Глава 2. Рыбы приповерхностных вод
Должен сознаться, что, помимо объективного профессионального интереса, я испытываю к рыбам и вполне определенные субъективные чувства: одни из них нравятся мне, другие более или менее безразличны, третьи (совсем немногие), вовсе не вызывают симпатий. Особое место в этом ряду принадлежит той группе, которая была первым объектом моего изучения и одновременно восхищения...
Представьте себя на палубе теплохода, курс которого проложен в тропической зоне открытого океана... Кругом бесконечное синее небо с белоснежными хлопьями редких кучевых облаков и такая же бесконечная, но еще более синяя гладь воды, ослепительно сияющая под ярким солнцем... Ничто не нарушает однообразия приятного глазу, но безжизненного ландшафта. Вдруг совсем близко, всего в нескольких метрах от борта, выскакивают на поверхность воды и, недолго проскользив по ней, стайкой взмывают в воздух, сразу разлетаясь широким веером, небольшие крылатые существа, кажущиеся так похожими на привычных птиц. Это — летучие рыбы...
Когда, стоя на мерно раскачивающемся полубаке, вглядываешься прямо вниз — в разрезаемую форштевнем тихо поющую воду, часто удается увидеть летучек еще до взлета. Застигнутые врасплох, они резко срываются с места, и почти мгновенно темно-синие силуэты миниатюрных подлодок, будто по мановению волшебной палочки, превращаются в игрушечные ширококрылые планеры (вот он — двухсредный транспортный аппарат, мечта конструкторской мысли!). Правда, в этой экстремальной ситуации летучие рыбы редко летят долго. Как правило, они вновь падают в воду через несколько метров, чтобы опять взлететь, теперь уже разогнавшись по всем правилам искусства.
Летучие рыбы (рис. 4) — представители очень характерного для верхней эпипелагиали тропиков семейства Exocoetidae, которое насчитывает в своем составе более 60 видов. Они не достигают особенно больших размеров: самый крупный вид — гигантская летучка (Cheilopogon pinnatibarbatus), обитающая на периферии тропической зоны, может иметь длину около 50 см при массе более 1 кг, а самые мелкие не превышают 15 см и весят всего 30— 50 г. Наиболее характерная особенность летучих рыб — огромные грудные плавники, соизмеримые с длиной туловища. При типичной для обитателей приповерхностного слоя окраске туловища (темно-синяя спина, серебристые бока и брюхо) грудные плавники имеют довольно разнообразный цвет: они могут быть как однотонными — прозрачными, синими, зелеными, коричневыми, так и пестрыми — с яркими полосами или пятнами.

Рис. 4. Сарганообразные рыбы открытого океана: двухкрылая (1) и четырехкрылая (2) летучие рыбы, макрелещука (3).
Уникальной особенностью летучих рыб является их способность к полету, развившаяся, очевидно, в качестве приспособления для спасения от хищников. Эта способность выражена в разных родах в неодинаковой степени. Полет примитивных видов летучих рыб, которые обладают сравнительно короткими грудными плавниками (представители родов Fodiator и Parexocoetus), менее совершенен, чем у видов с длинными крыльями. При этом эволюция полета в пределах семейства происходила, очевидно, в двух направлениях. Одно из них привело к образованию «двукрылых» летучих рыб, сравниваемых иногда с самолетами-монопланами и использующих при полете только грудные плавники, которые достигают у них очень больших размеров (типичный представитель — обыкновенный двукрыл Exocoetus volitans). Другое направление представлено четырьмя родами «четырехкрылых» летучих рыб (около 50 видов), которые уподобляются самолетам-бипланам. Полет этих рыб осуществляется при помощи двух пар несущих плоскостей, так как у них увеличены не только грудные, но и брюшные плавники, причем на мальковых стадиях развития те и другие имеют приблизительно одинаковую площадь. Оба направления эволюции полета привели к образованию форм, хорошо приспособленных к жизни в поверхностных слоях океана. Кроме развития «крыльев», приспособление к полету отразилось у летучих рыб в строении хвостового плавника, лучи которого жестко соединены между собой, а нижняя лопасть очень велика по сравнению с верхней, в необычном развитии огромного плавательного пузыря, продолжающегося под позвоночником до самого хвоста, и в других особенностях.
Наибольшей дальности и продолжительности достигает полет «четырехкрылых» летучих рыб. Развив в воде значительную скорость (порядка 60—65 км/ч), такая рыба выскакивает на поверхность моря и некоторое время (иногда совсем недолго) глиссирует по ней с расправленными грудными плавниками, энергично увеличивая скорость при помощи колебательных движений погруженной в воду длинной нижней лопасти хвостового плавника. Затем рыба отрывается от воды и, раскрыв брюшные плавники, планирует над ее поверхностью. В некоторых случаях летучка при полете слегка касается воды нижней лопастью хвостового плавника и, вибрируя им, получает дополнительное ускорение. Количество таких касаний может достигать трех-четырех, и в этом случае продолжительность полета, естественно, возрастает. Обычно летучая рыба находится в полете не более 10 с и пролетает за это время несколько десятков метров, но иногда длительность полета увеличивается до 30 с, а дальность его доходит до 200 и даже до 400 м. При наличии слабого ветра или восходящих токов воздуха над водой летучие рыбы пролетают большие расстояния и дольше находятся в полете. Высота полета обычно не превышает 1—2 м, но по утрам летучек нередко находят на открытых палубах судов, в том числе крупнотоннажных, в удалении на 10—12 м от воды. Рыбы залетают на палубу только на ходу судна и всегда с наветренной стороны: по-видимому, они привлекаются судовыми огнями и попадают у борта в восходящий, поток воздуха, высоко поднимающий их вверх.
Многие моряки и путешественники, наблюдавшие летучих рыб с палубы корабля, утверждали, что они ясно видели, что рыба машет крыльями точно так же, как стрекоза или колибри. В действительности «крылья» летучек при полете совершенно неподвижны и не совершают никаких взмахов или колебаний. Лишь угол наклона плавников может, по-видимому, меняться, и это позволяет рыбе несколько отклоняться от прямого направления. То дрожание плавников, которое отмечают очевидцы, представляет собой не причину полета, а его следствие. Оно объясняется непроизвольной вибрацией расправленных «крыльев», особенно сильной в те мгновения, когда рыба, уже находящаяся в воздухе, еще продолжает работать в воде хвостовым плавником.
Летучие рыбы обычно держатся небольшими стайками, как правило, до десятка особей. Эти стайки состоят из близких по размерам рыб, принадлежащих к одному виду. Отдельные стайки часто группируются в крупные косяки, а в наиболее кормных районах образуются иногда значительные скопления летучих рыб, состоящие из многих косяков.
Видовой состав летучих рыб заметно различается в прибрежных и удаленных от берегов районах. Одни виды встречаются только в непосредственной близости от побережий, другие могут выходить и в открытый океан, но для размножения возвращаются в прибрежную зону, третьи (их явное меньшинство) постоянно населяют океанические просторы. Основная причина такого разделения — различные требования к условиям нереста. Почти все летучие рыбы имеют икру, снабженную клейкими нитевидными придатками, которые обычно служат для ее прикрепления к донным или плавающим водорослям-макрофитам. Такого субстрата в открытом океане немного, так как участки, богатые плавучими водорослями саргассами, занимают в общем незначительные площади. Поэтому океанические летучие рыбы используют в качестве нерестового субстрата то небольшое количество плавающего материала, которое всегда имеется в море — в основном различный плавник берегового происхождения (дрейфующие водоросли, ветви и плоды наземных растений, кокосовые орехи), птичьи перья и даже беспозвоночных животных, например хондрофор, живущих на поверхности воды. Только «двукрылые» летучки (3 вида рода Ехоcoetus) имеют плавучую икру, утратившую клейкие нитевидные выросты, и именно поэтому они резко преобладают вдали от берегов, где на их долю приходится в разных районах от 70 до 90% всех летучих рыб.
Летучие рыбы питаются планктонными животными, обитающими в поверхностном слое, в основном мелкими ракообразными и крылоногими моллюсками, а также личинками рыб. Их основными пищевыми конкурентами в тропической зоне являются представители семейства светящихся анчоусов (Myctophidae) — группы, в целом характерной для глубоководной пелагиали (см. гл. III), но включающей также виды, которые поднимаются в темное время суток к самой поверхности ( К числу постоянных обитателей океанской эпипелагиали принадлежит и один вид настоящих анчоусов (семейство Engraulidae) — Stolephorus buccaneer, довольно обычный в открытых водах западной тропической части Тихого океана.). Приповерхностные, или никтоэпипелагические (от греческого слова «никтиос» — ночь), миктофиды, принадлежащие к родам Myctophum, Symbolophorus и др., достигают высокой численности. Размах суточных вертикальных миграций этих небольших рыбок (обычная длина 6—10 см) достигает 500—900 м: за какой-нибудь час они преодолевают, таким образом, расстояния, в несколько тысяч раз превышающие длину их тела. Никтоэпипелагические светящиеся анчоусы достаточно многочисленны и в умеренных водах — там, где летучие рыбы, область распространения которых ограничена, грубо говоря, изотермой 20° на поверхности, не живут. В умеренно тепловодной зоне они уступают; роль массовых потребителей зоопланктона родственной группе — семейству макрелещуковых (Scomberesocidae).
Все макрелещуковые не связаны с прибрежными водами ни в один из периодов жизненного цикла и являются характерными представителями собственно океанической группировки рыб. В состав этого семейства входят всего 4 вида. Область распространения макрелещуки (Scomberesox saurus) включает северную часть Атлантического океана и Средиземное море, причем в годы потепления Арктики отдельные экземпляры ловились даже в Баренцевом море у Новой Земли. Макрелещука обитает также в южном полушарии, где она встречается во всех океанах в широком поясе между 15—30° и 40—50° ю. ш. Сайра (Cololabis saira) населяет воды северной части Тихого океана (между 20 и 55° с. ш.) и Японского моря. Максимальная длина макрелещуки не превышает 45—50 см, сайры — 36—40 см. Заслуживает упоминания и еще один вид семейства — восточнотихоокеанская карликовая сайра (Elassichthys adocetus), которая при максимальной длине порядка 5—6 см является самой мелкой из настоящих эпиелагических рыб.
Макрелещуковые держатся в верхних слоях океана, обычно у самой поверхности воды, и, будучи возбужденными или напуганными, часто совершают прыжки над водой. Они редко выходят за пределы верхнего 10-метрового слоя, хотя сайра в период зимовки погружается на глубину около 50 м. Макрелещука и сайра — стайные рыбы, образующие в определенные периоды жизни значительные скопления. Они размножаются в открытом океане. При этом сайра откладывает икру, снабженную пучком клейких нитевидных придатков, на твердый плавучий субстрат — обычно на саргассовые водоросли, но, вообще говоря, на любые предметы, находящиеся в воде, в том числе на рыболовные сети и яруса для лова тунцов. Макрелещука, напротив, имеет пелагические икринки, развивающиеся в планктоне. Оба вида размножаются в наиболее прогретых частях своих ареалов, расположенных в субтропической зоне. Холодноводная часть области распространения используется только для летнего нагула.
Для макрелещуковых рыб очень характерны сезонные миграции, в ходе которых они перемещаются на значительные расстояния. Например, в северо-западной части Тихого океана в водах теплого течения Куросио сайра проводит зиму. Здесь при температуре 14—25° происходит ее нерест, во время которого рыба держится разреженными стайками. С наступлением весны начинается перемещение отнерестившихся особей к северу, и в начале лета (июнь—июль) сайра входит в воду холодного течения Оясио для нагула. В теплое время она появляется у Курильских островов и даже у берегов Восточной Камчатки, а иногда и в Беринговом море, где может достигать Олю-торского залива. В северных водах сайра откармливается в течение всего теплого времени года при температуре от 6 до 18°. С началом охлаждения вод и окончанием нагула стайки сайры вновь смещаются к югу. В этот период (сентябрь—ноябрь) образуются значительные скопления этой рыбы в районе стыка Оясио и Куросио, а к началу зимы вся сайра оказывается в теплых водах у южных берегов Японии.

Рис. 5. Океанические акулы: гигантская (1), кархародон (3), синяя (3)
Летучие рыбы, сайра и макрелещука, а также приповерхностные светящиеся анчоусы принадлежат к числу рыб, привлекаемых источниками искусственного освещения: реагируя ночью на судовые огни, летучие рыбы, например, нередко залетают на палубу судов, а в отдельных случаях даже в открытые иллюминаторы кают. Такая особенность поведения важна как для научного изучения этих рыб, так и для их промысла. Наблюдения на световых станциях, часто включаемых в программы экспедиционных рейсов, позволяют узнать много интересного.
Тихая темная ночь... Судно медленно дрейфует, развернувшись лагом к чуть заметному ветерку... В таинственно мерцающей голубой линзе воды, границы которой постепенно переходят в кромешную черноту, появляются то зигзагом проскакивающие освещенную зону сайры, сверкающие, как рыболовные блесны, то менее скоростные, но очень юркие светящиеся анчоусы, то медленно дрейфующие по поверхности с расправленными крыльями мальки летучих рыб... Взрослые летучки подплывают к свету поодиночке или стайками, а иногда и подлетают над водой, с силой ударяясь головой о борт судна... Появление кальмаров и хищных рыб — змеиных макрелей и корифен — всегда неожиданно: их стремительные броски в световом пятне, нередко заканчивающиеся захватом добычи, напоминают выпады разящего клинка или сверкание молнии... Крупные акулы (у поверхности наиболее обычна длиннокрылая), напротив, пересекают зону света медленно и чинно...
Иной раз, впрочем, приходится часами простаивать, облокотившись о планширь и ожидая появления очередной рыбы с готовым к броску закидным сачком — сеткой на обруче, к которому прикреплен длинный линь (сказать бы попросту «веревка», но этого слова в морском лексиконе нет). Ловля таким сачком — настоящее искусство, в котором истинным асом был на «Витязе» Григорий Касьянович Фисунов - старый моряк, долго занимавший пост техника-лаборанта при ихтиологической лаборатории. Ему удавались фантастические по дальности и точности броски... Правда, я тоже имел шанс отличиться. Однажды (было это в Панамском заливе), когда я глухой ночью «сачковал» в полном одиночестве на корме «Курчатова», в поле света вдруг появилось толстое, слегка заостренное бревно, которое неожиданно преобразилось в редкостного по величине саргана. И я поймал это чудище и тащил его, напрягая все силы (по рыбацкой оценке, пуда два он весил, если не больше), и почти вытащил... Вдруг гигант, до того смирно лежавший головой в сачке - большая часть его тела свисала через обруч,— сделал какое-то ленивое движение и сразу исчез, как будто и не существовал вовсе. О материальности происшедшего свидетельствовала только огромная дыра в сетке.
На световых станциях можно увидеть почти всех обитателей верхнего 20-метрового слоя эпипелагиали. Действительно, список океанических рыб, показывающихся более или менее регулярно у самой поверхности, можно пополнить совсем немногими видами. В их числе нельзя не упомянуть некоторых акул, рыб-лун, копьерылов и тунцов.
Китовая (Rhiniodon typus) и гигантская (Cetorhinus maximus) акулы (рис. 5), наибольшая официально зарегистрированная длина которых составляет соответственно 15,2 м (масса до 14 т) и 13,6 м (по неподтвержденным полевым данным, еще больше), занимают первое и второе места в ряду наиболее крупных из ныне живущих низших позвоночных. Оба вида в отличие от преобладающего большинства других акул питаются планктонными животными и мелкой рыбешкой и для удовлетворения своих энергетических потребностей нуждаются в высокой концентрации корма. Поэтому эти безвредные для человека акулы держатся в наиболее продуктивных районах океана (китовая — в богатых пищей участках тропической зоны, гигантская — в умеренно теплых водах обоих полушарий) , где нередко образуют стайные скопления. Обе они очень схожи по способу захвата пищи.
Китовая акула, неторопливо двигаясь (иногда — в вертикальном положении) у поверхности, всасывает съедобную живность, от фитопланктона и зоопланктона до стайных рыб (сардины, анчоусы, скумбрии, мелкие тунцы) и кальмаров. Затем она профильтровывает воду сквозь жаберные отверстия через своеобразное мелкое сито, образованное мягкой губчатой тканью, которая поддерживается хрящиками, соединяющими жаберные дуги. Затем отцеженные кормовые организмы через узкий пищевод попадают в желудок. В связи с таким способом питания зубы у этой акулы очень мелкие и многочисленные, они служат не для кусания, а только для «запирания» рта. Китовая акула, не имеющая естественных врагов, мало беспокоится о своей безопасности. Известны случаи, когда это медлительное животное, часто неподвижно покоящееся у поверхности воды, попадало под удар проходящих кораблей. Один из таких инцидентов произошел в 1905 г. Пассажирское судно, следовавшее в Индию, протаранило китовую акулу длиной, по оценке очевидцев, около 17 м и в течение 15 мин тащило ее перед собой на форштевне. Как сообщали газеты того времени, пассажиры, малознакомые с систематической зоологией, решили присвоить чудовищу «научное» наименование — Piscis Rudyardensis («Рэдьярдова рыба») в честь присутствовавшего на борту известного английского писателя Рэдьярда Киплинга.
Питающаяся на скоплении планктона гигантская акула плывет так же медленно (со скоростью около 3,5 км/ч), широко разинув пасть, и пропускает через ротовую полость воду, отфильтровывая планктон на весьма совершенном цедильном аппарате: каждая огромная жаберная дуга несет по переднему краю более 1000 длинных роговых жаберных тычинок. Ежечасно она процеживает около 1500 м3 воды. Желудок у гигантской акулы очень большой — у крупных питающихся особей в нем находили около тонны красноватой густой пасты, состоящей из планктонных рачков, перемешанных со слизью, которой они обволакиваются в полости рта. Таким образом, по способу питания гигантская акула мало отличается от усатых китов и, кстати, вместе с ними сильно пострадала от гарпунного промысла. Гигантские акулы (в прибрежных водах они более обычны, чем в открытом океане) встречаются у поверхности моря только весной и летом, когда вода наиболее богата планктонной пищей. В это время в наиболее кормных районах можно видеть целые стаи этих малоподвижных животных, насчитывающие до 20— 30 особей и медленно передвигающихся в поверхностном слое (английское название этого вида «basking shark» переводится как «акула, греющаяся на солнце»). Зимой гигантские акулы очень редко попадаются на глаза. Расчеты показывают, что только для возмещения энергетических затрат при плавании во время кормежки акула должна получать не менее 157,5 кДж/ч. Эта цифра намного превышает доступное поступление, так как количество планктона в умеренных широтах значительно понижается в зимнее время. Поэтому предполагают, что с наступлением зимы гигантская акула уходит из поверхностных вод, теряет жаберные тычинки, которые вновь отрастают только к следующей весне, и переходит в малоактивное состояние, соответствующее, по-видимому, зимней спячке млекопитающих. Подтверждением этому служит тот факт, что добытые ранней весной особи почти не содержат печеночного жира, используемого в период голодания.
В старые времена случайные встречи моряков с китовыми и гигантскими акулами, плавающими у поверхности с выступающими над водой спинным и хвостовым плавниками, немало способствовали широкому распространению различных россказней о морском змее и других невероятных чудовищах.
Среди акул, ведущих образ жизни активного хищника в открытом океане явно доминирует синяя акула (Prionace glauca) — возможно, наиболее процветающий вид современных хрящевых рыб. Она встречается во всех океанах и, по данным телеметрического слежения, придерживается температуры 12—16°, Поэтому в умеренно теплых и субтропических водах этот вид держится в поверхностных слоях океана, а в сильно прогретых сверху водах тропической зоны обычен на глубине порядка 100—150 м.
Наибольшая длина синей акулы, по-видимому, не превышает 3,8 м (при массе более 200 кг), хотя имеются недостоверные сообщения и о поимке более крупных особей. Питается она разнообразными рыбами и головоногими моллюсками, а также любой другой добычей, которую может отыскать и поймать, и иногда в течение долгого времени сопровождает медленно плывущие суда, поедая корабельные отбросы. Синяя акула живородяща и может приносить до 135 детенышей длиной около 50 см. Во время одной из экспедиций на «Витязе» половозрелая самка этого вида была поймана у юго-западной Австралии. Поднятая на судно акула разродилась на палубе 52 детенышами, совершенно подготовленными к самостоятельной жизни. Несколько новорожденных акулят, помещенных в большой аквариум, прекрасно себя чувствовали в течение нескольних суток. Районы размножения этой акулы находятся за пределами тропической зоны и приурочены к продуктивным прибрежным участкам, причем, как показано Ф. Ф. Литвиновым, северные и южные группы популяций вида имеют различия в форме зубов.
Наиболее обычная в поверхностных водах тропической и субтропической зоны (предпочитаемая температура 18—28°) длиннокрылая акула (Carcharhinus longimanus), названная так из-за очень больших грудных плавников, широко распространена во всех океанах и постоянно держится у поверхности (рис. 6). Он достигает 3,5—4 м в длину, но обычно встречаются некрупные особи длиной до 1,5—2 м и массой 20—60 кг. Как и синяя акула, она принадлежит к числу живородящих, но приносит в зависимости от размера всего 1 — 15 детенышей длиной 60—65 см. Питается длиннокрылая акула самыми различными объектами — рыбами, кальмарами, морскими птицами, любыми беспозвоночными — в общем всем, что она может отыскать в воде. Будучи очень медлительной и не имея возможности активно ловить быстроплавающих животных, она находит к ним другие подходы: наблюдали, в частности, как акула, плававшая с раскрытым ртом в скоплении мелких тунцов, питавшихся сардиной, терпеливо ожидала добычи, и действительно один из тунцов сам натолкнулся на нее. Приближающиеся к судам в открытых водах акулы, как правило, имеют в желудках только камбузные отходы. Следовательно, они могут следовать за кораблями, подбирая съедобные куски, выкинутые за борт. Эти акулы очень живучи. Говорят, будто бы пойманная и выпотрошенная акула, вновь брошенная в воду, продолжает как ни в чем ни бывало плавать около судна и может схватить наживленный крючок.

Рис. 6. Длиннокрылая акула о рыбами-лоцманами и рыбой-прилипалой
В тропических водах встреча с длиннокрылой акулой - самое заурядное событие. Одна, а то и две-три хищницы, почти всегда сопровождаемые рыбами-лоцманами, в любой момент могут появиться у борта во время океанологической станции и долго крейсировать поблизости, ожидая поживы. Акула в движении - удивительное и ни с чем не сравнимое зрелище. Она скользит в прозрачной воде без видимого усилия и без суетливости, почти расслабленно, по в то же время демонстративно мощно и с явным сознанием собственного достоинства и силы. Чувствуется, что это истинная владычица морская... Правда, лишь в отсутствие так называемого «царя природы» — был бы у «любителя» прочный крюк, надежный шнур и любое подобие наживки, и вожделенный трофей (конечно, бесполезный — ведь у нас не привыкли к акульему мясу), не заставит себя долго ждать.
Итак, обычно длиннокрылые акулы рыщут по морю поодиночке. Но ие всегда — мне дважды приходилось видеть большие их скопления: в 1958 г., когда «Витязь» проводил работы недалеко от островов Тонга, и в 1961 г. на одной из станций в Бенгальском заливе. В обоих случаях акулы не проявляли стайного поведения, каждая вроде бы «гуляла сама по себе» и не обращала внимания на сородичей, по в пределах видимости постоянно находились десятки экземпляров, и все довольно крупные. Оба раза акулы пе проявляли жадности к приманкам и брали крючок без особой охоты. Поэтому они понесли от «любительского рыболовства» довольно умеренные потери. В Бенгальском заливе контакт с длиннокрылыми акулами был более тесным, так как там мы спускали иа воду шлюпку, и этот редут они атаковали вал за валом, высовывая из воды головы и хватая зубами весла, а также багор и румпель, которыми мы пытались их отогнать. Вся эта картина напоминала описанную Германом Мелвиллом сцену последней погони капитана Ахава за Моби Диком, когда бесчисленные акулы крошили зубами лопасти весел его вельбота. Причина, которая заставила акул собраться в одном месте, остается мне непонятной...
Для эпипелагиали очень характерны и три рода ламновых акул (семейство Lamnidae) — Lamna, Isurus и Carcharodon, которые принадлежат к числу наиболее быстрых и подвижных хрящевых рыб. У всех этих акул, как и у тунцов, температура тела выше, чем температура окружающей воды (на 5—6°), что обеспечивает им более высокий уровень энергетического обмена по сравнению с преобладающим большинством рыб.
Акулы рода Lamna (их два вида, и оба встречаются в советских водах), достигающие 3—3,5 м в длину, живут в умеренно холодных водах обоих полушарий и питаются преимущественно стайными рыбами — тихоокеанскими лососями (лососевая акула L. ditropis) и сельдью (сельдевая акула L. nasus). Это быстрые пловцы, нередко образующие стаи. Они достигают половой зрелости в возрасте 5 лет и более и могут жить до 20—30 лет.
Их ближайшие родичи — серо-голубые акулы, или мако (два вида рода Isurus). Короткоперый мако (I. oxyrhinchus) — обитатель тропических вод, самая скоростная из всех ныне живущих хрящевых рыб (известный специалист по акулам Л. Компаньо называет ее «кречетом акульего мира»). Мако несколько крупнее сельдевых (3,5—4 м при массе до 450 кг) и нападает на очень крупную добычу — других акул, костистых рыб, кальмаров. Так, в желудках двух особей массой 300 и 360 кг были обнаружены съеденные меч-рыбы массой 54 и 67 кг. По-видимому, между акулами-мако и меч-рыбами могут разыгрываться настоящие сражения, о чем свидетельствует, в частности, обнаруженный в Аденском заливе труп выброшенной на берег серо-голубой акулы с обломками рострума меч-рыбы длиной 45 см, пробившего ее тело позади жаберных щелей.
Лисьи акулы (род Alopias с 3 видами) еще крупнее (до 6 м), но почти половина их длины приходится на сильно удлиненную верхнюю лопасть хвоста, которую они используют как основное орудие охоты: они сбивают рыб и кальмаров в плотную стайку и глушат их хвостовым плавником, действуя им как цепом. Чисто океанический вид — большеглазая акула-лисица (A. superciliosus) длиной до 4,5 м — обитает в нижней эпипелагиали и верхней мезопелагиали — на глубине до 500 м. Она питается алепизаврами, мелкими тунцами и меч-рыбой, кальмарами.
Все ламноидные акулы размножаются путем яйцежи-ворождения (оплодотворенное яйцо развивается в яйцеводе вплоть до рождения молоди) (Щележаберные рыбы (акулы и скаты) размножаются тремя способами: 1) откладывая на дно яйца в роговых капсулах (яйцерождение), как это делают, в частности, рогатые акулы и ромбовые скаты; 2) путем яйцеживорождения и 3) путем живорождения - в этом случае в заднем отделе яйцеводов («матка») имеется некое подобие детского места (плаценты), служащее для питания зародыша материнской кровью. Последний способ характерен для всех кархариноидных акул (кроме тигровой).) и характеризуются низкой плодовитостью — обычно приносят 2—5 крупных детенышей, редко больше. Особенностью этих акул является так называемая внутриутробная оофагия — поедание эмбрионами неоплодотворенных и развивающихся яиц непосредственно в яйцеводах матери (писатель И. И. Акимушкин назвал это явление «эмбриональным каинизмом»). Внутриутробное питание начинается довольно рано — уже при длине 4—5 см. Так, в желудках эмбрионов песчаной акулы (Eugomphodus taurus), которые при длине 10 см по развитию зубного аппарата почти не отличаются от взрослых рыб, находили большое количество яичного желтка и меньших по размерам (длиной до 4 см) зародышей. Наблюдения над живыми эмбрионами длиной около 26 см показали, что в последние 2—3 месяца перед рождением (общая продолжительность беременности составляет 9—12 месяцев) единственный остающийся к тому времени в яичнике эмбрион активно перемещается в овариальной жидкости среди выеденных им яйцевых капсул, периодически получая питание за счет новых порций неоплодотворенных яиц, поступающих по мере созревания.
Говоря о крупных акулах, нельзя не сказать об опасности, которую они представляют для человека. Как показывает накопленная статистика, большая часть зарегистрированных нападений по вполне понятным причинам приходится на теплые прибрежные воды: именно там собирается больше всего купающихся, ныряльщиков, подводных охотников. В открытом океане купание не получило распространения (более того, оно находится под строгим запретом почти на всех судах), и объектом нападения акул могут служить главным образом люди, оказавшиеся в воде в результате авиакатастроф или кораблекрушений. В большинстве случаев они гибнут или попадают в списки «пропавших без вести». Так, анализ 2,5 тыс. падений в воду самолетов, сбитых и потерпевших аварии над тропическим океаном в годы второй мировой войны, показал, что только в 38 случаях жертвы катастроф имели дело с акулами, а прямое нападение отмечено всего 12 раз. Эти сведения исходят, впрочем, от оставшихся в живых летчиков, которым, прямо скажем, очень повезло. Ведь в некоторых случаях кораблекрушения и авиакатастрофы сопровождаются массовой гибелью. В 1957 г., например, после аварии американского пассажирского самолета в 1000 милях к востоку от Гонолулу было обнаужено 19 трупов, изуродованных акулами, а катастрофа французского самолета над Атлантическим океаном мористее мыса Зеленого сопровождалась гибелью 63 пассажиров. Жертвами акул при крушении итальянского лайнера «Принципесса Мафальда», затонувшего в 1927 г. близ берегов Бразилии, стало, как полагают, около 300 человек. Еще больше людей (около 1000) погибло после того, как пароход «Нова Скотин» был торпедирован в 1942 г. в Индийском океане на подходе к Дурбану.
В официальных списках опасных акул значатся все перечисленные выше океанические виды, но документированные случаи их нападения на человека единичны (исключение составляет только акула мако). Следует отметить, однако, что два наиболее опасных вида — белая акула, или кархародон (Carcharodon carcharias), и тигровая акула (Galeocerdo cuvier), хотя и считаются приуроченными преимущественно к прибрежной зоне, вполне могут быть встречены и вдали от берегов. Об этом свидетельствуют, в частности нередкие поимки отдельных особей у отдаленных островов, в водах которых постоянные популяции обоих видов отсутствуют.
Кархародон — самая крупная из современных хищных акул, получившая очень широкую международную известность благодаря той рекламе, которую дал ей Голливудский кинобоевик «Челюсти». Наибольшая из измеренных особей этого вида, пойманная в водах Кубы, имела длину 6,4 ми массу 3300 кг, хотя, по-видимому, иногда встречаются и экземпляры длиной порядка 8м (Интересно отметить, что еще сравнительно недавно (возможно, уже в четвертичное время) в океане обитал еще один вид - гигантский кархародон (Carcharodon megalodon), треугольные, грубо зазубренные по краям зубы которого с 13-сантиметровой коронкой нередко находят в донных отложениях. Эта акула достигала в длину по меньшей мере 14,5 м (приведенные ранее размеры до 30 м были основаны на неправильно выведенном соотношении между длиной тела и зубов), и в ее пасти могли бы свободно разместиться несколько человек.). Обычно размеры кархародона меньше (длина 2—6 м, масса от 50 кг до 1,5т), причем самцы достигают зрелости при длине около 3 м, а самки при еще большей длине. Беременных самок пока не встречали (возможно, они держатся изолированно в ограниченных районах), а самые мелкие свободноживущие особи длиной 1,3—1,4 м ловились только в умеренно теплых водах. Кархародон ведет одиночный образ жизни, иногда надолго задерживаясь в том или ином участке океана (например, вблизи береговых рыбоперерабатывающих заводов и скотобоен). Обычно он держится у поверхности, но может опускаться и в глубокие слои: один экземпляр был пойман даже на глубине 1000 м. Этот вид широко распространен во всех океанах, преимущественно в умеренных широтах. Его нахождения отмечены в Средиземном и Красном морях, у атлантического побережья Европы до Ла-Манша) и Северной Африки, у Южной Африки, Сейшельских островов, в водах Австралии (кроме северного побережья) и Новой Зеландии, в западной части Тихого океана — от Филиппинских островов до Японского моря, у Новой Каледонии и Гавайских островов и у берегов Америки — от Калифорнийского залива до Аляски, от Мексиканского залива до Ньюфаундленда, от Панамского залива до Южного Чили и в водах Аргентины. Несмотря на столь широкое распространение, белые акулы всюду считаются довольно редкими, и об их жизни известно не так уж много. В результате телеметрического слежения за экземпляром длиной 4,6 м было установлено, что крейсерская скорость сытого кархародопа составляет около 3,2 км/ч при расходе энергии, не превышающем 0,2 ккал/кг/ч (расчет проведен по изменению температуры тела при переходе из холодной в более теплую воду: кархародоны способны к терморегуляции). Таким образом, наполнив желудок калорийной пищей (например, китовой ворванью), акула может полностью обеспечить свои энергетические потребности на месяц — полтора. Показано также, что внутрижелудочная температура у кархародона может на 7,4° превышать «забортную», причем, как предполагает Дж. Маккоскер, такое нагревание является произвольным.
Как показывают наблюдения за составом пищи, экземпляры длиной до 2 м питаются в основном некрупными акулами, костистыми рыбами (сельдь, скумбрия, морские окуни), кальмарами, крабами, но по мере роста кархародоп переключается на более солидные объекты, хотя не пренебрегает и относительно мелкими в местах их скоплений. Очень мощное зубное вооружение дает белой акуле возможность наносить своей добыче страшные повреждения и без особого усилия перекусывать кости и хрящи жертв, а широкая пасть и глотка позволяют проглатывать очень крупные куски. По-видимому, кархародон не особенно разборчив в выборе пищи, хотя чаще всего в желудках пойманных особей находили других акул, на которых он, очевидно, в основном охотится. При этом сравнительно небольшие акулы, не превышающие 2 м, проглатываются, как правило, неповрежденными, а более крупные, например гигантская акула, разрываются на куски. В состав излюбленной пищи крупных кархародонов входят также морские млекопитающие (дельфины, тюлени, котики, каланы), тунцы, морские черепахи. Этот вид не брезгует падалью (во времена прибрежного китобойного промысла атаки на загарпуненных китов не были редкостью) и даже отбросами, но несъедобные объекты (типа бутылок и тряпок) встречаются в желудках гораздо реже, чем у тигровой акулы.
Зарегистрировано немало случаев нападения белой акулы на людей, находящихся в воде, а также на парусные доски и лодки (избрав объектом нападения шлюпку, кархародон, как правило, атакует ее до тех пор, пока не затопит). Эти нападения отмечены, правда, не в открытых водах, а вблизи берегов — в бухтах и на пляжах (недаром в Австралии эта акула носит название «белой смерти»). При этом неоднократно указывалось, что большинство атак заканчивается гибелью объекта нападения и лишь немногим жертвам удается сохранить свою жизнь, отделавшись тяжелыми повреждениями. Статистика последних лет показывает, однако, что масштабы опасности были в значительной степени преувеличены. В водах Калифорнии, где за 32 года (с 1950 по 1982) отмечено 41 нападение кархародона на людей (в среднем 0,7 случая в год), лишь 4 из них привели к смертельному исходу (оказалось, что обычно акула делает лишь один укус).
Тигровая акула (максимальная длина около 5,5 м), обитающая в тропических и субтропических водах всех океанов, также относится к широко распространенным и крупным неритическим видам. В поисках пищи тигровая акула выходит в открытые воды, заходит в мелководные заливы и даже в устья рек, иногда попадаясь на глубине, не превышающей нескольких метров. Обычно она довольво медлительна, но становится быстрой и подвижной, когда учует пищу. Будучи очень прожорливой и неразборчивой в еде (считается, что это наиболее всеядный представитель щележаберных), эта акула пожирает крабов, лангустов, двустворчатых и брюхоногих моллюсков, кальмаров, различных рыб (в том числе и акул меньших размеров), морских черепах, панцири которых она легко рассекает мощными зубами, и любую доступную добычу. Для этого вида весьма обычен и каннибализм: однажды, например, крупная тигровая акула сожрала более мелкого представителя своего вида, попавшего на крюк тунцеловного яруса, но не насытилась и, схватив соседнюю наживку, оказалась пойманной. В желудках тигровых акул находили также упавших в воду перелетных птиц, бакланов, морских змей, куски дельфинов и крокодилов. Эта акула без излишней брезгливости относится и к падали и отбросам. Перечень съедобных объектов и несъедобных предметов, извлеченных из желудков, очень велик и включает кур, собак, кошек, свиней, ослов, обезьян, гиену, а также коровьи копыта, оленьи рога, мешки угля, куски дерева, консервные банки, бочонки, пивные бутылки, пачки из-под сигарет, разные плоды, картофель, кожаную обувь, тряпки, пластиковые мешки и др.
В тропических водах тигровая акула представляет собой едва ли не наиболее опасный вид. Известно очень много случаев, когда в желудках пойманных акул находили части тела человеческих жертв. Большая часть таких находок объясняется, вероятно, пожиранием трупов, но многие жертвы, несомненно, встретились с акулой еще живыми и здоровыми. Нападения отмечены во многих районах — у берегов Флориды, островов Карибского моря, Сенегала, Австралии, о-ва Новая Гвинея, островов Самоа и в Торресовом проливе. Список документально подтвержденных нападений тигровой акулы на людей и лодки достаточно внушителен, он включает 27 атак за несколько последних лет.
Несколько слов о том, как я сам отношусь к акульей опасности. В воде мне не раз приходилось видеть акул (в том числе двух-, трехметровых) метрах в 12—15, но они никогда не проявляли к моей персоне серьезного интереса. Все эти подводные встречи состоялись на коралловых рифах: крупные акулы любят крейсировать в толще воды у их внешнего края, там, где склон круто уходит вниз и растворяется в темно-синей бездне. Бывали случаи (особенно в первые встречи), когда казалось, что акула, заложив очередной плавный вираж, держит курс прямо на меня, но она всегда сворачивала на почтительном расстоянии. Поэтому пиетет (если не сказать страх), который я исходно испытывал к этим хищникам, постепенно исчез, хотя я всегда отмечаю каждое появление в поле зрения знакомого серого силуэта.
Явно агрессивное поведение акул я видел только один раз (если не считать происшествия в Бенгальском заливе, о котором я уже рассказывал). Было это при посещении необитаемого атолла Керолайн в архипелаге Центральные Полинезийские Спорады (они же острова Лаип) в 1974 г. Высадившись па небольшой островок, мы решили обойти его вокруг посуху и рассыпались в линию, прочесывая пляж в поисках выброшенных прибоем раковин. Я шел по щиколотку в воде а еще мористее (там глубина была по колено) держался гидрофизик К. Н. Федоров, бывалый мореход и путешественник. Вдруг он, взяв резкий старт, пулей выскочил па сушу. «Что это ты?» — «Да акула бросилась!» Я посмотрел в сторону лагуны: там действительно шныряло десятка полтора некрупных (от полуметра до полутора метров) рыбин с ярким черным пятном на плавнике. «Не бойся! Это черноперые рифовые — они вовсе не опасны». Теперь я остался крайним, но не успел сделать и десятка шагов, как совсем небольшая акула резко кинулась прямо на меня. Забыв свои уверения в ее безвредности, я в одно мгновение оказался рядом с Константином... Уже потом я прочитал, что акулы этого вида могут кусать за ноги людей, бредущих в воде, так как принимают эти ноги за свою привычную добычу. Считается, что лучшая защита от такого нападения — лечь в воду (но я не пробовал). Должен еще добавить, что, когда мы вечером садились в шлюпку для возвращения на «Академик Курчатов» (в это время начался прилив и пришлось добираться к ней по грудь в воде или вплавь), эти черноперые буквально вертелись между людьми (матросы безуспешно отталкивали их веслами), но никого не тронули.
Встреч с акулами-людоедами у меня не было, но я знаю, что они существуют и очень опасны. Я никогда не забуду судьбу двух моряков — моих товарищей по плаваниям на «Витязе», которые перешли работать на грузовое судно и погибли, когда этот транспорт, шедший с грузом руды из Австралии, попал в центр тайфуна и затонул в Филиппинском море. По тем останкам, которые, как говорили, были обнаружены через несколько дней плававшими в спасательных жилетах, опознать погибших не было возможности.
Как ни странно, в океанской пелагиали встречаются и представители такой, казалось бы, чисто донной группы щележаберных, как скаты. Среди них в первую очередь следует отметить пелагического хвостокола (Dasyatis violacea), который всесветно распространен в теплых водах на глубине до 240 м (обычен в верхней 100-метровой толще) и достигает 1,6 м в длину (ширина почти треугольного диска 80 см). Это — активный пловец-хищник, питающийся рыбами, креветками, кальмарами, иногда — медузами и размножающийся путем яйцеживорождения. В открытых водах могут быть встречены и другие скаты — гигантская манта (Manta birostris) (ширина диска до 6,7 м при массе более 2 т), планктофаг-фильтратор, в целом более характерный для неритической пелагиали тропиков, и преимущественно донные электрические скаты из рода Torpedo.
От щележаберных рыб представляется естественным перейти к их постоянным спутникам — рыбе-лоцману и прилипалам (см. рис. 6). Лоцман (Naucrates ductor) — один из немногих видов семейства ставридовых (Сагап-gidae), приуроченный в своем распространении к водам открытого океана. Эта рыба, максимальная длина которой не превышает полуметра, всегда держится рядом с каким-нибудь крупным пловцом (как правило, вместе с пелагической акулой) и не отстает от него ни при каких обстоятельствах. Как было показано академиком В. В. Шулейкиным, плавание относительно небольшой рыбы в слое трения, окружающем тело быстро движущегося «хозяина» (так называемое лоцманирование), позволяет такой рыбе плыть со скоростью хозяина при очень малых энергетических затратах. Лоцман отходит от акулы только при приближении к объекту, достойному специального обследования — другой большой рыбе, судну, островку плавучих водорослей, даже к какому-нибудь одиночному дрейфующему предмету — бревну, ящику или кокосовому ореху.
По-видимому, из лоцманирования развилось и контактное прикрепление к сопровождаемому объекту при помощи специальной присоски, столь характерное для рыб-прилипал (семейство Echeneidae с 8 видами). Эта присоска образовалась вследствие трансформации спинного плавника и представляет собой расположенный на плоской поверхности головы и передней части спины удлиненный овальный диск, окруженный упругим кожпо-мышечным валиком. Видоизмененные плавниковые лучи расщеплены по длине на уплощенные половинки, которые шарнирно прикреплены боковой стороной к телу и расположены внутри диска перпендикулярно продольной его оси. В результате образуется некоторое подобие жалюзи с вращающимися пластинами. При свободном плавании рыбы эти пластины лежат плашмя, а когда присоска прижата к какой-либо достаточно гладкой поверхности, они поворачиваются и занимают стоячее положение. При этом в полости, ограничейной краевым валиком диска, образуется серия камер с частичным вакуумом. Такое устройство присоски обеспечивает очень надежное прикрепление, и для отцепления рыбы нужно продвинуть ее вперед, чтобы опустить стоящие пластины и ослабить вакуум. Если же тянуть прилипало за хвост, то присасывание будет только усиливаться. Интересно, что прилипалы могут, подобно улитке, ползущей по стеклу, скользить по подстилающей поверхности, не отрываясь от нее, но лишь последовательно изменяя положение отдельных пластин присоски.
С помощью присоски прилипалы (длиной 30—80 см) прикрепляются к различным «хозяевам» — крупным рыбам, черепахам, китам, а иногда и к морским кораблям. Степень связи с «хозяином» и способность к активному передвижению и самостоятельной жизни сильно варьируют у разных видов прилипал. Одни из них (например, Phtheirichthys lineatus) обычно свободно плавают в поверхностных слоях воды и почти не пользуются присоской, другие прикрепляются к наружной стороне тела «хозяев», третьи локализуются в жаберной полости крупных рыб. Такие прилипалы, как акулья ремора (Remora remora), вообще вряд ли могут существовать сами по себе. Будучи снятыми с акулы и помещенными в аквариум, они начинают «тяжело дышать», совершая до 240 дыхательных движений в минуту. Даже дыхание приспособлено у них к постоянной жизни в прикрепленном состоянии (реморы покидают акулу только для питания), так как при движении «хозяина» вода свободно проходит через рот к жабрам «наездника» без специальных усилий. Часто реморы настолько плотно держатся на акуле, что остаются на ней даже тогда, когда ее поднимают на палубу.
Почти все прилипалы довольно специфичны в отношении выбора «хозяев»: есть виды, живущие только на акулах или только на китообразных, а самый мелкий вид (Rеmorina albescens) использует в качестве постоянного убежища ротовую и жаберную полости огромных скатов мант. Прилипалы, находящиеся в симбиотических отношениях с одиночно живущими рыбами, по-видимому, прикрепляются к ним парами, состоящими из самки и самца. Смысл прикрепления состоит в облегчении передвижения, и мнение о питании этих рыб остатками пищи «хозяев» и отходами их жизнедеятельности мало соответствует действительности. Прилипалы питаются в основном свободно живущими планктонными организмами и в меньшей степени эктопаразитами «хозяев». Их отношения с «хозяевами», несомненно, носят характер симбиотических связей, чем л объясняется крайняя редкость попадания прилипал в желудки рыб, которых они сопровождают.
Как прилипало находит себе хозяина? Кто его знает... Но вот какой случай произошел с моим хорошим другом В. Г. Нейманом на одном из островов Индийского океана. Группа участников экспедиции занималась сбором образцов коралловой фауны на прибрежном рифе, а мадрепоровые кораллы, надо сказать, обладают неприятным свойством оставлять долго не заживающие даже при самом легком контакте с ними царапины. Поэтому все были экипированы в полном соответствии с техникой безопасности, и на ногах Виктора красовались прекрасные резиновые сапоги. Вдруг он заметил, что какая-то рыба проявляет самый непосредственный интерес к его обуви. Приглядевшись, он узнал прилипало, пытающегося прикрепиться к гладкой поверхности голенища. Не будучи достаточно знакомым с повадками этих рыб, Виктор представил себе, что вслед за этим «наводчиком» незамедлительно появится и акула внушительных размеров, и, отбиваясь на ходу ломиком, которым пользовался для работы на рифе, с максимальной скоростью устремился к берегу. Прилипало преследовало его почти до самого пляжа, не желая, по-видимому, отказываться от столь полюбившегося ему «хозяина», и только упустив его из виду, отправился на поиски нового объекта своей привязанности.
А вот еще одна история, связанная с этими рыбами. Помню, как в Соломоновом море один и тот же прилипало фтейрихтис - его можно было опознать по светлому шраму на спине — систематически появлялся на световых станциях в течение нескольких ночей подряд. Нужно думать, что переходы между этими станциями он преодолел безбилетным пассажиром «Витязя», как бы на подножке вагона.
Среди эпипелагических рыб резко выделяются своим странным, «совсем не рыбным» экстерьером рыбы-луны (семейство Molidae). К этой группе относятся всего 4 вида — относительно некрупная (до 80 см) удлиненная луна (Ranzania laevis) и три гиганта, достигающие 3 м в длину и более 2 т по массе. Наиболее распространенный вид — обыкновенная луна (Mola mola) (ее латинское название переводится как «жернов», или «мельничный камень») имеет дисковидное кургузое тело бурого цвета, состоящее, как кажется на первый взгляд, из одной только головы (рис. 7). Несмотря на очень внушительные размеры, луна-рыба, не способная противостоять течениям, должна рассматриваться как планктонное животное. Закованное в жесткую толстую шкуру тело и отсутствие хвоста сильно ограничивают возможность активного движения, которое осуществляется только посредством вертикальных плавников и, возможно, ограниченно подвижной задней пластиной, действующей как кормовое весло.

Рис. 7. Обыкновенная (1) и удлиненная (3) рыбы-луны и опах (3)
В толще воды рыба-луна, может быть, плавает в вертикальном положении, но у поверхности, где она появляется нечасто, обычно лежит на боку или почти на боку. Рыба-луна — планктофаг, значительную часть ее пищи составляют сальпы, медузы и гребневики. Она считается рекордсменом среди всех рыб по плодовитости (300 млн икринок!). Все рыбы-луны размножаются только в центральных частях субтропических круговоротов, но взрослых рыб часто заносит далеко за пределы теплых вод (вплоть до Баренцева моря и залива Аляска).
Рыбы-луны относятся к отряду скалозубообразных рыб, большинство представителей которого обитает на мелководьях тропических морей. В его составе есть, однако, еще два настоящих эпипелагических вида, широко pacпространенные в тропической зоне, — океанический иглобрюх (Lagocephalus lagocephalus) длиной до 60 см и океаническая еж-рыба (Diodon eydouxii) длиной до 27 см.
Оба эти вида принадлежат к преимущественно прибрежным родам.
Отдаленное внешнее сходство с рыбами-лунами имеет совсем не родственный им пятнистый опах (Lampris guttatus) — такая же высокотелая, сжатая с боков рыба (см. рис. 7), имеющая, однако, вполне развитый хвостовой стебель и очень яркую окраску: спина сине-зеленая, бока пурпурно-золотистые с многочисленными серебряными пятнами, брюхо розовое, все плавники ярко-красные. Опах достигает длины около 1,5 м и массы 60 кг. Он широко распространен в теплых водах всех океанов (близкий вид — низкотелый опах L. immaculatus — обитает только в водах Субантарктики) и встречается от поверхности до глубины 200 м и более. Эти рыбы отличаются очень своеобразным способом плавания: основным движителем являются грудные плавники, совершающие частые машущие движения (как крылья птиц в полете).
Низкотелый опах, отличительной особенностью которого служит отсутствие светлых пятен на теле, был описан по экземпляру из вод Южной Африки еще в начале нынешнего столетия, но долгое время не признавался самостоятельным видом. В начале 80-х годов такая однотонно окрашенная рыба длиной около метра была поймана у острова Южная Георгия при лове криля (он же «антарктическая креветка») и в замороженном виде привезена в Калининград. Необыкновенный облик этого экземпляра - удлиненное толстое тело, нестандартная окраска, странное положение плавников,- к счастью не остался незамеченным Ефимом Кукуевым, сотрудником АтлантНИРО, в то время аспирантом Института океанологии. Рыба была переправлена в Москву (сейчас она хранится в Зоомузее МГУ), и мы с Ефимом опубликовали после ее изучения статью, доказывающую существование двух видов ламприсов. Оказалось, что низкотелый опах и до нас попадал в руки исследователей, но не распознавался ими как особый вид. Один из ихтиологов, участвовавший в дегустации экземпляра выловленного в море Беллинсгаузена, даже сообщил нам, что на вкус эта диковина гораздо приятнее, чем хорошо знакомый ему пятнистый вид (который тоже является деликатесной рыбой)..
К характернейшим обитателям поверхностных слоев тропического океана относятся и корифены. Большая корифена, или золотая макрель (Coryphaena hippurus) (рис. 8), достигает длины 4,8—2 м и массы около 30 кг (обычно не более 10 кг). Ее окраска отличается чрезвычайной яркостью: спина окрашена в сверкающий сине-зеленый цвет, бока и брюхо отливают серебром или золотом, имеют красноватый оттенок и украшены синими пятнами, хвостовой плавник ярко-желтый, спинной — темно-синий. Корифены — активные хищники, поедающие пелагических рыб и кальмаров. Основную их пищу составляют летучие рыбы, и иногда можно наблюдать, как корифена быстро плывет вслед за летящей в воздухе летучкой и хватает ее в тот самый момент, когда она опускается в воду. Интересно отметить, что отношения типа хищник — жертва складываются между корифеной и летучими рыбами еще в ранний период жизни: в опыте малек размером всего 26 мм, помещенный в аквариум с молодью летучих рыб длиной около 12 мм, незамедлительно проглотил одну рыбку и атаковал другую, прежде чем был отсажен. Этот вид отличается очень быстрым ростом — половая зрелость наступает в возрасте 1 года при длине 55 см, а продолжительность жизни не превышает 5 лет.
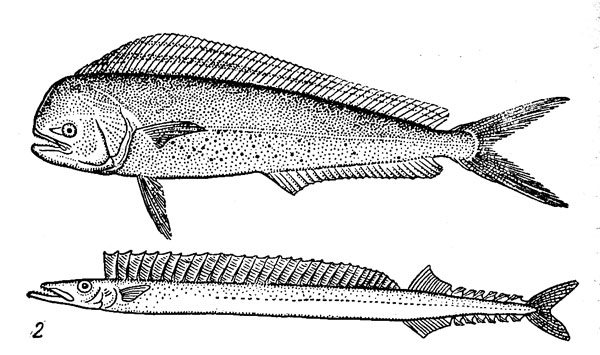
Рис. 8. Корифена (1) и змеиная макрель (3)
Корифены, как и некоторые другие пелагические рыбы, нередко держатся под плавником (скопления плавучих водорослей, наземная растительность, вынесенная в море, и другие предметы берегового происхождения). Под плавником корифены находят обильную пищу в виде укрывающейся здесь мелкой рыбешки. По этой же причине эти рыбы сопровождают плоты (они вносили немалое оживление в однообразную жизнь Т. Хейердала и его спутников при плавании на «Кон-Тики») и подходят к дрейфующим в открытом океане кораблям. Такое поведение корифен используется даже для их промысла. У берегов Южной Японии в море выставляют специальные бамбуковые плотики, под которыми собираются корифены, облавливаемые потом кошельковыми неводами.
Участники экспедиций на океанологических судах не очень-то избалованы рыбной ловлей. Правда, поймать длиннокрылую акулу не составляет большого труда (была бы наживка!), но ведь для этого не нужно и искусства. Совсем другое дело корифена... Когда подходит момент взятия на борт гидрологического буя, на тросе которого несколько суток болтались в воде самописцы течений, судовые рыбаки-любители находятся в нетерпеливом ожидании. Крепкие тунцовые крючки загодя наживлены бережно хранившимся в холодильнике куском кальмара или летучки, капроновые шнуры и толстые лески длиной по 20-30 м набраны аккуратными кольцами и крепко зажаты в руке, горящие глаза прикованы к приближающейся вехе буя... Все в полной готовности, ио все равно так неожиданен долгожданный крик: «Корифены!» И вот большая, удивительно красивая рыбина (иногда две или даже маленькая стайка) грациозно скользит в голубой воде вдоль борта, проверяя, не затаилось ли под его защитой что-нибудь годное в пищу. Крючки летят вслед рыбе, падают рядом с ней и перед самым ее носом, но хищник не обращает на них внимания: никому не ведомо, отчего зависит выбор приманки. Вдруг корифена круто поворачивает и резко ускоряет и без того стремительный «бег»... Мгновение, и паживка схвачена... Резкий рывок, бросок в сторону, взлет в воздух, еще один-два броска, и вот леска натянулась... Несколько секунд борьбы, и рыба поднята на борт. Вьющаяся па палубе, опа переливается всеми цветами радуги, мепяя окраску от сине-зеленой до золотисто-желтой... Все копчено. Ловцы расходятся по рабочим местам, надеясь, что им повезет в следующий раз.
Основной пищевой конкурент корифен — змеиная макрель (Gempylus serpens) (см. рис. 8) также питается в основном летучими рыбами и кальмарами. Эта тонкотелая зубастая рыба длиной до 1 м довольно обычна в теплых водах всех океанов. Днем она держится на значительной глубине, а ночью поднимается к поверхности и хорошо привлекается источником искусственного освещения. Атакуя свою добычу снизу (иногда почти по вертикали), змеиная макрель нередко совершает довольно высокие — 2—3 м по глазомерной оценке — прыжки над поверхностью воды.
Змеиная макрель принадлежит к подотряду скумбрневидных (Scombroidei), в составе которого немало океанических рыб. Одним из наиболее примитивных видов подотряда считают обычно гастерохизму (Gasterochisma melampus), которая в отличие от других его представителей имеет очень крупную чешую и лишена килей на хвостовом стебле. Эта рыба, достигающая длины 1,7 м и более, обитает только в умеренноводной эпипелагиали южного полушария (в поясе между 30 и 50° ю. ш.), предпочитая температуру порядка 8—10°.
Интересную группу скумбриевидных представляют тунцы — рыбы с идеально приспособленным для скоростного плавания веретеновидным телом. Их место в системе рыб вызывало до недавнего времени много разногласий. Дело в том, что у тунцов имеется совершенно уникальная система подкожных кровеносных сосудов (они снабжают кровью боковую мускулатуру), и эта особенность анатомии, впервые обнаруженная японским морфологом К. Кисинуйе, послужила основанием для предложенного им выделения тунцов в особый отряд, принятый вслед за этим автором академиком Л. С. Бергом и некоторыми другими исследователями. Такая точка зрения сейчас полностью отвергается, и близкое родство тунцов с неритическими скумбриями, королевскими макрелями и пеламидами, вместе с которыми они входят в состав семейства Scombridae (в нем, по Б. Коллетту и К. Науэн, 15 родов и 49 видов), никем не оспаривается. Необычность кровообращения представляет собой лишь одно из приспособлений тунцов к столь характерному для них продолжительному быстрому (до 90 км/ч) плаванию, сопряженному с большими энергетичесними затратами. В моменты, требующие максимального расхода энергии, температура тела тунцов значительно повышается — разница между ней и температурой окружающей среды может достигать нескольких градусов. Подкожная система кровеносных сосудов, лучше всего развитая у синего тунца, обеспечивает специальную доставку кислорода к тем мышцам, которые выполняют наибольший объем работы ( По мнению А. Н. Заикииа, эта система может обеспечивать также увеличение упругости тела путем эрекции (наполнения кровью) приповерхностных тканей, что позволяет рыбе значительно увеличить частоту колебательных движений хвоста. Аналогичный механизм был недавно обнаружен у китообразных.). Достижению той же цели способствует и высокая кислородная емкость крови этих рыб, у которых содержание гемоглобина в эритроцитах доходит до 21% (у пеламид оно не превышает 8—14%).
Постоянное плавание — наиболее характерная особенность биологии тунцов, и они прекрасно приспособлены к такому образу жизни. Как показал Ю. Г. Алеев, у этих скоростных пловцов (а также у скумбрий, пеламид, меч-рыбы, марлинов) функция движителя целиком перенесена на хвостовой плавник, а короткое обтекаемое тело практически не выполняет локомоторной работы. При остановке у этих рыб даже затрудняется дыхание, так как вода попадает в жаберную полость лишь через постоянно открытый рот при поступательном движении. Таким образом, специализация как неритических, так и океанических тунцов направлена на достижение высоких скоростей плавания и активное перемещение на дальние расстояния.
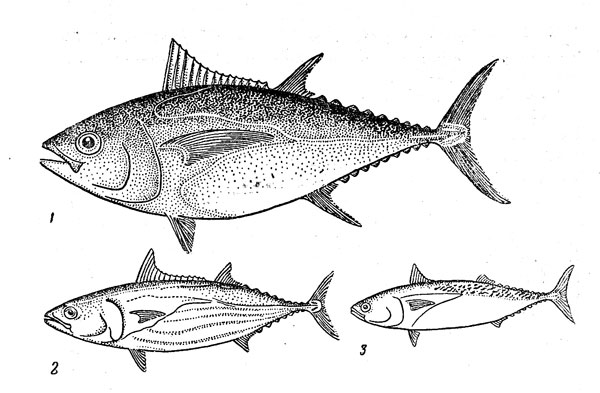
Рис. 9. Океанические тунцы: большеглазый (1), полосатый (3), макрелевый (3).
Среди океанических тунцов преобладают крупные виды, и лишь один скипджек, или полосатый тунец (Katsuwonus pelamis) (рис. 9), сравнительно невелик: 50—80 см при массе 3—10 кг (отдельные особи достигают, впрочем, длины 1 м и массы порядка 35 кг). Скипджек — характернейшая рыба поверхностных слоев открытого океана, встречающаяся, однако, и в прибрежных водах, особенно вблизи коралловых рифов. Он обитает во всех теплых морях, а в некоторых районах (например, в северо-западной и восточной частях Тихого океана) предпринимает довольно значительные сезонные перемещения и проникает в летнее время года в умеренно тепловодные участки. Встречается при температуре 15—30°, по нерест идет только в прогретых до 25° и выше водах. Для этого вида типичен стайный образ жизни. Стаи полосатого тунца развивают большую скорость (до 45 км/ч) и могут быть очень внушительными: иногда они включают десятки тысяч особей. Их пища чрезвычайно разнообразна в видовом отношении и состоит из сравнительно небольших рыб, кальмаров и ракообразных.
Род настоящие тунцы (Thunnus), объединяющий наиболее крупных представителей семейства, содержит шесть видов. Преимущественно в субтропических, реже в умеренно теплых и тропических водах всех океанов обитает синий, или обыкновенный, тунец (Th. thynnus), стайная рыба, обычно встречающаяся вдали от берегов. Это — наиболее крупный тунец, иногда превышающий в длину 3 м при рекордной массе 679 кг (обычный размер не более 1 2 м). Он обладает быстрым темпом роста и достигает длины 1 м в возрасте трех лет и 2 м в возрасте 7—9 лет, половой зрелости достигает в 4—5 лет. Его пища состоит из рыб и беспозвоночных, достаточно обильных в районе кормежки. Индивидуальная плодовитость крупных особей доходит до 10 млн икринок. Синий тунец во всех частях видового ареала совершает сезонные миграции, перемещаясь в основном вдоль берегов. Миграции в открытых водах изучены плохо, но известно, что отдельные особи могут совершать трансокеанские перемещения. Отмечены, в частности, случаи, когда рыб, помеченных у берегов Мексики, вторично ловили в районе Токио — в 5800 морских милях от места мечения. В Атлантическом океане особи с метками, прикрепленными у Флориды, ловились в Бискайском заливе.
В теплых и умеренно теплых водах обычно при температуре 13—25° обитает также длинноперый тунец, или альбакор (Th. alalunga), который никогда не показывается в поверхностных горизонтах в экваториальной зоне. В отличие от синего тунца эта рыба редко появляется у берегов. Молодые особи живут у самой поверхности, нередко собираясь в стаи под плавучими объектами, в том числе под скоплениями саргассов. Взрослые особи тяготеют к значительной глубине, которая в зависимости от температуры и содержания в воде кислорода (лимитирующие величины составляют 9,5° и 2 мл/л) варьирует до 380 м в Тихом океане до 600 м в Атлантическом. Альбакор нерестится у границ тропической области в весеннелетнее время соответствующего полушария.
Плодовитость этой рыбы до 2,5—3 млн икринок. Длинноперый тунец растет медленнее, чем тропические виды настоящих тунцов. Он становится половозрелым в возрасте 4—5 лет при длине около 90 см, а максимальные размеры достигают 1,3 м при массе 45 кг. Эта рыба тоже совершает значительные по протяженности миграции. Предполагается, например, что в северной половине Тихого океана стаи неполовозрелых альбакоров постоянно перемещаются между берегами Америки и Японии, придерживаясь вполне определенных путей. Эти миграции на некоторых участках были прослежены при помощи мечения, что дало возможность определить их скорость, составляющую в среднем 16—17 морских миль в сутки. Дальние миграции свойственны, по-видимому, и другим популяциям альбакора.
К типичным океаническим рыбам, почти не заходящим в прибрежные воды, относится и большеглазый тунец (Th, obesus) (см. рис. 9). Он широко распространен в тропических и субтропических районах всех океанов (оптимальная температура 17—22, предельная —13 и 29°), повсюду придерживается довольно больших глубин (до 250 м и более) и обычен в слое температурного скачка. У поверхности живут лишь молодые особи, образующие довольно плотные стаи. Особенно большое значение в пище этого вида имеют глубоководные и полуглубоководные рыбы, кальмары, пелагические осьминоги, крупные креветки. Большеглазый тунец достигает длины 45—50 см к концу первого года жизни, 70 см в два года и 155 см в шестилетнем возрасте, причем половая зрелость наступает при длине 100—130 см. Самый крупный из известных экземпляров этого вида был пойман у берегов Перу: его длина 236 см, масса 197 кг.
Для тропических вод океанской пелагиали очень характерен желтоперый тунец (Th. albacares), получивший свое название благодаря оранжево-желтой окраске спинного и анального плавников. Границы ареала этого вида, населяющего воды всех океанов, более или менее соответствуют положению изотермы 20° в наиболее теплое время года. Он обычен в поверхностном слое и редко опускается глубже 100 м. Кормовой спектр желтоперого тунца очень разнообразен, что, несомненно, связано с отсутствием у этого вида (как, впрочем, и у других тунцов) избирательности в питании. Достаточно отметить, что в желудках этого хищника были обнаружены рыбы, относящиеся к 50 различным семействам. Желтоперый тунец достигает 2 м длины и более и массы 175 кг (обычно значительно меньше). Этот вид характеризуется очень быстрым ростом: годовой прирост длины, по данным вторичных поимок рыб, составляет 20—40 см. Половое созревание наступает при достижении длины 50—60 см, а плодовитость составляет около 1 млн икринок у небольших рыб и до 8,5 млн у крупных особей. Нерест желтоперого тунца в тропической зоне идет во все сезоны года, а у ее краев ограничивается летники месяцами. Трансокеанских перемещений, подобных отмеченным для синего тунца и альбакора, желтоперый тунец не совершает.
Группу «мелких тунцов», которых считают перспективными объектами океанического промысла, образуют ауксиды, или макрелевые тунцы (род Auxis), и малые тунцы (Euthynnus). Представители обоих родов более характерны для неритических вод, но оба вида ауксид (это самые мелкие из тунцов, редко имеющие более 50 см в длину) попадаются в любом удалении от берегов. Они обитают у поверхности в наиболее прогретых районах всех океанов (температура воды 22—31°) и ведут стайный образ жизни. Из малых тунцов в открытый океан проникает, по-видимому, только один восточнотихоокеанский вид (Е. lineatus), обычная длина которого не превышает 65 см. Другой массовый океанический вид — южный тунец (Allothunnus fallai) (длина до 1 м, масса до 10 кг) распространен цир-кумглобально между 20 и 35° ю. ш. и питается главным образом эвфаузиевыми рачками.
Я уделил здесь тунцам так много внимания, поскольку именно они могут служить, на мой взгляд, символическим олицетворением некоей «идеальной» эпипелагической рыбы. Любой тунец независимо от видовой принадлежности и размера превосходно приспособлен к жизни в безбрежной и бездонной толще океана - он быстр, резок в движениях, обладает высокой маневренностью, неутомим.., Удивительна форма его обтекаемого тела: оно действительно веретеновидное, как справедливо пишут в определителях и описаниях, но, чтобы по-настоящему осознать, что это значит, нужно обязательно увидеть такое «веретено» воочию. Особенно впечатляет почти идеально круглое в любом участке поперечное сечение головы и туловища (ведь у рыб с привычной нам «рыбовидной» формой тела оно всегда сильно уплощено с боков). Поразительно в изящество исключительно тонкого хвостового стебля перед могучим движителем — прочным и жестким хвостовым плавником полулунной формы, который способен вибрировать с невероятной частотой. Первый тунец которого я увидел живым,— это была маленькая ауксида длиной сантиметров в 30,- навсегда остался в моей памяти, а следующие гиганты (мне, правда, не приходилось иметь дело с очень большими экземплярами, но и метр от конца рыла до выемки хвоста — не так уж мало!) немного добавили к исходному образу,

Рис. 10. Меч-рыба (1) и большой синий марлин(2)
Еще одна скумбриевая рыба — ваху (Acanthocybium solandri) (наиболее крупные особи имеют длину 2,1 м и достигают массы 80 кг) также может быть встречена и у берегов (преимущественно над свалом глубин), и в открытом океане, Ваху ведет одиночный образ жизни и редко образует стайки. Питается рыбой (миктофидами, летучками, мелкими тунцами) и кальмарами.
Особое место в подотряде скумбриевидных занимает меч-рыба (Xiphias gladius) (рис. 10), получившая свое название благодаря сильно удлиненной и уплощенной верхней челюсти, которая имеет форму меча длиной до трети длины всей рыбы. Этот вид очень широко распространен в тропических и субтропических районах всех океанов, а при кормовых миграциях кое-где проникает и в умеренно теплые воды. Во время откорма меч-рыба встречается в широком диапазоне температур (от 12—15° и выше), по нерест происходит только в сильно прогретой воде (не ниже 23,5°). Плодовитость очень велика — у относительно небольшой самки массой 68 кг насчитывалось около 16 млн икринок. Меч-рыба быстро растет (она достигает длины 50—60 см в массы 4 кг уже на первом году), половое созревание происходит при длине 140—170 см в возрасте пяти-шести лет, а полноразмерные особи достигают 4— 4,5 м при массе, превышающей 400 кг («рекордный» показатель — 550 кг). Меч-рыба — быстрый и активный пловец, развивающий скорость до 130 км/ч. Ее пища состоит преимущественно из рыб и головоногих моллюсков. Фактически она поедает любых животных, встречающихся на пути. В описке кормовых объектов этого вида значатся не только сравнительно мелкие приповерхностные и полуглубоководные рыбы (во время питания меч-рыба может опускаться на большую глубину), но и крупные хищники типа тунцов и даже акул (последние, правда, попадаются в; пище сравнительно редко). В отличие от марлинов и парусников, копьевидная верхняя челюсть которых имеет лишь гидродинамическое значение, «меч» рассматриваемого вида используется и для поражения добычи. Рыбы и кальмары, которых находят в желудках меч-рыб, довольно часто перерублены на части или имеют другие следы повреждений, нанесенных мечом. Некоторые черты поведения меч-рыб до сих пор не получили объяснения, в том числе случаи нападений на шлюпки и боты. Иногда обломки мечей извлекали даже из корпусов крупных судов. Причина этих нападений отнюдь не ясна, и все толкования такого поведения (простая случайность, обусловленная быстрым плаванием; ошибочное принятие лодки за акулу или кита; «бешенство» отдельных экземпляров) носят чисто умозрительный характер.
Близкие к меч-рыбе копьерылые рыбы (семейство Istiophoridae, объединяющее, по И. Накамура, 3 рода и 11—12 видов) также принадлежат к разряду активных хищников, развивающих в воде огромную скорость (порядка 100—130 км/ч). Быстрому плаванию очень способствует форма тела: их «копье» — вытянутая и заостренная, округлая в поперечном сечении и имеющая грубошершавую терковидную поверхность верхняя челюсть — служит, как показано В. В. Овчинниковым, для турбулизации набегающего потока и сильно уменьшает лобовое сопротивление при движении в плотной водной среде, а хвостовые кили увеличивают поперечную жесткость хвостового стебля и выполняют к тому же роль горизонтальных стабилизаторов (этой же цели служат у некоторых марлинов жесткие грудные плавники). Интересно отметить, что техническая мысль воспроизвела вполне аналогичные устройства в конструкциях сверхскоростных реактивных самолетов, также имеющих заостренный нос и хвостовые стабилизаторы. Представители этого семейства особенно характерны для пелагиали тропических и субтропических морей и океанов.
Полосатый копьеносец (Tetrapturus audax), длина которого достигает 3,5 м, а масса — 220 кг, обитает в Тихом и Индийском океанах и встречается преимущественно в субтропических водах при температуре 20—25°, Нерест этого вида в Тихом океане происходит на периферии тропиков летом соответствующего полушария, так что северная и южная популяции вида совершенно различны по сезону и месту икрометания (в Индийском океане он размножается повсеместно). Плодовитость полосатого копьеносца около 14 млн икринок, развивающихся в толще воды. Он питается различной рыбой — макрелещукой, сайрой, светящимися анчоусами, змеиными макрелями, алепизаврами и др., а также кальмарами и крупными ракообразными. Полосатый копьеносец предпринимает значительные горизонтальные миграции, в ходе которых эта рыба перемещается в более высокие широты в теплое время года и возвращается в тепловодпые районы ареала зимой.
Большой синий марлин (Makaira mazara) (см. рис. 10), также обитающий только в Тихом и Индийском океанах, несомненно, принадлежит к наиболее крупным костистым рыбам, живущим в наше время. Он достигает более 5 м длины и в ярусных уловах может иметь массу более 900 кг, хотя рекордный для спортивного рыболовства экземпляр, пойманный близ пляжа Вайкики (Гавайские острова), вытянул «всего» 818 кг.(Этот рекорд считается, впрочем, «неофициальным», так как в вываживании рыбы и ее подъеме на борт катера участвовал не один рыбак. Официальный рекорд значительно ниже: 523 кг. Такая рыба поймана у о-ва Гуам.) Это была, несомненно, самка, так как максимальная масса самцов не превышает 150 кг. Половое созревание самцов происходит при массе около 40 кг, самок — 50 кг, а в промысловых уловах резко преобладают сравнительно некрупные особи, весящие не более 100 кг.
Синий марлин распространен в тропических водах всех океанов и предпочитает температуру не менее 26— 27°. Это — типичная океаническая рыба, относительно редкая у берегов и держащаяся, как правило, в поверхностных слоях, только над термоклином. В желудках синего марлина чаще всего находят тунцов (особенно полосатого) и кальмаров, составляющих его излюбленную пищу, а также других крупных рыб — корифен, гемпилид, ставридовых и др.; глубоководных рыб этот хищник практически не поедает в отличие от тунцов и копьеносцев. Вид характеризуется очень высокой плодовитостью: у самки массой 130 кг было насчитано 27 млн, а у более крупной (420 кг) рыбы — 99 млн ооцитов. Нерест происходит в тропической полосе и продолжается в течение всего года в экваториальных районах, но, возможно, ограничен летними месяцами; на периферии области размножения. Систематических миграций эта рыба не совершает и значительных скоплений не образует. В Атлантическом океане вид замещен очень близким к нему атлантическим синим марлином (М. nigricans), максимальная длина которого не превышает 4 м, а масса — 600 кг (официальный любительский рекорд 581 кг). Это и есть та самая «большая рыба», которую выловил герой знаменитой повести Э. Хемингуэя «Старик и море». Нельзя не отметить, что все детали поведения марлина и других упоминаемых в этой замечательной книге рыб — летучек, корифен, акул — описаны автором с подлинно научной точностью.
Еще одна характерная для эпипелагиали группа (ее изучение сильно продвинуто работами Р. Хэдрича) — стрем матеевидные (подотряд Stromateoidei), в которой есть впрочем, и неритические виды. Для мальков строматеевидных очень характерно совместное обитание с медузами, сифонофорами или пелагическими оболочниками, под которыми они скрываются от хищников. Иногда рыбки поселяются даже в пищеварительной (гастроваскулярной) полости медуз или, как маленькие алеты (род Tetragonurus), внутри колонии пиросом огнетелок. Такое сожительство дает им не только хорошее укрытие, но и обеспечивает стол: здесь они имеют возможность питаться остатнами пищи своих «хозяев», а в некоторых случаях обкусывать некоторые части их тела.
В ходе приспособления к такому образу жизни у этих; рыб выработался особый механизм защиты от ядоносных стрекательных клеток медуз и сифонофор. Мальки строматеевидных значительно меньше других рыб восприимчивы к токсическим веществам своих «хозяев», хотя вовсе не обладают полным иммунитетом к ним. К тому же их тело часто покрыто толстой слизистой оболочкой. Во взрослом состоянии представители этого подотряда уже не нуждаются в подобном покровительстве, но многие из них продолжают «сохранять интерес» к мягкотелым беспозвоночным, используя их теперь исключительно в качестве объекта питания.
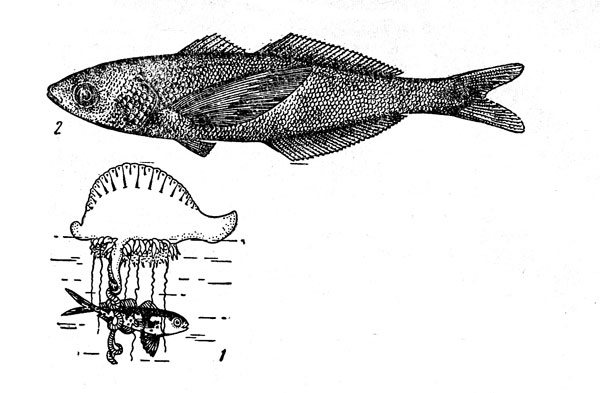
Рис. 11. Номей: малек (рыба-пастушок) под сифонофорой физалией (1) и взрослый экземпляр(2).
Особенно большую известность благодаря облигатном комменсализму молоди с плавающими на поверхности сифонофорами физалиями, яд которых опасен даже для человека, получил номей (Nomeus gronovii) (рис. 11), распространенный в теплых водах всех океанов. Пестрые мальки этого вида, имеющие яркие темно-синие пятна на серебристом теле, как правило, не встречаются вдали от сифонофор и получили в связи с этой особенностью поведения название «рыбы-пастушки». Пища физалий состоит из молоди рыб, которую они убивают стрекательными клетками, расположенными на ловчих шупальцах. Но маленькие номей относительно иммунны к токсину физалий, хотя и стараются при этом избегать непосредственного контакта с их щупальцами (известны, впрочем, и отдельные случаи поедания рыбок их опасными «хозяевами»). Несмотря на то что под сифонофорами отмечались лишь мальки и неполовозрелые особи длиной от 1 до 15 см, долгое время считалось, что сожительство номея и физалий имеет постоянный в течение всей жизни характер. Недавно было показано, однако, что взрослые рыбы этого вида живут самостоятельной жизнью, о чем свидетельствуют поимки большим пелагическим тралом экземпляров длиной до 35 см, резко отличающихся от более мелких однотонно-бурой окраской и совсем другими пропорциями тела.
Кубоглавы (род Cubiceps с 10 видами), обладающие очень длинными грудными плавниками, также обычны во всех районах, тропической и субтропической зон. Распространенный в Северо-Восточной Атлантике обыкновенный кубоглав (С. gracilis) в раннем возрасте держится у поверхности и часто ловится под медузами. Половое созревание наступает при достижении длины 20 см, но и после этого рост рыбы продолжается и максимальная длина достигает 1 м. Такие крупные особи живут в подповерхностных слоях. Другие виды кубоглавов значительно мельче: многие из них, например С. pauciradiatus, не превышают 20—25 см.
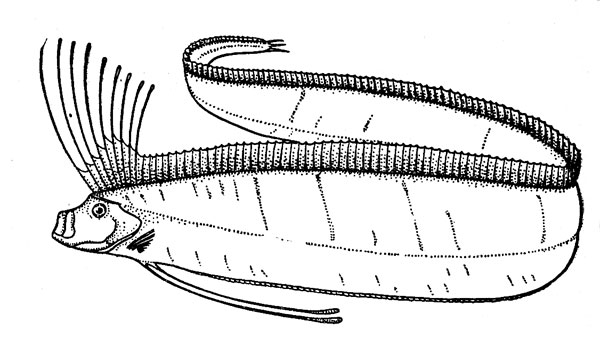
Рис. 12. Сельдяной король.
Некоторые представители рассматриваемого подотряда, например центролоф (Centrolophus niger), живущий в умеренно теплых водах Северной Атлантики и южного полушария, по мере роста существенно меняют глубину обитания. Мальки этого вида держатся у поверхности и, подобно другим строматеевидным, сопровождают крупных медуз. Они быстро растут (в аквариуме рыбка длиной 2 см всего за пять месяцев увеличила размер в 8,5 раз). Взрослые центролофы — однотонные черно-коричневые рыбы, длина которых достигает 1,2 м, — держатся только в нижних горизонтах эпипелагиали.
Для нижней эпипелагиали характерны и другие «полуглубоководные» рыбы, обитающие также в верхней мезопелагиали. В их числе следует упомянуть прежде всего представителей подотряда вогмеровидных (Trachipteroidei), включающего 9 родов (около 15 видов), в том числе сельдяных королей (Regalecus glesne) (рис. 12) и вогмеров (род Trachipterus). Для этих рыб характерен аллометрический рост, т. е. большая онтогенетическая изменчивость формы тела и относительных размеров его частей, и пот этому мальки, молодь и взрослые особи некоторых видов долго носили разные научные названия. Вогмеровидныв рыбы имеют удлиненное и плоское — ремнеобразное — тело, сужающееся к хвосту, и отличаются большой длиной (не менее 1м). Сельдяной король достигает в Северной Атлантике даже длины 5,5 м при массе порядка 250 кг (известны и случаи поимки рыб длиной 7—8 м), а лососевый король (Trachipterus altivelis) из северо-восточной части Тихого океана в возрасте 7—8 лет имеет длину 1,6 м и весит 4 кг. (При сходной в общих чертах форме тела соотношения этих цифр кажутся довольно странными, но они вполне соответствуют представлениям о пропорциональности массы рыб кубу их длины.)
Вогмеровидные — хищники, питающиеся разнообразными рыбами и беспозвоночными. Иногда они встречаются в косяках массовых промысловых видов, в частности сельди и дальневосточных лососей, которых, как считали в давние времена, ведут за собой (отсюда и название «короли», но на деле их связи с «подданными» имеют чисто гастрономический характер). Эти рыбы плавают обычно головой кверху, в положении, близком к вертикальному. При этом они поддерживают от опускания собственное тело, плотность которого больше, чем плотность воды, и поступательно перемещаются с небольшой скоростью за счет одних только ундулирующих движений длинного спинного плавника. Если нужно плыть быстрее, короли передвигаются, волнообразно изгибая свое длинное тело. У всех представителей подотряда тело окрашено в серебристо-белый цвет, на туловище, как правило, разбросаны короткие темные полосы и пятна, плавники у многих ярко-красные. Виды семейства лофотовых (Lophotidae) имеют своеобразный чернильный мешок, открывающийся в клоаку, и способны создавать маскирующую «дымовую завесу» подобно кальмарам и каракатицам.
Вероятно, встречи моряков с гигантскими сельдяными королями, плавающими у поверхности, были причиной многих историй о «морском змее», который в некоторых рассказах описывается как чудовище, имеющее лошадиную голову с развевающейся огненно-рыжей гривой. За такую гриву, видимо, принимали длинные лучи спинного плавника, образующие головной султан этой рыбы.
Еще одно нижнеэпипелагическое (или эпимезопелагическое) семейство — морские лещи (Bramidae) — также приурочено преимущественно к субтропическим и тропическим водам, хотя отдельные виды обитают и в сравнительно холодной зоне. Атлантический лещ (Вгата Ьгата), например, населяет Средиземное море и северную Атлантику и встречается при температуре 10—15°. В южной части области распространения он живет на глубине, а у его северных границ обычен и в поверхностном слое. Эта хищная рыба достигает длины 65—70 см и массы 6 кг и имеет, как и близкородственный северотихоокеанский лещ; (В. japonicus), некоторое промысловое значение. Среди более редких видов можно отметить серебряного морского леща (Pterycombus brama), рыбу с блестящим серебристым телом, желтыми грудными и иссиня-черными спинными а анальными плавниками; при этом оба вертикальных плавника сильно увеличены и имеют парусовидпую форму.
Рассматривая нижнеэпипелагических рыб, нельзя ни упомянуть еще один интересный вид — луваря (Luvarua imperialis), которого выделяют в особое семейство Luvaridae. Это семейство длительное время относили к скумбрие видным рыбам, но недавно было показано, что оно наиболее родственно, как это ни удивительно, прибрежным рыбам-хирургам (семейство Acanthuridae), обитающим главным образом на коралловых рифах. Луварь имеет удлиненное плоское тело и лобастую голову с крохотным ротиком. Он достигает 2 м в длину и весит до 150 кг, очень плодовит (47 млн ооцитов), его личиночные и мальковые стадии очень мало похожи на взрослых луварей. Основной пищей являются медузы и другие студенистые формы планктона, т. е. низкокалорийные объекты, и в связи с этим, как считают Д. Фитч и Р. Лавенберг, он обладает очень длинным кишечником (у метровой рыбы он составлял 92% длины тела). Луварь распространен во всех океанах, но всюду редок, и большинство поимок приходится на субтропическую зону.
Совершенно особую и очень своеобразную группировку в составе эпипелагического ихтиоцена представляют рыбы, ассоциированные с плавучими водорослями саргассами, которые обычно выносятся в океан с прибрежных мелководий, но кое-где (например, в Саргассовом море) существуют в замкнутых круговоротах без всякой связи с берегами. Помимо молоди некоторых океанических рыб (летучки, корифены) и рыб прибрежного происхождения, среди саргассов постоянно обитают два высокоспециализированных вида — морской клоун Histrio histrio (длина до 18 см) и пелагическая рыба-игла Syngnathus pelagicus (длина до 10 см). Внешний вид и окраска этих рыб, никогда не выходящих за пределы водорослевых кустов и скоплений (рис. 13), отлично маскируют их в саргассах, и их нелегко, обнаружить в этом укрытии, даже точно зная, что они скрываются там. Рыба-клоун привлекает добычу специальной приманкой своей «удочки» (первого луча спинного плавника) и заглатывает ее, быстро увеличивая объем ротовой полости при закрытом рте и открывая пасть, когда жертва оказывается прямо перед ней, — весь этот процесс укладывается в тысячные доли секунды.
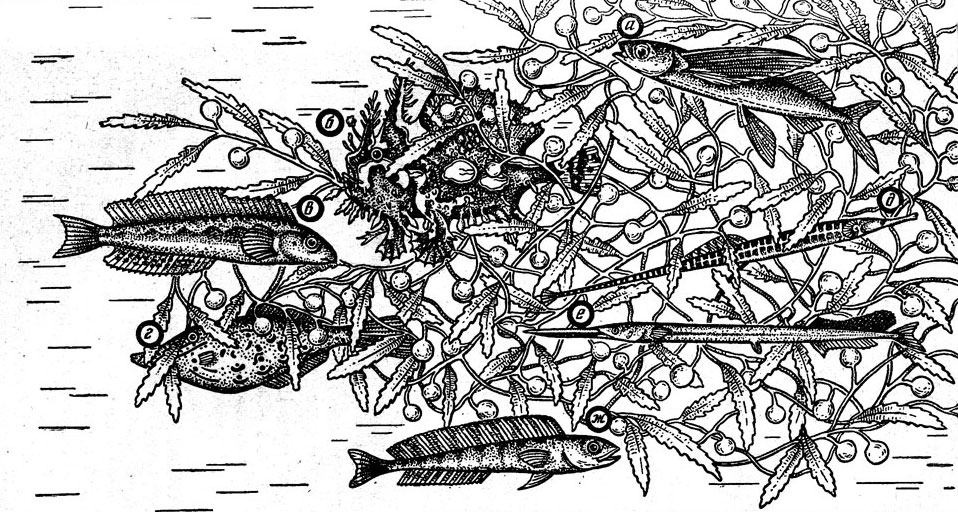
Рис. 13. Рыбы, ассоциированные с плавучими водорослями.
Почти все рыбы, перечисленные выше, живут в эпипелагиали постоянно — со времени появления на свет до конца своего существования. Таких рыб предложено называть голоэпипелагическими и противопоставлять их видам меропипелагическим, в обязательном порядке проводящим в верхних горизонтах открытого океана лишь определенную часть своего жизненного цикла, и ксеноэпипелагическим, представляющим собой более или менее случайный элемент в поверхностном слое (см. рис. 14) ( «Голос» по-гречески - весь, целый, «мерос» - часть, доля, «ксенос» - чуждый.).

Рис. 14. Основные группировки рыб эпипелагиали.
В 1964 г. директор Института океанологии В. Г. Корт поставил перед сотрудниками задачу обобщить в коллективной монографии все сведения о природе Тихого океана, накопленные к тому времени. Мне было поручено свести воедино то, что было известно об эпипелагических рыбах, которые тогда изучались советскими экспедициями (напомню, что в дальнем океане в те времена фактически работал один «Витязь») в очень узких рамках — в открытых водах проводились только световые станции и сборы ихтиопланктона. В отношении наиболее массовых и соответственно наиболее интересных для промысла видов - тунцов, марлинов и других - пришлось использовать только литературные данные. Эта компилятивная работа приносила мне мало удовлетворения (особенно на ее начальном этапе), но именно она заложила основы моих представлений о сложной структуре «композитного» океанического ихтиоцена. После выхода в свет многотомника «Тихий океан», который получил высокую оценку (группе его авторов была присуждена Государственная премия СССР), я значительно расширил свои главы, дополнил их сведениями по другим океанам и. подготовил монографию «Ихтиофауна океанской эпипелагиали», которая стала моей докторской диссертацией. Ее основные положения вошли в эту книгу но, конечно, в значительно видоизмененном изложении: за 20 лет, прошедших со времени ее опубликования, изучение рыб открытого океана очень сильно продвинулось вперед, да и сам я стал по-иному смотреть на многие вещи. Сейчас, например, я никак не дал бы той книжке ее старого названия (ведь в ней речь идет не столько о «фауне», сколько о «таксоцене»), отказался бы от ненужного, как вижу теперь, придумывания новых, порой труднопроизносимых терминов для обозначения отдельных экологических группировок рыб, внес бы многочисленные коррективы в биологические характеристики отдельных видов. Впрочем, «что написано пером, того не вырубишь топором»...
Итак, по числу видов временные обитатели эпипелагиали не уступают голоэпипелагической группе. Некоторые из них достигают в отдельных районах океана очень высокой численности и биомассы.
Все мероэпипелагические рыбы используют эпипелагиаль посредством замкнутых миграционных циклов. Именно этой особенностью они достаточно четко отличаются от ксеноэпипелагических рыб, не совершающих регулярных и целенаправленных перемещений между эпипелагиалыо ц другими биотопами. Большинство ксеноэпипелагических видов нормально обитают в неритической зоне, но, попав за ее пределы, они способны долго существовать (по-видимому, не размножаясь) и в практически неограниченном удалении от берегов. В качестве примера можно упомянуть хотя бы представителей трех семейств отряда сарганообразных — парусноперую летучую рыбу Parexocoetus brachypterus (семейство Exocoetidae), длинноносого полурыла Euleptoramphus viridis (Hemiramphidae), лентовидного саргана Ablennes hians (Belonidae), которые, являясь по своему существу неритическими, могут быть встречены в любой точке тропического океана. В северной части Тихого океана, в Баренцевом и Черном морях на большом расстоянии от берега можно наблюдать трехиглую колюшку, или колючку (Gasterosteus aculeatus),— обычного обитателя наших рек, озер и прибрежных морских вод. К этой же группе следует отнести и тех прибрежных альгофильных рыб, которые встречаются в тропической эпипелагиали вместе с плавающими водорослями или другими дрейфующими предметами берегового происхождения, например крокодилового саргана (Tylosurus crocodilus), пятнистого спинорога (Canthidermis maculatus), абудефдуфа (Abudef-duf saxatilis), терапона (Therapon theraps), чаба (Kypho-sus cinerascens). Как показал Б. И. Федоряко, эти и некоторые другие рыбы, будучи вынесенными в открытые воды еще на мальковой стадии, прекрасно чувствуют себя и вдали от берегов, там, где преобладающее большинство прибрежных рыб гибнет очень быстро. Способность к долгому выживанию в эпипелагиали, несомненно, содействует широкому расселению ксеноэпипелагических видов, для которых не существует барьеров в виде открытоводных пространств любого протяжения. Многие из них имеют поэтому циркумглобальные ареалы.
Временные обитатели эпипелагиали проникают в нее либо снизу — из глубинных вод, либо «сбоку» — из неритической зоны и с прибрежных мелководий. При этом мероэпипелагические глубоководные рыбы появляются в эпипелагиали лишь ночью (здесь речь идет только о взрослых особях), когда опи поднимаются вверх в ходе регулярных суточных миграций, а перед рассветом вновь уходят на глубины. К этой никтоэпипелагической группе принадлежат уже упоминавшиеся мельком приповерхностные светящиеся анчоусы и змеиные макрели, а также некоторые карликовые акулы и отдельные представители типичных мезопелагических семейств, о которых пойдет речь в следующей главе. Следует отметить, однако, что в отсутствие резкого температурного градиента между приповерхностными и глубинными водами (это характерно для внетропической зоны) обособленность эпи- и мезопелагической группировок рыб выражена значительно слабее, чем в районах с хорошо развитым термоклином, представляющим собой препятствие для вертикальных мигрантов.
Мероэпипелагические рыбы прибрежного происхождения, которые встречаются в открытых водах только в период нагула, по размножаются у берегов или даже в пресных водах, составляют очень характерный элемент населения эпипелагиали. Они нередко образуют значительную биомассу, в некоторых районах соизмеримую с биомассой постоянных обитателей эпипелагиали или даже превышающую ее. Типичными представителями этой группировки являются, в частности, настоящие (род Salmo) и тихоокеанские (Oncorhinchus) лососи. Представители последнего рода — кета, горбуша, нерка, кижуч, чавыча, сима — эндемичны для бассейна северной части Тихого океана и в период нагула являются наиболее массовыми рыбами поверхностного слоя на огромной акватории — от Берингова моря до 40° с. ш. Тихоокеанские лососи образуют многочисленные репродуктивные стада, нерестящиеся в реках дальневосточного побережья Азии (на юг до п-ова Корея и Японии; жилая форма нерки есть даже на о-ве Тайвань) и Северной Америки (до Калифорнии). Ареалы таких стад в открытом океане в значительной степени перекрываются, и между 165° в. д. и 155° з. д. даже азиатские и американские лососи встречаются совместно. Длительность морского периода жизни у разных видов неодинакова — от 1,5 лет у горбуши до 4—7 лет у чавычи, наиболее крупного вида, иногда достигающего 50 кг. Они питаются среднеразмерными и крупными планктонными животными — ракообразными, крылоногими моллюсками, молодью кальмаров и рыб. Границы морских ареалов тихоокеанских лососей претерпевают значительные сезонные изменения, однако даже в зимний период область их распространения не простирается за пределы субарктических вод.
Атлантическо-скандинавская (так называемая океаническая) сельдь (Clupea harengus) размножается в прибрежных водах Норвегии и Исландии, откладывая клейкую икру на водоросли, а в период нагула ведет пелагический образ жизни вдали от берега. В годы хорошего состояния запасов она совершает довольно протяженные миграции в открытую часть Северной Атлантики и достигает Баренцева моря, Шпицбергена, Гренландии (сейчас численность этого вида находится в состоянии депрессии). Примерно в этом же районе в современный период происходит нагул северной путассу (Micromesistius poutassou) — стайной рыбы семейства тресковых (Gadidae), размножение которой также ограничено склонами мелководий в неритической зоне.
Самая большая из хищных рыб Южного океана — антарктический клыкач (Dissostichus mawsoni) размножается у берегов Антарктиды, где держится и подрастающая молодь длиной до 1 м, тогда как более крупные (до 1,8 м) половозрелые рыбы, как было показано В. Л. Юховым, совершают долговременные миграции в пелагиаль, удаляясь па расстояние до 900 миль от материка. Такое распространение клыкачей было установлено по их нахождениям в желудках кашалотов, питающихся преимущественно на глубине 100—1000 м, что дало основание относить эту рыбу к временным обитателям мезопелагиали. Может быть, это действительно так, но, учитывая недостаточность имеющихся данных, нет оснований признавать строго обоснованным тезис об отсутствии клыкача в поверхностных горизонтах.
Другая группа мероэпипелагических видов представлена в эпипелагиали только ранними стадиями развития (вплоть до мальков). Для всех этих видов, живущих во взрослом состоянии в неритической пелагиали или у дна, прохождение эпипелагической стадии в онтогенезе является необходимой и неизбежной фазой жизненного цикла. Будучи хорошо адаптированными к весьма длительному пребыванию в толще воды, такие мальки нередко встречаются в огромном удалении от берегов и сохраняют шансы на благополучное возвращение в прибрежную зону, хотя значительная их часть, конечно, обречена на неизбежную гибель в открытых водах.
Разделение эпипелагических рыб открытого океана между голо-, меро- и ксеноэпипеелагической группировками, как правило, достаточно очевидно, хотя не абсолютно — как обычно, желание разложить природное разнообразие по полочкам вступает в противоречие с практикой жизни. Так, многие виды крупных акул (в частности, кархародон и тигровая), большая барракуда (Sphyrae-na barracuda), ауксиды, полосатый и желтоперый тунцы, ваху, большая корифена, по-видимому, в той или иной степени обычны и в неритической зоне, и в открытых водах (часто их называют нерито-океаническими). Не во всех случаях при современной степени экологической изученности можно четко разграничить мероэпипелагических и ксеноэпипелагических рыб.
Полный перечень отрядов, семейств и родов, представленных в эпипелагиали открытого океана, приводится ниже. Для каждого из родов указаны в скобках число эпипелагических видов и их принадлежность к числу постоянных (Г), временных (М) и случайных (К) обитателей биотопа (виды, представленные в эпипелагиали только молодью, не учтены).
Систематический состав эпипелагического ихтиоцена
Отряд миногообразные (Petromyzontiformes). Сем. миноговые (Petromyzontidae): Geotria (1 К).
Отряд катранообразные (Squaliformes). Сем. катрановые (Squalidae): Euprotomicrus (1 М); к числу мигрирующих ночью из мезопелагиали или живущих на стыке мезо- и эпипелагиали относятся еще 9 видов из 6 родов — Isistius, Squaliolus, Euprotomicroides и др. Отряд воббегонгообразные (Orectolobiformes). Сем. китовые акулы (Rhiniodontidae): Rhmiodon (1 Г).
Отряд ламнообразные (Lamniformes). Сем. ложнопесчаные акулы (Pseudocarchariidae): Pseudocarcharias (1 Г); лисьи акулы (Alopiidae): Alopias (1 Г и 2 К); большеротые акулы (Megachasmidae): Megachasma (1 К); гигантские акулы (Cetorhinidae): Cetorhinus (1 К); сельдевые акулы (Lamnidae): Lamna (2 Г); Isurus (2 Г), Carcharodon (1 К). Отряд кархаринообразные (Carcharhiniformes). Сем. серые акулы (Carcharhinidae): Carcharhinus (2 Г и 5-6 К), Prionace (1 Г), Galoocerdo (IK); молотоголовые акулы (Sphyrnidae): Sphyrna (3 К). Отряд хвостоколообразные (Myliobatiformes). Сем. скаты-хвостоколы (D аsyatidae): Dasyatis (1 Г); мантовые скаты (Моbulidaa): Mania (1 К).
Отряд гнюсообразиые (Torpediniformes). Сем. гнюсовые (Torpedinidae): Torpedo (2-3 К).
Отряд угреобразные (Anguilliformes): Сем. змеехвостые угри (Ophichthyidae): Benthenchelys (1 Г). Отряд сельдеобразные (Clupeiformes). Сем. сельдевые (Clupeidae): Clupea (1 M), Sardinops (1 К); анчоусовые (Еngraulidae); Stolephorus (1 Г), Engraulis (1 К). Отряд лососеобразиые (Salmoniformes). Сем. лососевые (Salmonidae): Oncorhynchus (5 M), Salmo (3 M). Отряд стомиеобразные (Stomiiformes). Сем. фотихтовые (Photichthyidae): Vinciguerria (2 M); астронестовые (Astronesthidae): Astronesthes (1-2 M). Отряд аулопообрэзиые (Aulopiformes). Сем. алепиэавровые (Alepisauridae): Alepisaurus (2 К); кинжалозубовые (Anotopteridae): Anotopterus (1 К), веретенниковые (Paralepididae): Paralepis и др. (3—4 К). Отряд миктофообразные (Myctophi formes). Сем. миктофовые (Myctophidae): Electrona (2 M), Benthosema (1 Г), Hygophum (2 M), Myctophum (13 M), Symbolophorus (около 10 M), Loweina (4 M), Tarletonbeania (1 M), Gonichthys (4 M), Centrobranchus (4 M). Отряд трескообразиые (Gadiformes). Сем. тресковые (Gadidae): Micromesistius (2 МЭн).
Отряд удильщикообразные (Lophiiformes), Сем. рыбы-клоуны (Antennariidae): Histrio (1 Г). Отряд capганообразные (Beloniforraes). Сем. летучие рыбы (Ехосоеtidae): Parexocoetus (1 К), Exocoetus (3 Г), Prognichthys (4 Г и 1 К), Cypselurus (5-6 К), Cheilopogon (5 Г и 10-12 К), Hirundichthys (3 Г и 3 КЭи);полурыловые (Hemiramphidae): Oxyporhamphus (2 Г), Euleptorhamphus (2 К); сарга новые (Belonidae): Platybelone (1 К), Ablennes (1 К), Tylosurus (2 К); макрелещуковые (Scomberesocidae): Scomberesox (1 Г), Cololabis (1 Г), Nanichthys (1 Г), Elassichthys (1 Г). Отряд опахообразные (Lampridiformes). Сем. опаховые (Lampridae): Lampris (2 Г); лофотовые (Lophotidae): Lophotus (1 Г), Eumecichthys (1 Г); радицефаловые (Radiicephalidae): Radiicephalus (1 Г); вогмеровые (Тrасhiptоridae): Trachipterus (5Г), Desmodema (2 Г), Zu (2Г); ремень-рыбы (Regalecidae): Regalecus (1 Г), Agrostichthys (1 Г).
Отряд колюшкообразные (Gasterosteiformes). Сем. колюшковые (Gasterosteidae): Gasterosteus (1 К); рыбы-иглы (Syngnathidae): Syngnathus (1 Г). Отряд екорпенообразмме (Scorpaeniformes). Сем. пипагоровые (Сус1орtегidае): Pelagocyclus (1 Г), Cyclopterus (1 К). Отряд окунеобразные (Pcrciformes). Сем. ставридовые (Carangidae): Elagatis (1 Г), Naucratus (1 Г), Decapterus (2-3 К), Trachurus (2-3 К); рыбы-прилипалы (Есhеnеidае): Phtheiricbthys (1 Г), Remoга (5 Г), Remorina (1 Г); корифеновые (Соrурhaenidae): Coryphaena (2 Г); морские лещи (В га mid а с): Таrасtes (2 Г), Brama (6 Г), Taractichthys (2 Г), Pterycombus (2 Г), Ptoraclis (3 Г); нототениевые (Nototheniidae): Dissostichus (2 M), Pleuragrainma (1 К), Paranotothenia (1 К); и костеевые (Icosteidae): Icosteus (1 Г); лунарепые (Luvari-
Jae): Luvarus (1 Г); барракудовые (Sрhуrnidae): Sphyrna (1 К); темнило вые (Gеmруlidae): Lepidocybium (1 К). Ruvettus (1 К), Gempylus (I M); скумбриевые (Sсоmbr idae): Gasterochisma (1 Г), Scomber (2 К), Acanthocybium (1 ГЭв), Allolhunnus (1 Г), Auxis (2 Г), Euthynnus (2 КЭп), Katsuwonus (1 Г), Thunnus (7 Г); меч-рыбы (Xiphiidae): Xiphias (1 Г); парусников ые (Istiophoridae): Istiophorus (1 К), Makaira (3 Г), Tetrapterus (5 Г); центролофовые (Сentrolophidae): Centrolophus (1 Г), Icichthys (1 Г), Pseudoicichthys (1 Г), Tubbia (1 Г), Schedophilus (5 Г); амарсиповыэ (Amar sipida e): Amarsipus (1 Г); номе ев ые (Nоmоidae): Cubiceps (9 Г), Nomeus (1 Г), Psenes (6 Г); алетовые (Tetragonuridae): Tetragonurus (3 Г). Отряд скалозубообразныс (Tetraodontif ormes). Сем. спи пороговые (Balistidae): Canthidermis (1 Г), Balistes (1 К), Xanthichthys (2 К); ска л о зубовые (Tetraodontidae): Lagocephalus (1 Г); ежи-рыбы (Diodontidae): Diodon (1 Г + 2 К); рыбы-луны (Мо1idae): Ranzania (1 Г), Masturus (1 Г), Mola (2 Г).
Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пе-лагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодной Атлантике, например, в последние годы резко увеличилась численность серого спиноорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus (рис. 15). Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Fix выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
Рис, 15. Максимально расширенные ареалы псевдонеритических рыб — атлантической сельди (1), сардины иваси и дальневосточной скумбрии (2), калифорнийской (3) и перуано-чилийской ставриды (4) и возможные пути расселения южной путассу (3).
В двух последних случаях вид может рассматриваться как временно или постоянно обосновавшийся в том или ином конкретном участке открытого океана. Примеры, иллюстрирующие сказанное, ограничиваются, к сожалению, данными за самые последние десятилетия, так как имеющиеся ряды наблюдений очень коротки. Большая часть этих примеров относится к Северной Пацифике.
Известно, что в период высокой численности японского анчоуса и японской скумбрии в 50-е годы оба вида были весьма обычными и в открытых водах северо-западной части Тихого океана, в которых предполагалось даже существование их особых «стад» — в действительности, по всей вероятности, псевдопопуляций. В этом же районе в период высокой численности сардины иваси (Sardinops sagax melanosticta) в 30-е годы ее ареал расширялся вплоть до южной части Берингова моря (наблюдения в открытом океане отсутствовали), а современный подъем численности, начавшийся в 70-е годы (за первые 5 лет роста запас возрос не менее чем вдвое), привел к расширению области распространения в восточном направлении вплоть до центральной части океана, где, судя по некоторым сведениям, также образовывались временные псевдопопуляции. У японской скумбрии, как показано В. А. Беляевым, усиление воспроизводства на нерестилищах в прибрежной зоне Японии в середине 70-х годов привело к заселению океанических участков и формированию в районе Северо-Западного подводного хребта (гора Кинмей и прилегающие воды) зависимой популяции псевдонеритического типа: в 1978—1981 гг. скумбрия наблюдалась там на всех фазах жизненного цикла (икра, личинки, мальки, сеголетки).
Интересный материал для размышлений представляют сведения по биологии и распространению двух восточнотихоокеанских видов ставрид из группы Trachurus picturatus. Размножение калифорнийской ставриды (Т. symmetricus simmetricus) было отмечено по нахождениям икры и личинок в зоне 40-х параллелей северного полушария в 1955г. в удалении до 1100 миль, а в 1972 г.— до 600 миль от берегов Северной Америки. Аналогичным образом нерест перуанско-чилийской ставри-ды (Т. s. murphyi) в 40-х широтах южного полушария отмечен в 1985 г. С. А. Евсеенко на огромном расстоянии (около 3000 миль) от побережья Чили. Никаких свидетельств в пользу результативности такого нереста, впрочем, не существует, так как в этих океанических участках — у крайних границ своего распространения на запад — оба вида представлены, судя по имеющимся данным, только очень крупными особями длиной 45—60 см (там нет ни мальков, ни сеголетков, ни созревающих рыб, а созревает ставрида в возрасте 2—3 лет). Таким образом, ати факты могут трактоваться лишь как доказательства существования временных псевдопопуляций (маловероятным представляется даже обособление зависимых популяций), которые не только существуют за счет постоянного пополнения из прибрежного запаса, но и являются фактически его излишком. Выселение перуанской ставриды в океан вполне объяснимо ростом ее численности в неритической зоне. Так, согласно рыбопромысловой статистике ФАО (Продовольственная и сельскохозяйственная организация ООН), уловы этого вида в прибрежных водах Перу и Чили (они, видимо, отражают и истинную численность ставриды) увеличились со 100 тыс. т в 1970 г. и 300—400 тыс. т в 1974—1976 гг. до 1100—1300 тыс. т в 1978—1980 гг., а в открытом океане эта рыба была обнаружена в 1978 г.
Какой же биологический смысл могут иметь безвозвратные уходы прибрежно-пелагических рыб от родных берегов? Какую выгоду получает неритическая популяция, посылая на гибель в открытом океане большие группы «лишних» особей? Такая постановка вопросов представляется не совсем корректной (ибо отнюдь не все в живой природе имеет прямое приспособительное значение), но ответ на них все-таки существует: в результате таких выселений появляется потенциальная возможность расширить видовой ареал, включив в него новые неритические участки, лежащие за «барьером» открытоводных просторов.
То, что я написал здесь, еще два-три года назад показалось бы очень спорным большинству моих коллег-ихтиологов. Обнаружение ставриды в юго-восточной части Тихого океана далеко за пределами перитической зоны, в которой ей, казалось бы, полагалось пребывать, послужило основой для рассуждения об особых океанических популяциях этой рыбы, совершенно самостоятельных и не имеющих никакого отношения к прибрежным группировкам своего вида. Высказывались даже предположения о существовании своего рода «ставридного пояса», сплошным кольцом опоясывающего земной шар в районе «ревущих сороковых» широт южного полушария, и специально посланные суда искали там (да и в других участках открытого океана) «большую рыбу» — обособленные популяции массовых прибрежных видов. Мои представления (а я неоднократно излагал их в докладах на разных ученых собраниях и просто в разговорах с товарищами по профессии) не встречали тогда особой поддержки, хотя не удостаивались и критики. Сейчас, однако, оппонентов этим взглядам практически не осталось, и обо всей этой истории можно было бы и не упоминать, если б не одно важное обстоятельство, на которое хочется обратить специальное внимание: ведь если все так, как здесь написано, то рассчитывать на неограниченно долгое существование той же перуанско-чилийской ставриды в дальнем океане не приходится. Цикличность процесса неизбежно предполагает грядущий спад численности и даже почти полное исчезновение этой рыбы из открытых для международного рыболовства вод. Правда, в те же сроки в каких-то иных участках Мирового океана могут появиться псевдопопуляции других видов. Прогнозировать ход этих пульсационных процессов — одна из важнейших, на мой взгляд, задач прикладной рыбохозяйственной науки.
В целом возникновение новых самостоятельных популяций неритических видов в результате выселения в открытый океан представляется довольно маловероятным событием, но именно этим способом, по-видимому, возникли «баночные» популяции некоторых рыб, в частности ставриды (Т. picturatus) и рыбы-сабли (Lepidopus caudatus), на подводных поднятиях северо-восточной Атлантики в районе Азорских островов. Современный прерванный ареал южной путассу (Micromesistius australis), обитающей сейчас в неритических водах Патагонии и Новой Зеландии (см. рис. 15), также может быть объяснен расселением из Атлантического океана в зоне южных 50-х широт, что могло сопровождаться образованием и исчезновением на пути следования зависимых популяций такого типа, как существующая ныне, по данным В. П. Шпака, в районе подводных гор Южно-Тихоокеанского поднятия на 130° з. д.
Проникновение прибрежно-пелагических видов в открытые воды не может быть причиной того, чтобы подвергать сомнению реальность обособленности неритнческих и океанических ихтиоценов пелагиали. Это явление (как и обратный процесс) наглядно демонстрирует, однако, динамичность биологической структуры эпипелагиали, подверженной сильному влиянию климатических факторов.
Как видно из всего сказанного, эпипелагические рыбы довольно разнообразны по экологии и образу жизни. Их изучение в этих аспектах еще далеко от завершения, но уже сейчас довольно отчетливо просматриваются некоторые общие закономерности, дающие представление об основных экологических характеристиках ихтиоцена и взаимоотношениях слагающих его видов. Эти закономерности представляют немаловажное значение для исследования биологической продуктивности океана, в частности, для изучения проблем, связанных с перераспределением органического вещества.
Размножение. Вполне естественно, что голоэпипелагические рыбы, живущие в полном отрыве от твердого субстрата, имеют пелагические икринки, проходящие развитие в толще воды, но, как уже говорилось, из этого правила есть исключения. Некоторые сарганообразные рыбы, а именно сайры и «четырехкрылые» летучки из родов Cheilopogon и Hirundichthys, подобно многим неритическим представителям своего отряда, откладывают клейкую икру на твердый субстрат, плавающий у поверхности. Икра других сарганообразных (полурыла Охурог-hamphus, «двукрылых» летучих рыб, макрелещуки) развивается в планктоне, однако общей особенностью эмбрионального развития всей этой группы служит его большая продолжительность (одна-две недели). Большая длительность эмбриогенеза отмечена также у сельдяных королей и их родичей. У остальных эпипелагических рыб развитие икринок заканчивается в очень короткий срок, особенно в тропических водах (у большеглазого тунца, например, оно продолжается при температуре воды 28— 29° всего 21 час).
Плодовитость эпипелагических рыб, как правило, очень велика. Только у миктофид (их длина, напомним, редко превышает 10 см), летучих рыб и макрелещуковых она исчисляется тысячами икринок, тогда как крупные рыбы поверхностного слоя продуцируют икру миллионами и десятками миллионов. В качестве примера можно указать, что змеиная макрель и ауксида выметывают до 1 млн икринок, полосатый тунец — до 2 млн, большеглазый тунец и ваху — по 6 млн, синий тунец — 10 млн, меч-рыба — 16 млн, сельдяной король — не менее 20 млн, лу-варь — 47 млн, большой синий марлин — почти 100 млн, рыба-луна — даже 300 млн икринок. Для преобладающего большинства эпипелагических рыб характерна порционность икрометания, приводящая к большой растянутости сроков нереста. В тропических водах в связи с относительным постоянством условий среды размножение может идти даже круглогодично (например, у летучих рыб и тунцов). Высокая плодовитость и ускоренное прохождение ранних стадий индивидуального развития направлены у костистых рыб на компенсацию огромной смертности икры и личинок при полном отсутствии родительской заботы о потомстве. В отличие от них все эпипелагические акулы и скаты (имеется в виду пелагический хвостокол — Dasyatis violacea, принадлежащий к числу постоянных обитателей открытого океана) относятся к яйцеживородящим или живородящим видам и приносят немного (от 2 до 100) крупных детенышей, почти не имеющих врагов.
Рост и возраст. Для эпипелагических рыб очень характерен быстрый рост и, как его следствие, раннее наступление половой зрелости. Аквариальные наблюдения показали, например, что малек сайры длиной 6 см через полгода достиг 20 см, а малек Psenes maculatus (семейство Nomeidae) длиной 7,5 см за три месяца вырос до 28 см (при массе 775 г). Сейчас известно, что самые мелкие из рыб поверхностного слоя —летучки рода Ехоcoetus и сайра — достигают половозрелости уже к концу первого года жизни, а предельная продолжительность их существования не превышает 1,5—2 лет. Быстрым ростом в течение морского периода жизни отличаются лососи. Горбуша, например, возвращающаяся на нерест в двухлетнем возрасте, за полтора года, проведенных в эпипелагиали, увеличивает длину от 3—5 см в момент ската из рек до 42—53 см (при массе 0,9—1,9 кг). Змеиная макрель живет 5 лет, морской лещ — около 8 лет. К особенно быстро растущим рыбам относятся тунцы, меч-рыба и марлины, годовой прирост которых составляет 20— 40 см. Желтоперый тунец впервые мечет икру в возрасте 2—3 лет, большеглазый — в 3 года, длинноперый — в 4—5 лет, меч-рыба — в 5—6 лет, а продолжительность их жизни не превышает 8—10 лет. К числу долгожителей эпипелагиали принадлежат, по-видимому, акулы: наибольший возраст синей акулы достигает по меньшей мере 20 лет, а сельдевой, возможно, и 30 лет.
Питание. В трофическом отношении все эпипелагические рыбы разделяются на две большие группы — мирных планктофагов и хищников (преимущественно ихтиофагов). При этом рыб, питающихся исключительно мелким планктоном, в эпипелагиали открытого океана сравнительно немного. Наибольшей численности среди них достигают голоэпипелагические макрелещуковые и летучие рыбы, а также мероэпипелагические светящиеся анчоусы в выходящие в открытые воды рыбы прибрежного происхождения — лососи, сельдевые, ставриды, скумбрии, тресковые, возможно, нототениевые. К этой группе относятся, естественно, и все планктонные личинки и мальки рыб, которые могут использовать в пищу даже фитопланктон. Почти у всех эпипелагических планктофагов пищевые спектры очень широки, а локальные и сезонные изменения состава их пищи определяются в значительной степени только наличием доступных кормовых объектов. Редкий пример избирательного питания показывают, однако, строматеевидные рыбы (номеи, кубоглавы и др.) и луварь, основным объектом потребления которых служат мягкотелые беспозвоночные — оболочники (сальны и пиросомы) и кишечнополостные (медузы, гребневики, сифонофоры), на 99% состоящие из воды. Для измельчения этого корма у строматеевидных имеется даже специальный орган — снабженные внутри острыми зубами мешковидные выросты с обеих сторон глотки позади жаберных дуг. Возможно, впрочем, непосредственным источником питания рыб служит в этих случаях непереваренная пища их жертв.
Большинство рыб эпипелагиали относится к числу хищников, пищей которых являются рыбы, кальмары и крупные ракообразные. Размеры добычи в значительной степени определяются величиной самого хищника, а состав пищи меняется в зависимости от специфики участка или горизонта, на котором происходит питание. Так, обычной пищей корифен в открытых водах служат летучие рыбы, приповерхностные миктофиды, пелагические осьминоги-аргонавты и кальмары, однако в водах, богатых водорослевым плавником, они поедают преимущественно ставридовых, спинорогов и других рыб, ассоциированных с плавучим субстратом.
Особенно внимательному изучению подвергалось в последние годы питание тунцов, меч-рыб и копьерылов. Полученные данные демонстрируют прежде всего огромную пищевую пластичность этих рыб, потребляющих самые разнообразные кормовые объекты — преимущественно рыб и головоногих моллюсков. В списках компонентов питания полосатого тунца, например, значится более 180 видов жертв, большеглазого тунца — 152 вида, желтоперого тунца — 243 вида, причем основную пищу двух последних видов составляют объекты, обитающие на границе эпи- и мезопелагиали. Как показано О. Д. Бородулиной, «осредненные» рационы этих хищников очень сходны, но желтоперый тунец поедает больше номеевых, летучих рыб и других тунцов, а в желудках большеглазого чаще встречаются более глубоководные виды, и это снижает остроту их конкурентных отношений. Спектры питания тунцов заметно меняются в зависимости от их размеров, что также связано, помимо прочих причин, с различной глубиной охоты особей разных возрастных групп. Так, мелкие особи желтоперого тунца поедают главным образом ракообразных, среднеразмерные — рыб, кальмаров и ракообразных, крупные — рыб и кальмаров. В пище тунцов и других крупных хищников отсутствуют, однако, интерзональные мигранты, поднимающиеся ночью с глубин в те горизонты, где откармливаются тунцы, или пересекающие эти горизонты (это можно объяснить только питанием хищников в светлое время суток).
Таким образом, для эпипелагических рыб типичны слабо выраженная избирательность питания и как ее следствие наличие широких пищевых спектров. Большинство епипелагических рыб питается практически любым доступным кормом, подходящим по размерам и имеющимся в данном месте в достаточном количестве. В схеме трофической структуры океанской пелагиали (рис. 16) рыбы занимают несколько верхних уровней — до пяти в тропических водах, характеризующихся наибольшей сложностью пищевых взаимоотношений.
Распространение. Хорошо известно, что границы ареалов эпипелагических рыб, принадлежащих к нектону (т. е. способных к активным горизонтальным перемещениям) , определяются главным образом температурой воды. Связь распространения с температурными условиями показана для многих рыб — акул, лососей, летучих рыб, тунцов, марлинов и др. Двукрылая летучая рыба Exocoetus volitans, например, встречается при температуре 22—29°, будучи обычной при 24—28°. В результате область распространения этого вида в наиболее теплой западно-экваториальной части Тихого океана была прервана осенью 1961 г. примерно на 15° по широте (т. е. на 900 морских миль), а в центральной части океана, где в области экватора температура несколько понижена благодаря поднятию глубинных вод, не имела такого разрыва. Минимальные значения температуры, отмеченные для разных видов массовых скумбриевидных рыб открытого океана в период нагула, варьируют от 10—12° у длинноперого тунца и меч-рыбы до 18—20° у желтоперого тунца и синего марлина. Температурный диапазон встречаемости в океане лососей рода Oncorhynchus полностью укладывается в пределы 2,3—12°, причем наиболее холодолюбивым видом является нерка, обычная при 2,3—9°, а наиболее теплолюбивым — кижуч, предпочитающий температуру 7—12°, Подобные примеры, количество которых может быть многократно увеличено, вполне убедительно демонстрируют значение температуры воды как главного фактора, лимитирующего географическое распространение нектонных рыб.
Рис. 16. Схема основных пищевых связей в сообществах тропической зоны.
Этот фактор нельзя, конечно, считать единственным. Ограничивающая роль может принадлежать и другим показателям, найрййер содержанию в воде кислорода (2 мл/л — предельное значение для опускающегося вглубь альбакора), солености воды (35,5%о — минимальная, как полагают, величина для приближающейся к берегам длиннокрылой акулы) или количественному распределению кормовых организмов.
В ареалах нектонных рыб, как уже говорилось, можно различить три структурные части — область размножения, нагульную область и область выселения планктонных стадий раннего развития. Пространственное взаиморасположение репродуктивной и нагульной частей ареала у разных видов может существенно варьировать, В некоторых случаях они полностью совпадают, в других — частично перекрываются, в третьих — нацело разобщены.
Правда, при современной изученности распространения нектонных рыб невозможно выделить структурные части ареалов для большинства эпипелагических видов. Достаточно детальные представления имеются лишь о распространении относительно немногих рыб. Ареал северотихоокеанской сайры, например, состоит из репродуктивной части, занимающей южные районы области распространения (на периферии Северного субтропического круговорота), нагульной части, приходящейся на субарктические воды, и области выселения мальков в заливе Аляска. Полная или частичная пространственная разобщенность нерестовой и нагульной частей ареала имеется также у сельдевой и синей акул, скумбрещуки, субтропической летучей рыбы Hirundichthys rondeletii, длинноперого тунца, полосатого марлина, меч-рыбы и некоторых других видов. При этом основные структурные части ареалов некоторых широко распространенных нектонных рыб эпипелагиали могут располагаться в значительно различающихся условиях среды.
Значительные перемещения многих нектонных рыб в пределах своих ареалов, связанные с периодическими нерестовыми, нагульными и зимовальными миграциями, приводят к существенным изменениям границ распространения отдельных видов в разные сезоны. Особенно сильно меняется распространение в океане у тех рыб, которые обитают в водах, подверженных заметным сезонным изменениям температуры,— у сайры, лососей, некоторых тунцов и акул и т. п.
Число планктонных рыб в эпипелагиали невелико: если исключить из рассмотрения никтоэпипелагических мигрантов, к ним относятся лишь карликрвые виды семейства макрелещуковых, а также пассивные в течение всей жизни гиганты — рыбы-луны (по всей вероятности, сюда следует добавить сельдяных королей, вогмеров, парусных морских лещей рода Pteraclis). Можно думать, что географическое распространение этих рыб определяется, как и распространение других планктонных животных, циркуляцией вод океана, а основы их ареалов более или менее соответствуют крупномасштабным системам замкнутых или полузамкнутых круговоротов.
Миграции. Характерной чертой многих эпипелагических рыб служит их способность к направленным перемещениям в пространстве — вертикальным и горизонтальным миграциям. Оставляя в стороне миграции никтоэпипелагических видов (они будут рассмотрены в гл. III), следует отметить, что наибольшие по размаху вертикальные перемещения совершают тунцы рода Thunnus и особенно меч-рыба. По имеющимся данным, желтоперый тунец может опускаться до глубины 160 м, большеглазый — до 250 м, а длинноперый еще глубже — до 380 и даже 600 м. Меч-рыба также была однажды сфотографирована на глубине, превышающей 600 м.
Активные горизонтальные миграции некоторых эпипелагических рыб, обусловленные пространственным разобщением районов нереста и нагула в соответствии с различиями абиотических (прежде всего температурных) и биотических (в основном кормовых) условий, достигают очень большой протяженности и часто имеют довольно сложный характер. Они еще недостаточно изучены и лишь в немногих случаях подтверждены данными мечения, позволяющими прослеживать их пути. Иногда, впрочем, такие миграции хорошо заметны по перемещениям массовых скоплений и районов промысла. У океанических скумбриевидных рыб они достигают наибольшего размаха: у них отмечены даже сверхдальние трансокеанические миграции.
Особенно значительные перемещения совершают, по-видимому, генеративно тепловодные (т. е. имеющие тропическое происхождение) рыбы, выходящие для нагула за пределы тропических вод. Так, детальное изучение биологии длинноперого тунца (альбакора) в северной части Тихого океана показало, что для этого вида в период, предшествующий наступлению половой зрелости, характерны весьма сложные перемещения.
Опыты по мечению длинноперого тунца начались в 1952 г., а уже на„ следующий год было получено неоспоримое доказательство трансокеанских миграций: рыба, помеченная в районе Калифорнии в августе 1952 г., была обнаружена в японских водах через 323 дня после выпуска. В последующие годы в западной части Тихого океана было поймано еще пять рыб, кроме того, два тунца были возвращены из центральной части океана (район о-ва Мидуэй). Средняя скорость перемещения, по данным мечения, достигает 16—17 миль (около 30 км) в сутки. Мечение длинно-Езрого тунца производилось также в центральной части Тихого океана, восточнее Японии. Рыбы, помеченные в центре океана, ловились впоследствии и у Японии, и у Калифорнии, а тунцы, выпущенные в японских водах, добывались лишь в средней части океана.
Результаты мечения, данные по размерному и возрастному составам уловов, а также сведения о сезонных перемещениях районов промысла позволили представить в общем виде картину миграций неполовозрелого альбакора в северной половине Тихого океана. В промысловых уловах у Японии и Калифорнии длинноперый тунец, как правило, встречается, начиная с двухлетнего возраста, причем в восточной части океана добываются рыбы в возрасте от 2 до 5 лет, а в западной — от 1 до 10 лет. Исходя из возможности одной трансокеанской миграции в год, американские ученые предложили схему миграций этого вида, которая вполне согласуется с имеющимся фактическим материалом. Эта логичная картина, воссозданная в начале 60-х годов, рассматривается теперь лишь как одна из возможных моделей миграционного цикла альбакора.
Способность к дальним активным миграциям представляет собой одно из весьма существенных приспособлений рыб к обитанию в эпипелагиали. Такие миграции делают возможным более полное использование пищевых ресурсов океана, приводя в некоторых случаях даже к полному разобщению репродуктивной и вегетативной частей ареала (особенно характерны в этом отношении миграции, в ходе которых рыбы пересекают границу между тропической и бореальной областями). Именно поиски путей к расширению области нагула — основная причина активных перемещений. Нерестовые и зимовальные миграции служат, как правило, только обратному возвращению мигрантов из районов, в которых происходил их откорм.
Рассматривая миграции эпипелагических рыб, нельзя обойти молчанием вопрос о возможных механизмах их ориентации в открытых водах. Эта проблема активно разрабатывается в последние годы, но все ещё очень далека от разрешения, хотя высокая чувствительность рыб к таким факторам, как температура, соленость, химический состав воды, давно доказана в опытах и не вызывает сомнений. Среди предложенных к настоящему времени гипотез, частично проверенных экспериментально, следует отметить астронавигацию — ориентацию по Солнцу (в том числе по поляризации солнечного света) и другим небесным телам, ориентацию по магнитным полям (у некоторых рыб, в том числе тунцов и марлинов, в черепных костях обнаружен биогенный магнетит), по электрическим полям (восприятие сверхслабых токов может обеспечить ориентацию относительно течений) и даже по гравитационным полям. Все эти гипотезы не дают полного объяснения наблюдаемым в природе процессам, однако твердо установленный факт направленных перемещений некоторых видов рыб требует продолжения научного поиска. Решение вопроса, возможно, будет найдено в результате комплексного подхода.
Итак, подведем некоторые итоги. Эпипелагический ихтиоцен не очень разнообразен в отношении систематического состава, но включает довольно разнородные элементы (табл. 2). Помимо постоянных обитателей поверхностных слоев открытого океана — голоэпипелагпческпх рыб — в его состав входят мероэпипелагические рыбы, проводящие здесь определенную часть жизни, и ксеноэпипелагическне рыбы, нормально живущие в других биотопах. Всего в океанской эпипелагиали представлено не более 300 видов (исключая ранние стадии развития прибрежных и глубинных рыб), принадлежащих, насколько известно, к 53 семействам. Голоэпипелагические виды, число которых не превышает 140, входят в состав 35 семейств, но лишь немногие из этих семейств (всего 16) полностью ограничены в своем распространении эпипелагиально.( Все эти цифры меньше приведенных в моей сводке «Ихтиофауна океанской эпипелагиали» (1968): за прошедшие 20 лет выяснилось, что в состав этой группы включались и но характерные для нее виды. Впрочем, и сейчас «эпипелагиаль» нередко понимается слишком широко.)
Обособленность эпипелагического биотопа лучше всего выражена в тропической зоне — там, где постоянно существующий слой теплых вод («термосфера» А. Брууна), обычная толщина которого составляет 100—200 м, отделен от холодного глубинного слоя главным термоклином, также имеющим постоянный характер. В умеренных и высоких широтах термическая обособленность верхних слоев существует только летом, когда формируется слой скачка, разрушающийся в холодный сезон вертикальной конвекцией в результате охлаждения поверхности воды. Соответственно этому эпипелагический ихтиоцен вполне четко выражен только в тропической зоне, в которой и обитают почти все упоминавшиеся виды. Достаточно сказать, что для перечисления голоэпипелагических рыб, постоянно живущих к северу и югу от 40-х широт, вполне хватило бы пальцев на руках — список «холодолюбивых» видов включает гигантскую акулу, по 2 вида сельдевых акул и морских лещей рода Brama, 2—3 вида строматеевидных рыб, низкотелого опаха, гастерохизму, и даже добавление к этому перечню тех видов, которые заходят в умеренные воды для летнего нагула (сайра, макрелещука, альбакор, меч-рыба) или заносятся течениями (луна-рыба, пятнистый опах) не меняет существа дела. Воды северной части Тихого океана, Северной Атлантики и Субантарктики характеризуются явным преобладанием мероэпипелагических и ксеноэпипелагических рыб, иногда достигающих значительной численности. В Арктике и Антарктике специфическое рыбное население пелагиали крайне бедно (голоэпипелагических рыб нет вообще), но включает очень своеобразный криопелагический ихтиоцен, впервые выделенный А. П. Андрияшевым. Криопелагические виды — антарктические широколобики (род Pagothenia), арктические тресковые рыбы — сайка (Boreogadus saida) и ледовая треска (Arctogadus glacialis) — в массовом количестве живут в непосредственном контакте с припайным и дрейфующим льдом. Эти рыбы потребляют в пищу подледный планктон и сами служат кормом многим хищникам, в том числе ныряющим птицам и морским млекопитающим.
Таблица 2
| таксоцен | Группировка видов | всего | ||||||||
| голоэпипелагическая * | мероэпипелагическая ** | ксеноэпипелагическая *** | ||||||||
| верхняя | нижняя | весь состав | неритическая | глубинная | весь состав | неритическая | глубинная | весь состав | ||
| щележаберные рыбы | 4 | 2 | 11 | - | 1 | 1 | 10 | 18 | 28 | 40 |
| костистые рыбы | 45 | 53 | 127 | 13 | 45 | 58 | 49 | 4 | 53 | 238 |
| весь ихтиоцен | 49 | 65 | 138 | 13 | 46 | 59 | 67 | 14 | 81 | 278 |
(* В графу Весь состав включены верхне и нижнеэпипелагические виды и виды, обитающие во всём диапазоне глубин.
** Без видов, представленных молодью
*** Редкие виды и виды, представленные только ранними стадиями развития не учтены.)
Для эпипелагиали особенно характерны рыбы крупного размера, относящиеся главным образом к нектону, способные к быстрому продолжительному плаванию и ведущие активный образ жизни,— именно здесь обитают рыбы-рекордисты по длине и массе тела (китовая акула — 15,2 м и 14 т), скорости плавания (меч-рыба — 130 км/ч) и его дальности (синий тунец — 5800 миль, т. е. 10 тыс. км). Планктонных рыб в этом биотопе немного, но и среди них имеются настоящие гиганты (луна-рыба «диаметром» 3 м и массой 1,5 т). Почти все обитатели эпипелагиали имеют широкие пищевые спектры, быстро растут, рано достигают половой зрелости, имеют сравнительно непродолжительную жизнь и большую индивидуальную плодовитость при отсутствии заботы о потомстве. По внешнему облику, образу жизни и экологическим характеристикам они резко отличаются от рыб глубоководной пелагиали.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://animalkingdom.su/ 'Мир животных'