Филогения и геологическое прошлое животного мира
Как мы уже указывали, система живых существ неразрывно связана с их филогенией. Это две стороны одного и того же явления: система - это статическое состояние современного животного мира, филогенетика дает представление о процессе становления современных таксономических подразделений и тем самым об их родственных связях. Естественная система без филогении, так же как и филогения без естественной системы, теряет свой реальный смысл.
В прилагаемой схеме филогенетического развития животных (рис. 1) отражены современные представления. Многие части этой схемы достаточно общепризнанны и не вызывают сомнений. К ним можно отнести возникновение губок и кишечнополостных от одноклеточных животных, возникновение низших червей (турбеллярий) от кишечнополостных и немертин от низших червей (турбеллярий). Несомненно возникновение членистоногих от кольчатых червей (морских кольчецов - полихет).

Рис. 1. Схема филогенетического развития животных
Также общепризнанно близкое родство групп, относящихся к вторичноротым. В остальном филогенетические связи более или менее спорны. Прежде всего вызывает сомнения генезис губок. Еще более неясны происхождение и генетические связи олигомерных групп пер-вичноротых - форонид, мшанок и плеченогих, ошибочно объединявшихся иногда термином Tentaculata (щупальцевые) или Prosopygii (переднепорошицевые).
Развитие эмбриологии, сравнительной анатомии, экспериментальной генетики и сравнительной биохимии будет способствовать построению достаточно достоверной филогенетической схемы. Много очень важных данных для построения филогении дает также и палеонтология. Так, отпечатки трилобитов и их четырехсегментных личинок дали возможность сблизить их с классом хелицерат и приблизить к ним представителей класса мечехвостов (Xiphosura), имеющих трилобитообразную четырехсегментную личинку. Самые древние иглокожие карпоидеи (Carpoidea) были уже сидячими организмами (вели прикрепленный образ жизни), но сохраняли еще двустороннюю симметрию. Это одно из доказательств, что предками современных иглокожих были двусторонне-симметричные организмы. Генетические взаимоотношения внутри типа моллюсков, особенно головоногих, смогли быть разъяснены благодаря богатейшим ископаемым остаткам вымерших групп. Но, конечно, наиболее замечательные факты дает палеонтология для суждения о генеалогии современных групп позвоночных (группа кистеперых рыб). Это касается в равной степени и вопроса о возникновении наземных позвоночных, происхождении рептилий, птиц и млекопитающих.
В филогенетических построениях очень большое значение имеет принцип эволюционного развития, который лежит в их основе (принцип монофилии или полифилии). Большинство зоологов придерживаются дарвиновского принципа монофилии, однако имеются и сторонники полифилии.
Принцип монофилии требует единого корня происхождения как для всего животного мира, так и для каждой систематической группы,
Полифилитическая концепция, наоборот, допускает возможность возникновения отдельных систематических групп животных, как и всего животного мира, из нескольких корней путем конвергентного схождения. Эта концепция находит свое применение в палеонтологии.
Борьба двух концепций идет и по вопросу о происхождении человека. Советские антропологи, так же как и многие ученые за рубежом, считают, что все человеческие расы произошли от одной предковой группы. Некоторые зарубежные антропологи склонны искать разные корни вида Homo sapiens - азиатские, африканские, австралийские.
В истории выработки естественной системы и филогении таких примеров можно было бы привести очень много. Всего 150 лет назад великий французский специалист по сравнительной анатомии Ж. Кювье объединил фораминифер и моллюсков за внешнее сходство в строении раковины, присоединяя к ним и усоногих ракообразных, что уже не соответствует современным представлениям.
Вся история современной системы животных - это стремление к естественной системе и утверждение принципа монофилии.
Принятая в настоящем издании филогения животных построена по монофилитическому принципу.
Геология дает возможность расположить в хронологическом порядке время появления, наибольшего развития и вымирания многих групп животных, остатки которых сохраняются в ископаемом виде (рис. 2). Прежде чем рассмотреть эту схему, следует сделать некоторые предварительные замечания.
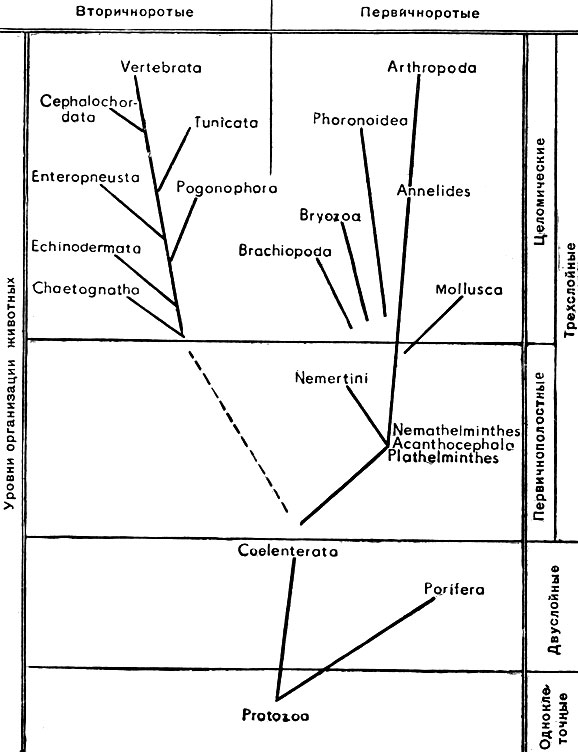
Рис. 2. Филогенетические связи некоторых групп животных
Более или менее достоверная пелеонтологическая летопись застает животный мир не на первых этапах его развития, а в довольно зрелом состоянии. Период с кембрия до наших дней - это, возможно, только одна десятая всего времени развития на Земле животных. При этом следует иметь в виду, что было бы весьма ошибочно переносить представления о скорости развития наземной фауны на все животное царство, а именно на гораздо более богатое типами и классами животное население морей и океанов, иначе говоря, галосферы*. Темп филогенетического развития подтипов наземных членистоногих - трахеат и хелицерат (классы многоножек, насекомых и паукообразных) - и подтипа наземных позвоночных (большинства амфибий, рептилий, птиц и млекопитающих), а также некоторых других небольших групп почвенных и пресноводных беспозвоночных (саркодовых, жгутиковых, инфузорий, турбеллярий, коловраток, олигохет, моллюсков, ракообразных и некоторых других) в истории развития животного мира занимает особое место и представляет собой бурный процесс заселения не заселеной до того наземной и пресноводной среды выходцами из океана. Если мы перенесем темпы развития с наземной фауны на морскую, то получим значительное несоответствие. Совсем другими темпами характеризуется развитие морских животных, включающих 15 типов фауны. Пока суша не была заселена растительностью, на ней было невозможно и развитие жизни животных. Только в силурийское время у морских побережий стали возникать предковые для наземной растительности формы (псилофитовая флора) и быстрым темпом стала развиваться наземная растительность. Что же касается морей и океанов, то в них хлорофилл констатирован в отложениях, отстоящих от нас на 31/2 млрд. лет. В силурийское время у морских побережий и в сильно опресненных прибрежных водах уже подготовлялись "десантные части" для захвата воздушной среды и пресных вод суши; это были членистоногие, некоторые группы червей и позвоночные. Морфофизиологические особенности ряда групп морской фауны и огромное пространство расположенной рядом суши, предоставившей к тому времени первичные (растительные) пищевые ресурсы, создали условия для бурно развернувшегося и протекавшего быстрыми темпами процесса видообразования, на первых этапах лишенного острой конкуренции. В этом процессе ярче всего сказалась свойственная живым организмам характерная способность расширять свой жизненный ареал и захватывать новые среды обитания в пределах возможного для их морфологических и физиологических структур.
* (Галосфера - морские воды, от греческого halos - соленый.)
Что касается морской фауны, то можно с полной уверенностью констатировать, что из нижнего кембрия уже известны многочисленные представители современной морской фауны. В этой истории мы совсем не знаем каких-либо катастрофических смен фауны, о которых часто, со времен Ж. Кювье, говорят специалисты по геологической истории наземных рептилий и позвоночных.
Эволюция морской фауны протекала за палеонтологическое время (с начала палеозоя) очень медленно и спокойно. Сколь относительно медленны темпы эволюции морской фауны, можно видеть на следующих примерах.
Близкий к замечательному моллюску Neopilina ископаемый род Tryblidium существовал в кембрии 400-500 миллионов лет тому назад, головоногие моллюски наутилиды известны с юры (свыше 100 миллионов лет) и дожили до нашего времени, род Limulus (мечехвост) известен с верхней перми (около 200 млн. лет), плеченогие Lingula и Crania известны с девона (250-300 млн. лет). Кистеперые рыбы, к которым относится замечательная латимерия, впервые пойманная в живом виде у южных берегов Африки, а затем у Коморских островов, также имеют возраст 250-300 млн. лет. Такие обычные ныне роды двустворчатых моллюсков, как Nucula, Leda, Modiolus, Ostrea, Lima, Avicula и др., существуют с кембрия, т. е. 400-500 млн. лет. Симпсон в своей известной книге "Темпы и формы эволюции" (1944) определяет длительность существования некоторых родов двустворчатых моллюсков не менее как в 275 млн. лет, а некоторых - более 400 млн. лет. Средний геологический возраст для родов двустворчатых моллюсков он определяет в 80 млн. лет. Наряду с этим можно упомянуть, что пресноводное ракообразное щитень (Apus cancriformis) существует с триаса, т. е. свыше 150 млн. лет, с теми же родовыми признаками.
Таким образом, можно считать несомненным положение, что с кембрия по наше время морская фауна не претерпела (по сравнению с наземной) каких-либо очень крупных принципиально важных изменений и находилась примерно на том же этапе эволюционного развития, что и ныне. Во всяком случае, все типы и большинство классов морских животных уже существовали. Если представить себе длительность всего исторического развития современной морской фауны, то период с начала кембрия до наших дней должен рассматриваться только как последний короткий этап. Если при этом допустить, что темп эволюции в докембрийское время не мог существенно отличаться по своему характеру и длительности от послекембрийского, то необходимое время для предшествующей кембрию эволюции морской фауны должно было во много раз превышать послекембрийское.
Геологическая история животных вскрывает еще одно явление исключительного интереса - это процесс вымирания.
Можно сказать, что почти в наше время (в конце XVII в.) вымерла птица дронт (или додо) на Маврикиевых островах, а у нас на Дальнем Востоке - морское млекопитающее стеллерова (морская) корова (во второй половине XVIII в.).
В настоящее время почти полностью могли бы быть уничтожены такие ценные животные, как морские котики, калан, (морская выдра) и другие, если бы человек не взял их под охрану. Исчезли дикие лошади, коровы и т. д. Все эти случаи- результат встречи с человеком, его промыслом или бытом.
Но геология знает другие более интересные примеры вымирания, в которых человек не играл никакой роли: трилобиты, аммониты, наутилоидеи, белемниты, некоторые классы иглокожих, панцирные земноводные (стегоцефалы), птеродактили и др. Все это уже вымирание не отдельных видов, а целых больших групп и очень важных элементов фауны, в состав которых входили сотни, а иногда и многие тысячи видов. В современной фауне имеются резко обособленные систематически и морфологически группы, представленные несколькими видами, являющимися осколками древних богатых видами групп. Упомянутая выше Neopilina, представленная ныне тремя видами в глубинах восточной стороны Тихого океана,- это, вероятно, последний осколок, может быть, многочисленного в палеозое и в допалеозойское время класса однокрышечковых моллюсков (Моnоplacophora). Целый класс Priapuloidea представлен в настоящее время всего 5 видами, но его остатки обнаружены в палеозойских отложениях. Вероятно, это была в прошлом обильная видами группа животных, но отсутствие скелетных частей стерло из прошлого их остатки. Замечательная группа погонофор, ранга типа, блестяще описанная А. Ивановым, представлена в наше время несколькими десятками глубоководных видов древнего облика, но в прошлом также, вероятно, имела довольно широкое распространение.
Удивительный существующий ныне осколок древних гигантострак (из хелицеровых членистоногих) - мечехвосты (Limulus) существуют в количестве 5 видов, также как остаток весьма многочисленной группы Nautiloidea из головоногих моллюсков, существующий ныне в числе 4 видов рода Nautilus. Подобных примеров можно было бы привести немало, и в каждом случае можно высказать подозрение: не вымирающие ли это организмы? Тем более удивительно, что немногочисленные остаточные виды богатых видами вымерших групп часто производят впечатление благоденствующих ныне форм, как, например, Nautilus, Limulus, Priapulus.
Как объяснить причины их вымирания? Прежде всего, конечно, конкуренцией с другими, более молодыми и биологически более сильными группами. Для трилобитов это могли быть ракообразные, для головоногих моллюсков - рыбы и киты, для птеродактилей - птицы и т. п. Однако Только конкуренцией не объяснить всего явления вымирания.
Помимо конкуренции между видами и отдельными особями, вымирание диких животных объясняется резкой сменой природных условий и невозможностью для этих животных быстро приспособиться к новым условиям среды. Мамонт вымер, вероятно, одновременно с исчезновением оледенения в северных областях Сибири и Америки и в значительной степени под воздействием человека. Возможно, что основная причина вымирания животных часто кроется в сочетании этих двух факторов.
Геологическое прошлое Земли и геологическая хронология как бы подразделяются на две части - докембрийский период и от кембрия до наших дней. Как хронология, так и животные послекембрийского периода изучены значительно лучше, чем докембрийского.
На прилагаемой схеме дана геологическая история различных групп животных, известных по ископаемым остаткам, которая сопоставлена с историей некоторых групп растений, начиная с кембрийского периода (см. рис. 1). Как видно, весь этот период укладывается всего лишь в 500-600 млн. лет. Точность знаний об отдельных эпохах этого периода и его хронологии тем меньше, чем дальше вглубь веков уходит его история. Докембрийское время, которое еще недавно исчислялось учеными сотнями миллионов или в крайнем случае одним миллиардом лет, в настоящее время все более и более растягивается. Крупнейший специалист по докембрию академик А. А. Полканов дает следующую хронологическую схему для докембрия Северной Европы.
| Периоды и эпохи | Абсолютный возвраст в млн. лет |
|---|---|
| Протерозой | |
| Верхний | 1670 - 715 |
| Средний | 1880 - 1750 |
| Нижний | |
| Архей | |
| Верхний | 2140 - 1900 |
| Нижний | 2700 - 2200 |
| Катархей | 3590 - 2710 |
Очевидно, это тот период в истории Земли, когда земная кора уже была по температурному режиму доступна для развития жизни. Академик А. П. Виноградов тоже считает, что температурные условия на поверхности Земли приближались к современным в течение 2-3 млрд. лет, а возраст земной коры он определяет в 5 млрд. лет. Академик О. Ю. Шмидт допускал время существования твердой оболочки Земли в 7 млрд. лет.
Когда и как появилась жизнь на нашей планете, остается неясным, но, в частности, можно допустить, что возникновение фотосинтетической деятельности растительных организмов на Земле надо относить к периоду, отдаленному от нас на 3-4 млрд. лет. Уже давно стало очевидно, что остатки растений и животных из докембрийских отложений редки, спорны и неясны. Создалось мнение, что все эти остатки подверглись перекристаллизации (метаморфизации) и как документы науки утеряны.
Существует также допущение, что отсутствие ископаемых организмов в докембрии объясняется тем, что докембрийские животные не имели скелетов. Допускают также, что основные группы палеозойских животных сформировались на рубеже кембрия. Однако все эти допущения не обоснованы в нужной мере, и вопрос о ничтожном количестве докембрийских ископаемых остатков животных пока остается без ответа. См. (Таблица (рис. 1)) поясняет то, что было сказано в предыдущем изложении. В самом начале кембрия мы застаем уже все основные группы современных морских животных, имеющих скелеты, за исключением рыб: моллюски, ракообразные, мшанки, плеченогие, иглокожие. На этой схеме хорошо видно, как растения и животные выбирались на сушу и на сколько наземные животные моложе морских.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'