рис. 32, 19). Здесь камеры сильно вытянуты параллельно продольной оси раковины и расположены в нескольких пересекающихся плоскостях. Вся раковина в целом оказывается продолговатой и по форме несколько напоминает тыквенное зерно. Устье расположено на одном из полюсов и обычно снабжено зубцом.
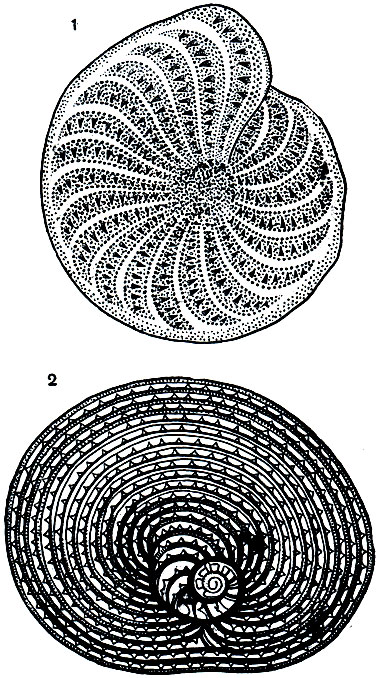
Рис. 33. Раковины фораминифер: 1 - Elphidium strigilata; 2 - Archiacina verworni
Большой сложностью строения отличаются раковинки, относящиеся к циклическому типу (роды Archiacina, Orbitolites и др., см. рис. 33, 2; 34). Число камер здесь очень велико, причем внутренние камеры располагаются по спирали, наружные же-концентрическими кольцами.
В чем заключается биологическое значение столь сложного строения многокамерных раковинок корненожек? Специальное исследование этого вопроса показало, что многокамерные раковинки по сравнению с однокамерными обладают гораздо большей прочностью. Основное биологическое значение раковинки - это защита мягкого протоплазматического тела корненожки. При многокамерном строении раковинки эта функция осуществляется весьма совершенно.
Как устроено мягкое протоплазматическое тело фораминифер?
Внутренняя полость раковины заполнена цитоплазмой. Внутри раковинки помещается и ядерный аппарат. В зависимости от стадии размножения (о чем будет сказано несколько ниже) ядро может быть одно или их несколько. Из раковинки через устье наружу выдаются многочисленные очень длинные и тонкие псевдоподии, ветвящиеся и анастомозирующие между собой. Эти особые свойственные фораминиферам ложные ножки называют ризоподиями. Последние образуют вокруг раковинки тончайшую сеточку, общий диаметр которой обычно значительно превосходит диаметр раковинки (рис. 34). У тех видов фораминифер, у которых имеются поры, ризоподии выдаются наружу через поры.

Рис. 34. Фораминифера Orbitolites complanatus с ризоподнями
Функция ризоподии двояка. Они являются органоидами движения и захвата пищи. К ризоподиям "прилипают" различные мелкие пищевые частицы, очень часто это бывают одноклеточные водоросли. Переваривание их может происходить двояко. Если частичка мала, она постепенно как бы "скользит" по поверхности ризоподии и через устье втягивается внутрь раковинки, где и происходит переваривание. Если пищевая частица велика и не может быть втянута внутрь раковины через узкое устье, то переваривание происходит вне раковинки. Вокруг пищи при этом собирается цитоплазма и образуется местное, иногда довольно значительное утолщение ризоподии, где и осуществляются процессы пищеварения.
Выполненные за последние годы с применением цейтраферной киносъемки исследования показали, что цитоплазма, входящая в состав ризоподии, находится в непрерывном движении. Вдоль ризоподии в центростремительном (к раковине) и центробежном (от раковины) направлениях довольно быстро текут токи цитоплазмы. По двум сторонам тонкой ризоподии цитоплазма как бы струится в противоположных направлениях. Механизм этого движения до сих пор остается не выясненным.
Размножение фораминифер происходит довольно сложно и у большинства видов связано с чередованием двух разных форм размножения и двух поколений. Одно из них бесполое, второе - половое. В настоящее время процессы эти изучены у многих видов фораминифер. Не вдаваясь в детали, рассмотрим их на каком-нибудь конкретном примере.
На рисунке 35 изображен жизненный цикл фораминиферы Elphidium crispa. Этот вид представляет собой типичную многокамерную фораминиферу со спирально закрученной раковинкой. Начнем рассмотрение цикла с многокамерной корненожки, обладающей маленькой зародышевой камерой в центре спирали (микросферическое поколение).
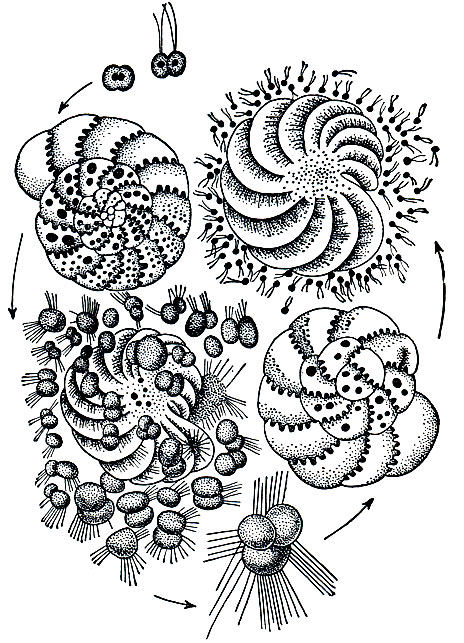
Рис. 35. Жизненный цикл фораминиферы Elphidium crispa: слева внизу - выход зародышей, образовавшихся в результате бесполого размножения; сверху-выход гамет и их копуляция
В цитоплазме корненожки первоначально имеется одно ядро. Бесполое размножение начинается с того, что ядро последовательно несколько раз делится, в результате чего образуется множество небольшого размера ядер (обычно несколько десятков, иногда свыше сотни). Затем вокруг каждого ядра обособляется участок цитоплазмы и все протоплазматическое тело корненожки распадается на множество (по числу ядер) одноядерных амебообразных зародышей, которые выходят через устье наружу. Сразу же вокруг амебовидного зародыша выделяется тонкая известковая раковинка, которая и явится первой (эмбриональной) камерой будущей многокамерной раковины. Таким образом, при бесполом размножении на первых стадиях своего развития корненожка является однокамерной. Однако очень скоро к этой первой камере начинают добавляться следующие. Происходит это так: из устья сразу выступает наружу некоторое количество цитоплазмы, которая тотчас же выделяет раковинку. Затем наступает пауза, в течение которой простейшее усиленно питается и масса протоплазмы его увеличивается внутри раковины. Затем вновь часть цитоплазмы выступает из устья и вокруг нее образуется очередная известковая камера. Этот процесс повторяется несколько раз: возникают все новые и новые камеры, пока раковина не достигнет характерных для данного вида размеров. Таким образом, развитие и рост раковины носит ступенчатый характер. Размеры и взаимное положение камер определяется тем, какое количество протоплазмы выступает из устья и как эта протоплазма располагается по отношению к предшествующим камерам.
Мы начали рассмотрение жизненного цикла Elphidium с раковинки, обладавшей очень маленькой эмбриональной камерой. В результате бесполого размножения получается раковинка, эмбриональная камера которой значительно крупнее, чем у той особи, которая приступала к бесполому размножению. В результате бесполого размножения получаются особи макросферического поколения, которые существенно отличаются от дающего им начало микросферического поколения. В данном случае потомство оказывается не вполне похожим на родителей.
Каким же путем возникают особи микросферического поколения?
Они развиваются в результате полового размножения макросферического поколения. Происходит это следующим образом. Как и при бесполом размножении, половой процесс начинается с деления ядра. Количество образующихся при этом ядер значительно больше, чем при бесполом размножении. Вокруг каждого ядра обособляется небольшой участок цитоплазмы, и таким путем формируется огромное количество (тысячи) одноядерных клеток*. Каждая из них снабжена двумя жгутиками, благодаря движению которых клетки активно и быстро плавают. Эти клетки являются половыми клетками (гаметами). Они сливаются друг с другом попарно, причем слияние затрагивает не только цитоплазму, но и ядра. Этот процесс слияния гамет и есть половой процесс. Образующаяся в результате слияния гамет (оплодотворения) клетка носит название зиготы. Она дает начало новому микросферическому поколению фораминиферы. Вокруг зиготы сразу же по ее образовании выделяется известковая раковина - первая (эмбриональная) камера. Затем процесс развития и роста раковины, сопровождающийся увеличением числа камер, осуществляется по тому же типу, как и при бесполом размножении. Раковина получается микросферической потому, что размер зиготы, выделяющей эмбриональную камеру, во много раз меньше одноядерных амебовидных зародышей, образующихся при бесполом размножении. В дальнейшем микросферическое поколение приступит к бесполому размножению и вновь даст начало макросферическим формам.
* (У некоторых видов фораминифер не все ядра участвуют в формировании гамет. Часть их остается в виде вегетативных ядер, не участвующих в процессах размножения. Этим вегетативные ядра напоминают макронуклеусы инфузорий.)
На примере жизненного цикла фораминифер мы встречаемся с интересным биологическим явлением закономерного чередования двух форм размножения - бесполого и полового, сопровождающегося чередованием двух поколений - микросферического (развивается из зиготы в результате оплодотворения) и макросферического (развивается из одноядерных амебоидных зародышей в результате бесполого размножения).
Отметим еще одну интересную особенность полового процесса фораминифер. Известно, что у большинства животных организмов половые клетки (гаметы) бывают двух категорий. С одной стороны, это крупные, богатые протоплазмой и запасными питательными веществами неподвижные яйцевые (женские) клетки, а с другой - мелкие подвижные сперматозоиды (мужские половые клетки). Подвижность сперматозоидов обычно связана с наличием у них активно двигающегося нитевидного хвостового отдела. У фораминифер, как мы видели, морфологических (структурных) различий между половыми клетками нет. По строению своему все они одинаковы и благодаря наличию жгутиков обладают подвижностью. Здесь нет еще структурных различий, которые позволили бы различать мужские и женские гаметы. Такая форма полового процесса является исходной, примитивной.
Как уже было сказано, огромное большинство современных видов фораминифер являются придонными (бентическими) организмами, встречающимися в морях всех широт от прибрежной зоны вплоть до самых больших глубин мирового океана. Изучение распределения корненожек в океане показало, что оно зависит от ряда факторов внешней среды - от температуры, глубины, солености. Для каждой из зон типичны свои виды фораминифер. Видовой состав фораминифер может служить хорошим показателем условий среды обитания.
Среди фораминифер имеются немногочисленные виды, ведущие планктонный образ жизни. Они постоянно "парят" в толще водяной массы. Типичный пример планктонных фораминифер - разные виды глобигерин (Globigerina, рис. 36). Строение их раковинок резко отличается от строения раковинок донных корненожек. Раковинки глобигерин более тонкостенные, а главное, несут многочисленные расходящиеся во все стороны придатки - тончайшие длинные иглы. Это одно из приспособлений к жизни в планктоне. Благодаря наличию игл поверхность тела, а именно отношение поверхности к массе - величина, называемая удельной поверхностью, возрастает. Это увеличивает трение при погружении в воду и способствует "парению" в воде.
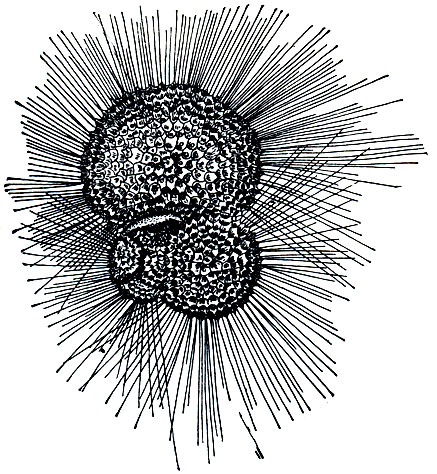
Рис. 36. Планктонная фораминифера Globigerina bulloides
Широко распространенные в современных морях и океанах фораминиферы были богато представлены и в прежние геологические периоды начиная с самых древних кембрийских отложений. Известковые раковинки после размножения или смерти корненожки опускаются на дно водоема, где входят в состав отлагающегося на дне ила. Процесс этот совершается десятки и сотни миллионов лет; в результате на дне океана образуются мощные отложения, в состав которых входит несметное количество раковинок корненожек. При горообразовательных процессах, которые совершались и совершаются в земной коре, как известно, некоторые области дна океана поднимаются и становятся сушей, суша опускается и становится дном океана. Большая часть современной суши в различные геологические периоды была дном океана. Это относится в полной мере и к территории Советского Союза (за исключением немногих северных районов нашей страны: Кольский полуостров, большая часть Карелии и некоторые другие). Морские донные отложения на суше превращаются в горные осадочные породы. Во всех морских осадочных породах присутствуют раковинки корненожек. Некоторые же отложения, как например меловые, в основной своей массе состоят из раковин корненожек. Столь широкое распространение фораминифер в морских осадочных породах имеет большое значение для геологических работ, и в частности для геологической разведки. Фораминиферы, как и все организмы, не оставались неизменными. В течение геологической истории нашей планеты происходила эволюция органического мира. Изменялись и фораминиферы. Для разных геологических периодов истории Земли характерны свои виды, роды и семейства фораминифер. Известно, что по остаткам организмов в горных породах (окаменелостям, отпечаткам и т. п.) можно определить геологический возраст этих пород. Для этой цели могут быть использованы и фораминиферы. Как ископаемые они благодаря своим микроскопическим размерам представляют очень большие преимущества, так как могут быть обнаружены в очень небольших количествах горной породы. При геологической разведке полезных ископаемых (в особенности при разведке нефти) широко используется метод бурения. При этом получается колонка породы небольшого диаметра, охватывающая все слои, через которые прошел бур. Если эти слои представляют собой морские осадочные породы, то в них при микроскопическом анализе всегда обнаруживаются фораминиферы. Ввиду большой практической важности вопрос о приуроченности определенных видов фораминифер к тем или иным осадочным породам известкового возраста разработан с большой степенью точности.
Подкласс лучевики, или радиолярии (Radiolaria)
Еще более обширную по числу видов группу морских саркодовых, чем фораминиферы, образуют лучевики, или радиолярии (Radiolaria).
Это отдельный подкласс в классе саркодовых, насчитывающий не менее 7- 8 тыс. видов. Кроме современных видов, радиолярии богато представлены и в ископаемом состоянии. Это обусловлено тем, что у большинства их, так же как и у фораминифер, имеется минеральный скелет.
Радиолярии, так же как и фораминиферы, - исключительно обитатели моря.
Все радиолярии - планктонные организмы. Жизнь их протекает в состоянии "парения" в морской воде. Наибольшее число видов радиолярий приурочено к тропическим и субтропическим водам. В холодных морях число видов их невелико.
Строение радиолярий сложно и разнообразно. Вся их организация несет ясно выраженные черты приспособления к планктонному образу жизни, которые весьма совершенны и затрагивают разные стороны строения.
Размеры радиолярий варьируют в довольно широких пределах - от 40 - 50 мк до 1 мм и более. Имеются немногочисленные колониальные формы радиолярий, размеры которых достигают величины нескольких сантиметров.
Обратимся прежде всего к рассмотрению протоплазматических частей тела радиолярий. Большинство их имеет более или менее ясно выраженную сферическую форму. Характерная особенность строения радиолярий - это наличие центральной капсулы (рис. 37).

Рис. 37. Радиолярия Thalassicola nucleata. В центре темно окрашенная центральная капсула, две зоны сильно вакуолизированной цитоплазмы (мелкоячеистая и крупноячеистая), наружу выдаются радиально расположенные тончайшие псевдоподии
Центральная капсула представляет собой мембрану, состоящую из органического вещества и окружающую центральные части цитоплазмы с ядром. Стенки центральной капсулы обычно пронизаны многочисленными мелкими порами, через которые внутрикапсулярная цитоплазма сообщается с экстракапсулярной. Центральную капсулу следует рассматривать как скелетное образование, защищающее внутренние части цитоплазмы и ядерный аппарат. У некоторых радиолярий внутрикапсулярная полость сообщается с экстракапсулярным пространством не многочисленными мелкими отверстиями, а широким отверстием, напоминающим устье раковинок корненожек.
Наружный слой цитоплазмы - эктоплазма - образует у радиолярий широкую зону. В этой зоне располагаются разнообразные включения, составляющие главную массу наружного слоя тела радиолярии. Сама цитоплазма представлена лишь тонкими прослойками между включениями. Основная масса этих включений - слизь, образующая в совокупности мощный слой, называемый калиммой. Кроме слизи, в цитоплазме радиолярий имеются и другие включения, в частности очень часто капли жира. Все эти разнообразные включения уменьшают удельный вес животного и могут рассматриваться как одна из форм приспособления к "парению" в толще воды.
У многих радиолярий в цитоплазме имеются иногда в значительных количествах зеленые (зоохлореллы) и желтые (зооксантеллы) включения. Это одноклеточные водоросли. Некоторые из этих водорослей относятся к отряда панцирных жгутиковых - Dinoflagellata. Перед нами типичный пример симбиоза простейшего животного организма с растительным. Это сожительство полезно для обоих компонентов. Водоросли получают в теле радиолярии защиту и, вероятно, некоторые питательные вещества, а также углекислоту, образующуюся при дыхании. Углекислота необходима для фотосинтеза зеленого растения. Водоросли в результате фотосинтеза выделяют свободный кислород, используемый радиолярией для дыхания. Кроме того, часть водорослей может перевариваться радиолярией, т. е. служит источником пищи. Водоросли встречаются лишь у радиолярий, живущих на небольших глубинах, куда проникает свет. У глубоководных форм они отсутствуют.
От тела радиолярии наружу отходят многочисленные тончайшие псевдоподии: (рис. 37, табл. 2 и 3), у некоторых видов анастомозирующие между собой. Они служат для улавливания пищи.
Лишь очень немногие виды радиолярий лишены скелета. У огромного большинства их имеется скелет, выполняющий двойную функцию - защитную и способствующую "парению" в толще воды. Разнообразие форм их скелетов очень велико. Многие скелеты радиолярий, имеющие часто правильную геометрическую форму, необычайно привлекательны. В скелетах радиолярий сочетается большая легкость (у планктонных организмов скелет не может быть тяжелым) с прочностью и часто с наличием разнообразных выростов, увеличивающих поверхность животного. Радиолярии - это одни из наиболее красивых и изящных организмов. Известный немецкий зоолог и дарвинист второй половины XIX в. Эрнст Геккель, бывший вместе с тем незаурядным художником, издал интересный атлас рисунков, который он назвал "Красота форм в природе". В этом обширном атласе собраны изображения множества беспозвоночных животных, которые Геккелю, как художнику, казались наиболее привлекательными с эстетической точки зрения. В атласе (имеется его русское издание) значительное место отведено радиоляриям. Геккель много занимался их изучением. Он является автором больших монографических исследований в этой области зоологии.
Мы не имеем возможности в этой книге дать подробное описание различных форм радиолярий и ограничимся лишь некоторыми наиболее интересными представителями этой обширной группы простейших (см. табл. 2 и 3).
Радиолярии обладают минеральным скелетом. У большинства он слагается из кремнезема (SiО2). В одном из отрядов радиолярий скелет состоит из сернокислого стронция (Sr2SО4).
Подкласс радиолярий состоит из 4 отрядов. Для каждого из отрядов характерны свои типичные формы скелета. Остановимся кратко на рассмотрении этих отрядов (табл. 2-3).
В отряде Spumellaria встречаются единичные виды, лишенные скелета (рис. 37), но у большинства имеется кремневый скелет. Исходная и наиболее примитивная форма его - это отдельные разбросанные в эктоплазме одноостные или трех- и четырехостные микроскопические иглы. У многих Spumellaria эти иглы спаиваются друг с другом, в результате чего получаются ажурные скелетные шары (см. табл. 3, 4,5).
Очень часто от шаров отходят радиальные иглы. У некоторых видов образуется не один, а несколько шаров, вложенных друг в друга и соединенных радиальными иглами. Вероятно, эти вложенные друг в друга шары образуются последовательно, по мере роста простейшего (процесс этот остается неизученным). Очень разнообразны кремневые скелеты в отряде Nasselaria. Исходным форматом здесь, по-видимому, является четырех лучевая спикула. Три луча ее образуют треножник, поддерживающий центральную капсулу, четвертый же направлен вверх, образуя апикальную иглу. К этой основной спикуле присоединяются кольца, соединяющие иглы спикулы (см. табл. 3, 3). Эти кольца, разрастаясь, образуют очень разнообразные и причудливые формы скелета в виде ажурных шапочек, шлемов, шаров и т. п. (см. табл. 3, 2, 3, 6). Весьма характерен и типичен скелет радиолярий, относящихся к отряду Acantharia. В химическом отношении он имеет иной состав, чем Spumellaria и Nasselaria, а именно состоит из сернокислого стронция, который довольно легко растворим в морской воде. Поэтому после отмирания животного скелет акантарий растворяется, тогда как кремневые скелеты Spumellaria и Nasselaria опускаются на дно и входят в состав ила.
Основу скелета акантарий составляют 20 радиально расположенных игл, сходящихся в центре животного (см. табл. 3, 8). Эти иглы образуют пять поясов, по 4 иглы в каждом. Их свободные концы торчат из тела радиолярии наружу. Эта исходная для Acantharia форма скелета у разных видов претерпевает разнообразные видоизменения. Иглы могут быть развиты в различной степени. Например, у Diplocercus (см. табл. 2, 4) преобладают две иглы. Очень красив скелет Lithoptera (см. табл. 2, 7). Здесь преимущественно развиты 4 иглы, на которых развиваются ответвления, образующие решетки. Весь скелет приобретает характер ажурной пластинки. Скелетные иглы акантарий прикрепляются к наружному слою цитоплазмы при помощи особых волоконец, расположенных вокруг игл. Эти волоконца способны сокращаться. При их сокращении или, наоборот, удлинении меняется общий объем протоплазматического тела радиолярии. Эти изменения представляют собой очень тонкую "настройку" для поддержания животного в состоянии "парения" в толще воды. При колебании температуры или солености плотность морской воды не остается постоянной. Увеличение или уменьшение объема тела радиолярии меняет удельный вес животного в соответствии с физическими свойствами воды.
В высшей степени разнообразны и трудно сводимы к какой-либо общей схеме кремнеземные скелеты четвертого отряда радиолярий - Pheodaria (см. табл. 2, 3, 6), куда относятся наиболее глубоководные виды. Скелет одних, аналогично Spumellaria, представляет вложенные друг в друга шары. У других имеются 2 створки, окружающие центральную капсулу. К этим элементам скелета добавляются разнообразные, иногда ветвящиеся радиальные отростки - иглы. Нередко имеется периферически лежащая зона тонких полых игл, расположенных в несколько слоев. Многообразие в строении скелета у Pheodaria поразительно!
От представителей других отрядов радиолярий представители Pheodaria отличаются тем, что центральная капсула их имеет одно или три широких отверстия, сообщающих внутрикапсулярную полость с экстракапсулярным пространством (вместо многочисленных пор, имеющихся у других радиолярий). Кроме того, в экстракапсулярной цитоплазме в области отверстия, ведущего в полость центральной капсулы, у Pheodaria имеется особое, обычно окрашенное в коричневый цвет скопление пигмента, выделительных телец и нередко пищевых включений. Эти резко выделяющиеся благодаря своему яркому цвету на фоне бесцветной цитоплазмы образования носят название феодиума. Вопрос о физиологическом значении феодиума остается неясным.
Процессы размножения радиолярий до сих пор изучены недостаточно, несмотря на то что многие ученые занимались исследованием этих интересных животных. Объясняется это в значительной мере тем, что никому еще не удалось длительное время содержать культуру радиолярий в аквариумах. Эти подлинные "дети моря" не выносят лабораторных условий существования. У некоторых крупных видов, которые имеют скелет, состоящий из отдельных игл, наблюдалось размножение путем деления надвое. У видов, обладающих сложным монолитным скелетом, такой способ размножения невозможен, так как прочный минеральный скелет не может разделиться на две половинки. По-видимому, у таких видов происходит формирование одноядерных зародышей (бродяжек), подобно тому как это происходит при бесполом размножении фораминифер.
Свойствен ли половой процесс всем радиоляриям, остается неясным. Выяснение этого вопроса требует дальнейших исследований. Дело чрезвычайно усложняется еще и тем обстоятельством, что в теле радиолярий часто живут симбиотические, а иногда и паразитические водоросли и жгутиконосцы. Эти организмы в свою очередь при размножении образуют снабженные жгутиками бродяжки, которые выходят из тела радиолярии. Не всегда легко бывает решить вопрос, имеем ли мы дело с бродяжками, принадлежащими самой радиолярии, или же с бродяжками живущих в их теле растительных организмов.
Как уже указывалось, радиолярии являются преимущественно обитателями теплых морей. Для Атлантического океана, например, установлено, что в экваториальной области число видов радиолярий из отряда Acantharia в 10 раз превышает таковое в северных районах.
Такие же примерно соотношения наблюдаются и в Тихом океане. В арктических морях радиолярий мало. Например, в Карском море их найдено только 15 видов.
Указанные закономерности в географическом распределении радиолярий справедливы для поверхностных слоев океана, температура которых определяется широтным фактором. По мере углубления в толщу водной массы различия в температурах между южными и северными широтами постепенно стираются, в связи с чем уменьшаются и различия в фауне радиолярий.
Кроме отчетливо выраженной зависимости распределения радиолярий от широтного фактора, что связано в первую очередь с температурой, у них отчетливо выражена и вертикальная зональность. Этот вопрос был изучен, например, довольно подробно в области Курило - Камчатской впадины Тихого океана, где глубины достигают 10 тыс. м. Среди радиолярий можно различить 2 группы видов. Одна не приурочена или слабо приурочена к какой-либо определенной глубине и встречается в разных глубинных зонах. Такие виды называют эврибатными. Другие, напротив, более или менее характерны для определенной глубины - это стенобатные формы. В области Курило - Сахалинской впадины было обнаружено довольно много таких стенобатных видов, причем некоторые из них были найдены лишь на глубинах, превышающих 4000 м.
Для стенобатных видов радиолярий Курило - Камчатской впадины Тихого океана их приуроченность к определенным глубинам совпадает с приуроченностью к определенным температурам. Наиболее глубоководные (абиссальные) виды живут при постоянной температуре 1,5-2,0° С. Среднеглубинные виды распространены в относительно теплой водной массе с температурой 3,5° С. Виды, приуроченные к небольшим глубинам, заселяют характерный для этой области океана холодный слой воды с температурами около 0° С. Наконец, виды, живущие в поверхностных слоях, подвергаются сезонным колебаниям температуры (средняя годовая температура этого слоя 2,6° С).
В Неаполитанском заливе Средиземного моря русский ученый проф. В. Т. Шевяков проводил в течение нескольких лет наблюдения за распределением радиолярий. В частности, оказалось, что радиолярии отряда Acantharia чрезвычайно чувствительны к малейшим изменениям условий среды, в том числе, например, к опреснению. Обычно акантарии распределены преимущественно в поверхностных слоях моря. Однако после сильных дождей они "спасаются" от опреснения и опускаются на глубины 100-200 м. Через 1-2 суток акантарии вновь поднимаются в поверхностные слои.
Acantharia оказались очень чувствительными и к волнению. При сильной волне они уходят на глубину 5-10 м. В зимние месяцы в связи с похолоданием поверхностных слоев моря акантарии также опускаются на глубины 50-200 м, где температура воды выше.
Среди четырех рассмотренных выше отрядов радиолярий Acantharia являются по преимуществу обитателями поверхностных слоев моря. Три остальных отряда (Spumellaria, Nasselaria, Pheodaria) в большей своей части приурочены к глубинным частям.
Из морей, омывающих берега Советского Союза, наиболее богаты радиоляриями дальневосточные моря. Внутренние моря (Каспийское, Азовское) совершенно лишены радиолярий. Это связано, очевидно, с их опреснением по сравнению с Мировым океаном. В северных морях как европейской, так и азиатской части СССР радиолярии очень не многочисленны, что связано с преобладающими здесь низкими температурами.
Радиолярий нередко находят и в ископаемом состоянии в осадочных морских породах. В ископаемом состоянии известны представители двух отрядов - Spumellaria и Nasselaria. Скелет Acantharia, состоящий из сернокислого стронция (SrSO4), в ископаемом состоянии не сохраняется, так как довольно легко растворяется в воде. Ископаемые Pheodaria не описаны.
Ископаемые радиолярии встречаются в осадочных породах различного геологического возраста, начиная с кембрийских отложений. Это говорит о том, что радиолярии представляют собой очень древнюю группу животного мира. Существуют некоторые древние осадочные породы, в которых среди прочих органических остатков скелеты радиолярий преобладают, составляя основную массу их (такие породы получили название радиоляриты). На территории Советского Союза радиоляриты известны в силурийских и девонских отложениях Урала, в Западной Сибири, на Дальнем Востоке (в пермских отложениях Сихотэ-Алиня). Остров Барбадос (Карибское море), входящий в группу Малых Антильских островов, в основном слагается из трепела - породы, состоящей из скелетов радиолярий. На острове есть гора высотой 360 м, построенная из трепела. Здесь найдено свыше 200 видов радиолярий, причем скелеты их очень хорошо сохранились.
Радиолярии, кроме пород, состоящих преимущественно из их скелетов, присутствуют в различных количествах во многих других осадочных морских породах (известняки, мел, сланцы и т. п.).
В связи с таким широким распространением в морских отложениях радиолярии наряду с фораминиферами играют важную роль при определении возраста горных пород.
Подкласс солнечники (Heliozoa)
Третий подкласс класса саркодовых - это солнечники (Heliozoa). Сюда относится всего несколько десятков видов простейших, часть которых обитает в пресной, часть в морской воде. Большинство солнечников - свободноплавающие в толще воды организмы, лишенные минерального скелета. Некоторые виды ведут прикрепленный к субстрату образ жизни.
В небольших пресноводных водоемах - прудах, канавах - в летнее время нередко можно встретить солнечника Actinosphaerium eichhorni (рис. 38).
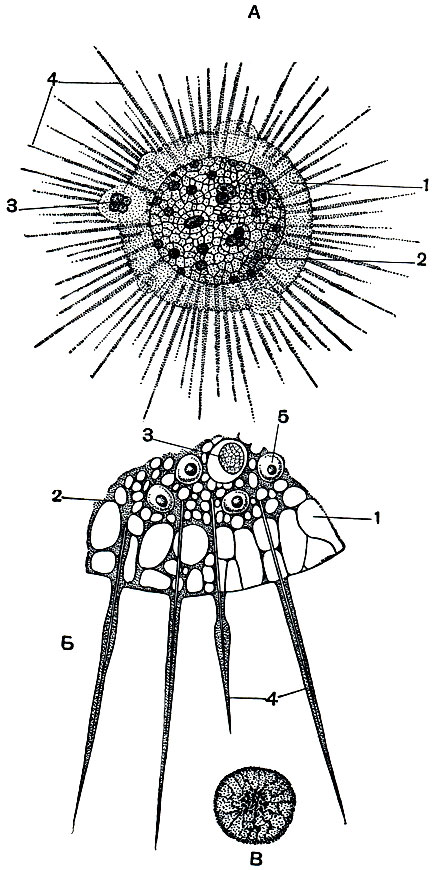
Рис. 38. Солнечник Actinosphaerium eichhorni: А - общий вид; Б - участок тела при большем увеличении; В - строение ядра при большом увеличении. 1 - эктоплазма; 2 - эндоплазма; 3 - пища; 4 - аксоподий; 5 - ядро
Тело Actinosphaerium шарообразно, достигает в диаметре 1 мм, так что его хорошо видно простым глазом. Многочисленные тонкие совершенно прямые псевдоподии расходятся по радиусам во все стороны. Постоянство формы псевдоподий обусловлено тем, что внутри них проходит прочная эластичная осевая нить. Такой тип строения псевдоподий носит название аксоподий.
Цитоплазма солнечника имеет ячеистую структуру и распадается на два слоя: наружный, более светлый, - корковый (эктоплазма) и внутренний - сердцевинный (эндоплазма). Эктоплазма занимает примерно 1/6-1/10 всего диаметра тела животного. В этом наружном слое вакуоли особенно крупны и заполнены прозрачным содержимым. Среди них выделяются по своим размерам две (а иногда и большее количество) особенно крупные вакуоли, которые периодически сокращаются. Это сократительные вакуоли. Частота их сокращения при комнатной температуре около минуты.
Внутренний слой цитоплазмы (эндоплазма) обладает гораздо более мелкой ячеистостью, чем эктоплазма, и заполнен обычно многочисленными пищевыми включениями. Здесь происходит внутриклеточное пищеварение. Не переваренные остатки пищи выбрасываются наружу через слой эктоплазмы. Существенным отличием солнечников от радиолярий является отсутствие у них центральной капсулы. Каким образом солнечник ловит добычу и чем он питается? Добыча улавливается псевдоподиями. Если какое-либо мелкое животное или растение прикоснется к псевдоподии, то оно тотчас же прилипает к ней и очень скоро теряет способность двигаться. Вероятно, животных парализует какое-то ядовитое, выделяемое цитоплазмой солнечника вещество. К захваченной добыче склоняются соседние псевдоподии (аксоподии), и она их совместными движениями подводится к поверхности коркового слоя цитоплазмы. Нередко навстречу пище со стороны эктоплазмы образуется временная короткая тупая псевдоподия, способствующая захвату ее. В эктоплазме пища не задерживается, а тотчас же переходит в эндоплазму, где и переваривается.
Пищевые объекты солнечников разнообразны. Это могут быть различные простейшие (инфузории, жгутиконосцы), но также и мелкие многоклеточные животные (например, коловратки, мелкие ресничные черви).
Ядерный аппарат солнечника Actinosphaerium состоит из множества одинаковых ядер (у крупных особей число их может достигать 200 и даже более), всегда расположенных только в эндоплазме. Actinosphaerium - пример многоядерного простейшего.
Солнечники могут вести себя в водоеме различным образом. Часто они "парят" в придонном слое толщи воды. Кроме того, они могут передвигаться и по субстрату, как бы катиться по дну, медленно изгибая при этом аксоподии.
Размножаются солнечники Actinosphaerium делением. Тело солнечника при этом перетягивается на две более или менее одинаковые половинки. Этому предшествует деление ядер, которое, однако, не протекает одномоментно, а растягивается на весь промежуток времени между двумя делениями солнечника.
Кроме бесполого, у солнечников имеется и половое размножение, протекающее довольно сложно.
Кроме крупного Actinosphaerium, в пресных водах встречаются и другие, более мелкие виды солнечников, близких по строению к Actinosphaerium. Укажем, например, на часто встречающихся мелких Actinophrys sol, обладающих, в отличие от Actinosphaerium, лишь одним ядром. В море тоже встречаются солнечники, по своему строению напоминающие Actinosphaerium, например многоядерные Cаmptonema nutans.
Среди солнечников, как пресноводных, так и морских, имеются некоторые виды с минеральным скелетом из кремнезема, подобно радиоляриям. Примером может служить пресноводная Clathrulina elegans (рис. 39). Это небольшой солнечник с типичными радиальными аксоподиями, тело которого окружено тонким скелетом в форме ажурного шарика. При помощи тонкой ножки организм этот прикрепляется к субстрату. В эндоплазме имеется одно ядро. При размножении Clathrulina протоплазма делится внутри скелета на две дочерние особи, которые выходят наружу через отверстие в скелете, а затем образуют свой новый скелет и стебелек.
Рис. 39. Солнечник Clathrulina elegans: 1 - аксоподии; 2 - скелет; 3 - дочерние особи внутри скелета материнской особи
В процессе эволюции саркодовые, приспосабливаясь к различным средам обитания, дали большое разнообразие форм. Организация саркодовых еще очень примитивна. На примере амебы мы видим одного из наиболее просто устроенных животных организмов. Но и в пределах примитивной организации саркодовые проделали большой эволюционный путь от амебообразных организмов к сложно устроенным фораминиферам и радиоляриям. Однако усложнение строения в пределах класса саркодовых лишь в относительно малой степени затронуло живые протоплазматические части клетки. Органоидами движения и захвата пищи на всех этапах эволюции саркодовых остаются псевдоподии. Зато большого прогрессивного развития достигают в пределах саркодовых скелетные элементы, выполняющие, с одной стороны, защитную и опорную функцию, а с другой - способствующие "парению" в воде (увеличение удельной поверхности), что имеет важное биологическое значение, так как многие морские саркодовые - планктонные организмы.
Развитие сложных скелетных образований сыграло большую роль и в эволюции донных (бентических) саркодовых - фораминифер. Развитие сложного и разнообразно устроенного скелета обеспечило фораминиферам биологическое процветание как в прежние геологические эпохи, так и в современной морской фауне.
Наконец, относительно небольшое число видов саркодовых избрало в качестве среды обитания другие организмы и стало паразитами. В отношении сложности строения паразитические саркодовые остались на уровне голых амеб.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'