Зоогенная регуляция элементов структурной организации эфедро-саксаулового биогеоценоза в условиях песчаной пустыни
Статическая картина морфологии структуры биогеоценоза обычно индицируется топографией растительного покрова и его ярусной расчлененностью. Функциональная же обусловленность элементов структуры, а в экстремальных зонах и сам тип структурной организации биогеоценоза и циклы ее динамических трансформаций, выявляющихся нередко в средообразующем эффекте, определяются деятельностью животных, которые в условиях аридных зон выступают в роли важнейших и активных биогеоценотических агентов, регуляторов и нередко лимитирующих факторов. Этот феномен назван "зоогенной регуляцией в биогеоценозе" (Залетаев, 1973).
К категории этого явления относятся как природные процессы, которые от начала до конца развиваются "под контролем" животных, так и те процессы, которые возникают и инициируются животными в их начальной стадии, а затем в механизме их развития ведущая роль передается растениям, микроорганизмам или комплексу биотических и абиотических факторов. Таковы зооэоловые процессы (Залетаев, Зубкова, 1968), зоофитоэоловые и различные формы консервации органических веществ в природе.
В условиях биогеоценоза черносаксаулового леса с участием эфедры шишконосной, типичного для гипсоносных песков пустыни Каракум, зоогенная регуляция оказывается условием, определяющим в сочетании с фитогенным засолением и эоловыми процессами формирование типичного сложнобугристого рельефа поверхности песков, закономерную смену стадий в циклическом развитии биогеоценотической структуры и лимиты "экологического возраста" черносаксаулового леса.
Главнейшими агентами зоорегуляции в функциональных узлах этого зонально типичного для южных пустынь биогеоценоза оказываются полуденные песчанки с их роющей деятельностью, приуроченной к контактным участкам края проекций крон и непокрытых древесно-кустарниковой растительностью полян с илаком - песчаной осокой вздутой, в меньшей степени - большие песчанки и затем - тонкопалые суслики (см. рис. 14). Важная роль принадлежит повреждающим древесину саксаула насекомым - саксауловому усачу (Turcmenigena varentzovi) и златкам (Julodis) в сочетании с древесным грибом - порией пустынной (Poria desertorum). Пория развивается в ходах личинок, особенно в зоне корневой шейки дерева. Под влиянием гриба древесина становится рыхлой и сыпучей, что усиливает ценотический эффект повреждающей деятельности личинок жуков и способствует более быстрому выпадению перестойных и даже спелых деревьев.
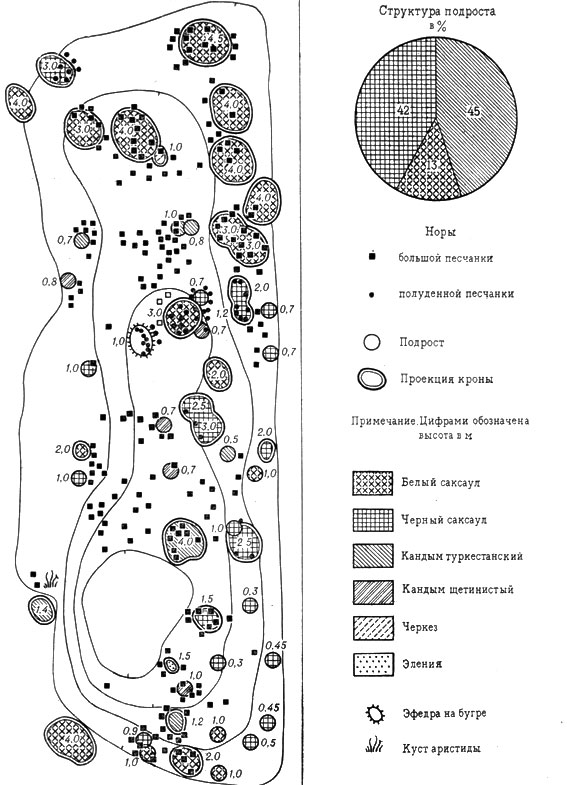
Рис. 14. Поселение большой песчанки котловинно-лощинного типа и колонии полуденной песчанки в кандымово-илаковом белосаксаульнике с участием расселяющегося черного саксаула (песчаная пустыня южного типа, Восточные Каракумы. Пробная площадь 0,2 га)
Основные характеристики биогеоценоза эфедрового черносаксаульника в долинных понижениях в условиях Репетекского заповедника: древостой - 490-500 деревьев черного саксаула на 1 га, спелые деревья - 50-56%, отдельные кусты и куртины эфедры - в среднем 70 на 1 га (при этом 22,2% эфедровых кустов растут не на буграх, 33,3% - образуют песчаные с погребенным опадом бугры до 1 м высотой и выше и 44% кустов встречаются в виде куртин на долголетних сложных буграх - эфедровых городках зоофитоэолового происхождения). Этот тип насаждения нередко бывает представлен вариантами с большей или меньшей разреженностью древостоя и кустарников (см. табл. 14).
| Вид дерева или кустарника и их возрастная группа | Количество | Распределение | |||||
| шт/га | % | Одиночное, шт/га | Групповое (по 2-5 шт.) | На эфедровых городках | |||
| Куртины, шт/га | Растения, шт/га | Число бугров, шт/га | Растения, шт/га | ||||
| Черный саксаул: | 480 | 52,4 | 96 | 44 | 204 | 16 | 180 |
| спелые и перестойные деревья (15-40 лет) | 212 | 23,2 | 60 | 44 | 132 | 8 | 20 |
| приспевающие (5-15 лет) | 112 | 12,1 | 28 | 12 | 24 | 12 | 60 |
| подрост (1-4 года) | 156 | 17,1 | 8 | 20 | 40 | 16 | 108 |
| пень на бугре | 3 | 0,4 | -* | - | - | - | - |
| Эфедра шишконосная: | 308 | 34,0 | 8 | 44** | 188 | 16 | 112 |
| взрослые кусты | 300 | 33,0 | - | 44 | 188 | 16 | 112 |
| подрост | 8 | 1,0 | 8 | - | - | - | - |
| Кандымы: | 100 | 11,0 | 100 | - | - | - | - |
| взрослые кусты | 32 | 3,7 | 32 | - | - | - | - |
| подрост | 68 | 7,3 | 68 | - | - | - | - |
| Черкезы: | 24 | 2,6 | 2,4 | - | - | - | - |
| взрослые кусты | 8 | 0,9 | 8 | - | - | - | - |
| подрост | 16 | 1,7 | 16 | - | - | - | - |
* (Знак указывает на отсутствие в составе насаждения данной категории растений.)
** (Куртины эфедры на буграх.)
В этом типе биогеоценоза со сложной структурой бывает до двадцати поселений на 1 га полуденных песчанок, включая как заселенные, так и оставленные зверьками. Большие песчанки образуют здесь в среднем до 7 небольших поселений, тонкопалый суслик - до 1,5-3 нор на 1 га (редко 5 нор). Наибольшая численность тонкопалого суслика наблюдается в разреженных белосаксаульниках. Поселения полуденных песчанок на эфедрогенных буграх и "городках" составляют 60% от их общего количества в эфедровом черносаксаульнике; поселения под саксаулом, преимущественно на периферийной части засоленной подкроновой салфетки и в зоне ее бордюрной растительности, - 26,7%, под кандымами - 13,3%.
В эфедровом черносаксаульнике можно выделить 6 парцелл:
- оголенные, лишенные растительности подкроновые пятна (салфетки);
- периферийные полосы их до 0,4-0,6 м шириной (где грызунами нарушается плотная засоленная поверхностная корка и развиваются ветроэрозионные процессы, что содействует частичному рассолению почвы);
- бордюрные кольца подкроновой растительности до 0,5-1,2 м шириной;
- эфедрогенные бугры с кустами эфедры шишконосной и ее подкроновыми микрогруппировками;
- подкроновая растительность кандымов и черкезов нередко на прикустовых холмиках песка, надутого ветром;
- поляны с осокой вздутой, создающей проективное обилие от 15 до 23% и закрепляющей песок своей дерниной.
В эфедровых черносаксаульниках с разной степенью разреженности древостоя, отличающихся количественными соотношениями видов кустарников, суммарная площадь парцелл и их пропорции изменяются в довольно широком диапазоне. В прямой зависимости от соотношения площадей парцелл наряду с влиянием других факторов находится в биогеоценозе общее количество нор песчанок обоих видов и соответственно выходов из нор. Именно число последних и учитывалось на единицу площади как показатель для определения размеров роющей деятельности грызунов.
При возрастании площади подкроновых салфеток (до определенного предела, соответствующего моменту смыкания крон) коррелятивно возрастает суммарная площадь бордюрных колец из растений-спутников черного саксаула. При этом темп роста площади бордюрных группировок опережает увеличение суммарной площади подкроновых пятен фитогенного засоления. Вместе с тем увеличивается и число нор песчанок, поскольку они локализуются преимущественно по периферии подкроновой салфетки и в краевой зоне бордюрных колец растительности. Для получения более четкого представления о преимущественной приуроченности нор песчанок к тому или иному типу парцелл можно использовать простое понятие "насыщенность парцеллы норами", которое представляет собой расчет количества нор, приходящегося на суммарный гектар каждой парцеллы (как если бы вся гектарная площадка была занята только одной парцеллой). Так, если норы полуденных песчанок в периферийной части подкроновой салфетки составляют 34% общего количества нор, то насыщенность норами этого типа парцеллы оказывается равной 57%.
Кривая насыщенности парцелл норами указывает на особое значение, которое имеют в жизни черносаксаулового биогеоценоза периферия подкронового пятна фитогенного засоления, кольца бордюрной растительности и эфедрогенные бугры (см. рис. 9). Именно в этих участках биогеоценоза получают наибольшее развитие ветроэрозионные процессы, непосредственно связанные с объемом роющей деятельности песчанок и обеспечивающие вторичное усиление ее биогеоценотического эффекта. Смена ряда стадий биогеоценотического цикла во многом зависит от развития ветровых процессов в этих парцеллах, где наблюдается активизация геоморфологических явлений, изменения солевого и водного режима почвогрунтов и идущие параллельно с ними закономерные сукцессии микрогруппировок подкроновой растительности.
Прежде чем рассмотреть последовательные стадии песчанково-черносаксаулового циклогенеза, необходимо коснуться узла биоценотических связей, анализ которого может объяснить причину того явления, когда черносаксауловые древостои даже в заповедных условиях в южных пустынях имеют предельный возраст 43-45 лет (в то время как в ботанических садах дерево черного саксаула способно жить более 100-150 лет).
Экологическое лимитирование возраста черносаксаульников в песчаных пустынях Средней Азии представляет собой результат взаимодействия нескольких факторов: деятельности личинок саксаулового усача, повреждающих древесину; температуры среды, определяющей характер локализации личинок в стволе и корне, и влияния на прочность древесины паразитического гриба - пории пустынной, поселяющегося в ходах личинок и разрушающего древесину изнутри.
Спелые деревья черного саксаула в возрасте 25- 30 лет и старше поражаются личинками усача и златок на 100%. Размеры повреждения древесины особенно возрастают у перестойных деревьев, достигших возраста 40-43 лет. В поперечном сечении корневой шейки к этому времени насчитывается до 30-46 ходов личинок. Если учесть, что ширина личиночного хода 4X8 мм, а сечение, следовательно, равно примерно 0,3 см2, то легко представить себе, насколько источенной оказывается древесина в участках концентрации личинок. На разломах корневая шейка перестойного саксаула напоминает соты с непрочными, крошащимися стенками (см рис. 15). Суммарная площадь поврежденной древесины на поперечном сечении корневой шейки дерева достигает 7,3-24,5 см иногда больше, что при ее размерах (по диаметру) 14X16 см и 23X24 см представляет очень существенное повреждение, снижающее сопротивление ствола на излом. Именно поэтому перестойные деревья с тяжелой кроной и обычно с наклоненным стволом при сильных ветрах не способны выдержать совмещенных сил ветра и тяжести и нередко обрушиваются, имея еще вполне жизнеспособный ассимиляционный аппарат непродуктивные органы и нормально функционирующую корневую систему. Это явление можно назвать скорее "энтомовалом", чем ветровалом. Для перестойных черносаксаульников очень типична картина упавших, развалившихся стволов с живой зеленой кроной.
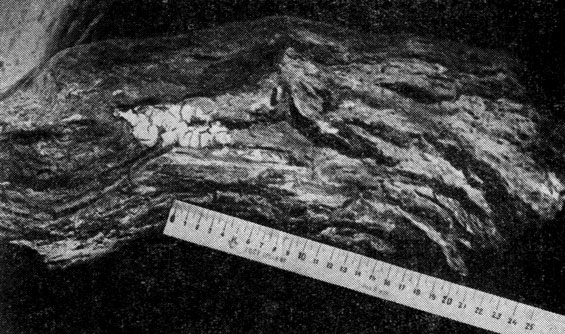
Рис. 15. Корневая шейка и верхний отрезок корня черного саксаула, поврежденные личинками саксаулового усача (видны крупные личиночные ходы и тело древесного гриба - пории пустынной, - развивающегося в ходах личинок и способствующего разрушению древесины, которая становится рыхлой и сыпучей)
Как показали наблюдения, личинки саксаулового усача и златок концентрируются в узком отрезке ствола и корня до 40-60 см длиной, примыкающем к корневой шейке. Общая длина ходов в стволовой части дерева и в корне колеблется от 24 до 63 см, обычно около 40 см (по 20-25 см в обе стороны от корневой шейки). При перенаселении комлевой части личинками длина ходов увеличивается. Причина указанного распределения личинок в древесине заключается в особенностях градиентов температуры разных точек ствола и кроны дерева.
Измерения температуры среды, проводившиеся в разных точках эфедро-саксаулового биогеоценоза, в разные сезоны, показали, что температура поверхности корневой шейки и почвы у комля черного саксаула оказывается относительно низкой и наиболее стабильной по сравнению с показателями температуры в кроне и тем более на поверхности почвы (см. рис. 16).
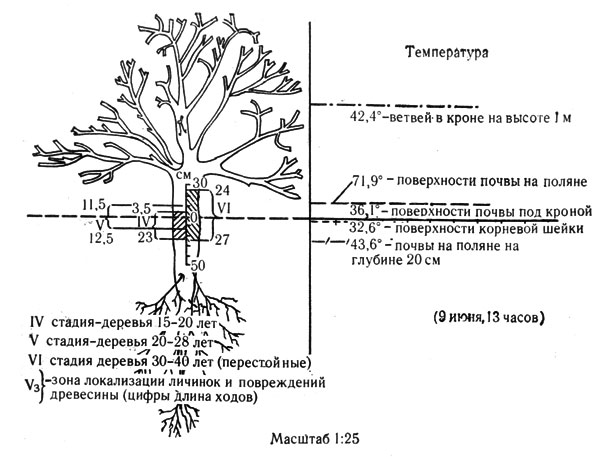
Рис. 16. Локализация личинок саксаулового усача в древесине ствола и корня черного саксаула в зависимости от температуры среды
В сезон высоких температур среды, с мая по сентябрь, комлевая часть дерева оказывается относительно менее нагреваемой точкой в биогеоценозе.
Разница температур в кроне и под комлем саксаула достигает 9-10°. Это обстоятельство вместе с отмеченной относительной стабильностью температур под комлем создает в корневой шейке деревьев и кустарников в пустыне более комфортные условия для беспозвоночных животных, чем и объясняется преимущественная локализация здесь личинок усачей и златок. Этот факт относится к явлениям того же порядка, что и "смена стаций" (Бей-Биенко, 1962), и "смена биоценотических горизонтов" (Гиляров, 1951).
Воздействие личинок на древесину усиливается в результате жизнедеятельности паразитического гриба - пории пустынной. Этот гриб проникает внутрь ствола вслед за личинками усача и разрастается затем по их ходам. Беловатое тело гриба и потемневшие, сыпучие и ломкие участки древесины вокруг него можно обыкновенно обнаружить на разломах комлевой части дерева. Химическое разрушение тканей древесины черного саксаула в результате деятельности пории, усиливающее ломкость ствола у корневой шейки, оказывается также одним из существенных биоценотических факторов, участвующих в комплексе, лимитирующем экологический возраст черносаксаульников в Средней Азии, на уровне 43-45 лет, всюду на тех территориях, где численность саксаулового усача обеспечивает стабильное поражение древостоев.
Биогеоценотический цикл черного саксаула и сукцессия его подкроновой растительности включает 12 последовательных стадий трансформаций песчанково-черносаксаулового ценотического узла, в процессе смены которых изменяется уровень фитогенного засоления подкроновой площадки, качественный состав, обилие и морфология ее травянисто-кустарничкового бордюра и доля участия зоофитоэоловых процессов. Цикл закономерно повторяется (табл. 15). Знание функциональной обусловленности биогеоценотической структуры, ее зонально типичной схемы важно при осуществлении "принципа биогеоценотической мелиорации" в практике освоения и облесения пустынных земель.
| Стадии циклогенеза | Возраст | Виды растений на подкроновом пространстве, преимущественно на "бордюрном кольце" |
| I стадия подроста черного саксаула | 1-4 года. Высота 0,4-1,2 м |
Carex physodes (проективное покрытие 12-15%, такое же, как на илаковых полянах) |
| II стадия молодого деревца. Появление под кроной пятен - скоплений опада. Проникновение на затененное подкроновое пространство злаков. Проективное покрытие подкроновой растительности увеличивается до 30-35% |
5-8 лет. Высота деревца 1,2-2 м |
Carex physodes (проективное покрытие увеличивается до 20%) Bromus (Anisantha) tectorum (проективное покрытие 10-20% или меньше) |
| III стадия "разнотравно-злакового бордюрного кольца". Начало фитогенного (содового) засоления подкронового пространства, постепенный "уход" травянистой растительности из-под кроны и образование "бордюрного кольца" по периферии проекции кроны из злаков, преимущественно костра и шизмуса арабского, и разнотравья. Появление отдельных нор полуденных песчанок на "бордюрном кольце растений" |
8-15 лет. Высота дерева 2-4 м |
Bromus tectorum* Schismus arabicus Eremopyrum orientate Senecio subdentatus Microcephala lamellata Ceraiocephalus falcatus Carex physodes |
| IV стадия "злаково-ромашникового бордюра оголенного подкронового пятна ("салфетки") и сформированной "зоны нор" полуденных песчанок". Фитогенное засоление под кроной вызывает отмирание травянистой и кустарничковой растительности. Образуется оголенная солевая корка ("содовый солончак") толщиной 8-12 мм. Микрогрупиировки подкроновой растительности, преимущественно шизмусово-ромашниковые и разнотравно-кохиевые концентрируются на периферии пятна засоления, где на границе двух разных сред образуют хорошо развитое кольцо растительности шириной 50-80 см, дающее втрое-вчетверо большую надземную фитомассу, чем песчаная осока с той же площади. На этой стадии формируется "зона нор" полуденных песчанок, по периферии пятна засоления и бордюрного кольца растений, стимулирующая начало ветроэрозионных процессов |
15-20 лет. Приспевающие деревья. Высота 4-6 м. Ширина кроны обычно равна ее высоте в редкостойном черносаксаульнике (от 230 до 360 деревьев на 1 га) |
Schismus arabicus Microcephla lamellata Eremopyrum orientate Senecio subdentatus Kochia schrenkiana Bromus tectorum Ceratocephalus falcatus Streptoloma desertorum Atriplex dimorphostegia Silene nana |
| V стадия "злаково-солянкового (шизмусово-лондезиевого) бордюрного кольца и развития активных зооэоловых процессов в краевой зоне нор". В зоне нор, располагающейся по обе стороны бордюрного кольца растений, под воздействием роющей деятельности полуденных песчанок нарушается целостность поверхности почвогрунта, разрыхляется плотная корка засоления и активизируются ветроэрозионные процессы. Наиболее сильный вынос рыхлого материала происходит в краевой полосе по внешней стороне бордюрного кольца растений, где формируется желоб выдувания, соответствующий направлению ветровых потоков, огибающих крону и травянисто-кустарничковое кольцо. В составе "бордюра" начинают преобладать солянки. Засоление подкронового пространства усиливается за счет вымывания солей из опада и смыва их с кроны |
20-28 (30) лет. Спелые деревья. Высота 6-8 (9) м. Начало частичного усыхания кроны |
Schismus arabicus Londesia eriantha Kochia schrenkiana Suaeda lipskyi Eremopyrum orientate Eremopyrum buonapartis Ceratocephalus falcatus Bromus tectorum Atriplex dimorphostegia Bromus danthoniae |
| VI стадия "дерева на столообразном бугре и двухъярусного кольца бордюрной растительности". Зона нор расширяется в сторону ствола. На осветленных участках под усохшей кроной поселяются большие песчанки (смена преобладающего вида - "биорегулятора") | 28-40 лет. Перестойные деревья. Частичное отмирание кроны | Верхний, внутренний ярус кольца (зона нор): Londesia eriatha Kochia schrenkiana Schismus atabicus Suaeda lipskyi Atriplex dimorphostegia Streptoloma desertorum Ceratocephalus falcatus Eremopyrum buonapartis Brotnus tectorum Microcephala lamellata Senecio subdentatus Silene nana В теневом пятне с северной стороны дерева (зона нор): Bromus tectorum Schismus arabicus Eremupyrum orientate Eremopyrum buonapartis Kochia schrenkiana Londesia eriantha Malcolmia grandiflora Microcephala lamellata Ceratocephalus falcatus Senecio subdentatus Alyssum desertorum Schismus culycinus Papaver pavoninum В нижнем ярусе кольца: Bromus tectorum Schismus arabicus Eremopyrum orientate Senecio subdentatus По внешнему краю желоба выдувания: Horaninowia minor Nonea caspica Arnebia decumbens Bromus tectorum Carex physodes. |
| VII стадия останцового бугра с пнем и "энтомовалом". Эрозионное рассоление полкронового пятна и асимметричное сокращение его площади. Миграция однолетних солянок и злаков в направлении к пню | 40-43-50 лет | Бордюрная растительность верхнего яруса кольца и околопневой поросли (зона нор): Kochia schrenkiana Londesia eriantha Schismus arabicus Suaeda lipskyi Schismus culycinus Eremopyrum orientate Eremopyrum buonapartis. Бордюрная растительность нижнего яруса кольца у желоба выдувания: Schismus arabicus Eremopyrum orientate. Бордюрная растительность желоба выдувания (внешний край): Horaninowia minor Astragalus chiuensis Heliotropium dasycarpum Carex physodes Aristida pennata. |
| VIII стадия сокращающегося останцового бугра с широким выполаживающимся желобом выдувания, который заполняется продуктами активной дефляции с бугра. Нарушение целостности коры фитогенного засоления, делящейся на фрагменты норами преимущественно большой и отчасти полуденной песчанок | 50-70 лет | Бордюрное кольцо растений на останцовом бугре нарушено, имеет вид группы пятен из микрогруппировок однолетних солянок и злаков, обычно у пня: Londesia erianiha Kochia schrenkiana Sueda lipskyi Schismus arabipus Bromus tectorum Eremopymm orientate Растения в желобе выдувания и по его внешнему краю: Horaninowia ulicina, H. minor Carex physodes (вторичное наступление) Salsola sp. Haloxylon aphyllum (подрост в зоне нор и на делювии) |
| IX и X стадии выполаживания остаточного бугра за счет заполнения желоба выдувания разрушенным материалом и закрепления дерниной наступающей осоки. Активное рассоление. Корки "содового солончака" почти не остается. Сокращение числа обитаемых нор большой и особенно полуденной песчанок | 70-90 лет | Остаточное пятно однолетне-солянковой растительности на месте бывшего пня и рядом: Londesia eriantha Kochia schrenkiana Schismus arabicus Schismus culycinus. На делювии в остаточном "желобе заполнения": Horaninowia ulicina Aristida pennata Carex physodes |
| XI стадия образования воронки выдувания (отрицательная форма рельефа, происходит вынос песка) на месте останцового бугра черного саксаула, представлявшего собой стабилизированный исходный уровень поверхности песка. По краю воронки встречаются норы большой песчанки (маленькие колонии) или отдельные норы тонкопалого суслика | 90-110-120 лет | По краю воронки выдувания: Aristida pennata Horaninovia ulicina Heliotropium arguzioides Astragalus chivensis Подрост: Calligonum setosum C. turcestanicum На месте разрушившегося пня: Carex physodes Schismus arabicus (единично) и подрост Haloxylon aphyllum У нор встречаются: Bromus tectorum Senecio subdentatus |
| XII стадия выравнивания воронки выдувания и зарастания ее песчаной осокой; ограничивается вынос песка и начинает преобладать процесс накопления подвижных частиц грунта | 120-150 (?) лет | В растительном покрове доминирует на незакрепленных участках песка: Carex physodes Heliotropium argwtioides Astragalus chivensis В зоне единичных нор землероев, преимущественно больших песчанок, обычен (до 30-50%) подрост черного саксаула |
* (Последовательность видов в списках отражает их соотносительное преобладание на каждой стадии)
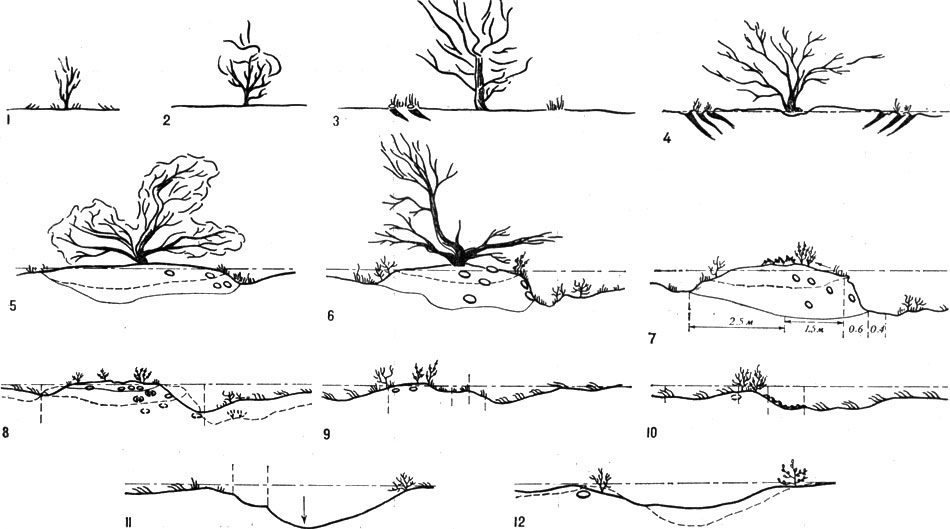
Рис. 17. Стадии биогеоценотического циклогенеза черного саксаула: 1 - первая; 2 - вторая; 3 - третья; 4 - четвертая; 5 - пятая; 6 - шестая; 7 - седьмая; 8 - восьмая; 9 - девятая; 10 - десятая; 11 - одиннадцатая; 12 - двенадцатая
Онтогенетический цикл черного саксаула - вида, в сфере жизнедеятельности которого оказываются многие виды растений и животных, описывается как пример биогеоценотического циклогенеза, включающего взаимодействия различных организмов и закономерные сукцессии подкроновой растительности на разных стадиях цикла (см. рис. 17).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'